Topic: - Plant Responses and Adaptive Mechanisms Under Heat Stress
Abstract
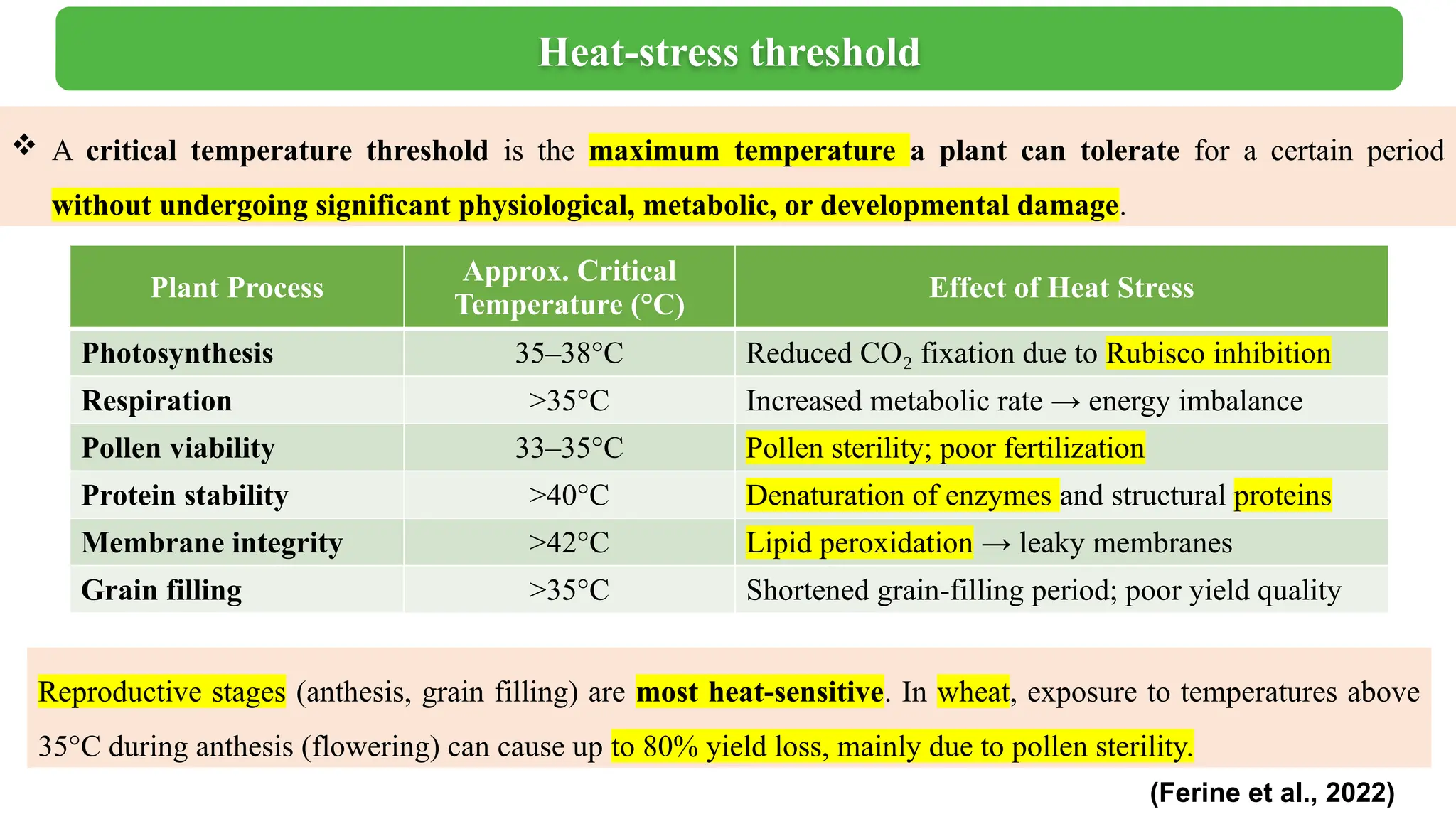
Heat stress is a major abiotic factor affecting crop productivity worldwide, particularly under the increasing pressure of climate change. High temperatures during day & night crucial growth stages (especially flowering and grain filling) can disrupt cellular homeostasis, reduce pollen viability, and lead to spikelet sterility and yield loss. Plants have evolved various adaptive mechanisms to mitigate these effects, including physiological, biochemical, and molecular responses. One of the most critical traits under heat stress is pollen viability, which directly determines the reproductive success of plants. Transpiration cooling helps lower leaf temperature and protect floral organs, enhancing heat tolerance in plants. From a biochemical point of view, the accumulation of osmotic regulating substances such as proline and the activity of antioxidant enzymes (SOD, CAT, POD) help to eliminate harmful reactive oxygen species (ROS), thereby reducing oxidative damage. Thermotolerant genotypes maintain higher photosynthetic rates, chlorophyll stability, and stomatal conductance, thereby supporting biomass accumulation and yield. At the molecular level, heat tolerance is supported by the expression of heat shock proteins (HSPs) and their regulatory genes, heat shock transcription factors (HSFs), which protect proteins from denaturation and assist in protein refolding during stress events. Plant heat tolerance is a complex trait determined by the interaction of morphological, physiological, biochemical and molecular factors. Integrating these insights into breeding programs can facilitate the development of climate-resilient plant varieties that can withstand rising global temperatures.
Keywords: Antioxidant enzymes, heat stress, membrane stability, pollen viability, transpiration cooling





![How Heat Stress Causes in Plants?
https://www.researchgate.net/figure/Possible-effects-of-heat-stress-on-different-parts-of-plants_n 2025] fig1_333853824](https://image.slidesharecdn.com/plantresponsesandadaptivemechanismsunderheatstress-251205100140-1291e1a0/75/Plant-Responses-and-Adaptive-Mechanisms-Under-Heat-Stress-pptx-6-2048.jpg)










































![Devi, N., Kumari, A., Yadav, S., & Kumar, M. (2022). Heat stress effect on growth and reproductive traits in chilli (Capsicum
spp.). Journal of Pharmacognosy and Phytochemistry, 11(5), 2590–2594.
Das, R., & Prasad, P. V. V. (2022). Reproductive stage heat stress responses in papaya (Carica papaya L.): Physiological insights
and implications. Scientia Horticulturae, 304, 111280. https://doi.org/10.1016/j.scienta.2022.111280
Aryan, S., Gulab, G., Habibi, N., Amin, M. W., Habibi, S., & Irie, K. (2025). Physiological responses of rice varieties to heat
stress: exploring photosynthetic efficiency, oxidative stress, and yield resilience. Discover Plants.
Devasirvatham, V., Gaur, P. M., Mallikarjuna, N., Raju, T. N., Trethowan, R. M., & Tan, D. K. Y. (2013). Reproductive biology of
chickpea response to heat stress in the field is associated with the performance in controlled environments. Field Crops
Research, 142, 9–19. https://doi.org/10.1016/j.fcr.2012.11.011
Malasala, M., Dhekale, B., & Mohanty, U. C. (2018). Impact of climate variability on various Rabi crops over Northwest India.
Theoretical and Applied Climatology, 131, DOI: 10.1007/s00704-016-1991-7.
Approaches in Enhancing Thermotolerance in Plants: An Updated Review - Scientific Figure on ResearchGate. Available from:
https://www.researchgate.net/figure/Possible-effects-of-heat-stress-on-different-parts-of-plants_fig1_333853824
[accessed 26 Jun 2025]](https://image.slidesharecdn.com/plantresponsesandadaptivemechanismsunderheatstress-251205100140-1291e1a0/75/Plant-Responses-and-Adaptive-Mechanisms-Under-Heat-Stress-pptx-50-2048.jpg)
































































