Downloaded 30 times



![Severe fungal infection in a fruit fly (color) with a disabling mutation
in the signal-transduction pathway required for the synthesis of the
antifungal peptide drosomycin.
[Electron micrograph adapted from B. Lemaitre et al., 1996, Cell 86: 973;
courtesy of J.A. Hoffman, University of Strasbourg.]
Toll-LIKE RECEPTORS (TLRs)](https://image.slidesharecdn.com/nucleicacidrecognitionbytheinnateimmunesystem-160226055050/85/Nucleic-acid-recognition-by-the-innate-immune-system-3-320.jpg)
![ TLR 3
First TLR implicated in viral nucleic acid sensing
Binds with dsRNA with very less sequence specificty
Minimum 40bp length required for responsiveness and
affinity increased in proportion to dsRNA length
(Botos et al., 2009)
Also recognize synthetic RNA analogs:
Poly inosine – poly cytidylic acid [poly(I:C)]
(Alexopoulou et al., 2001)
Recognizes the genomic DNA of Reo virus, dsRNA
intermediates of West Nile virus, encephalomyocarditis virus
and certain siRNAs
LIGAND SPECIFICITY AND MICROBIAL
RECOGNITION](https://image.slidesharecdn.com/nucleicacidrecognitionbytheinnateimmunesystem-160226055050/85/Nucleic-acid-recognition-by-the-innate-immune-system-7-320.jpg)



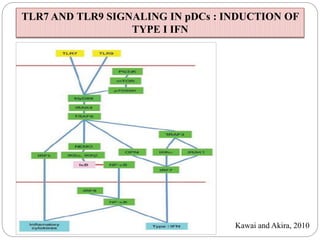



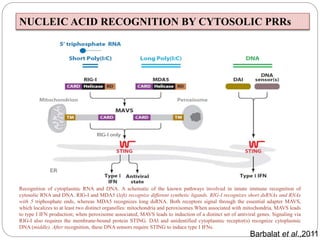
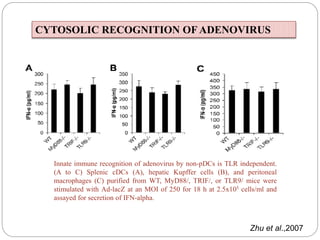
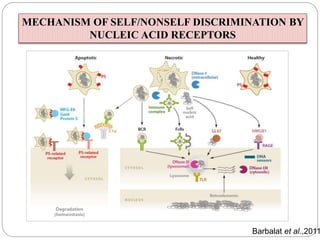





The document discusses the recognition of nucleic acids by the innate immune system, focusing on various pattern recognition receptors (PRRs) such as Toll-like receptors (TLRs) and RIG-I-like receptors (RLRs). It highlights the mechanisms by which these receptors identify viral and bacterial nucleic acids, initiate immune responses, and their roles in maintaining immunological balance. Additionally, it covers the therapeutic implications of these interactions for treating infections and autoimmune diseases.










































































