Downloaded 1,428 times



![Toll: Origin of the word (1985)
The gene in question, when mutated, makes the Drosophila (fruit fly) embryo look
unusual.
The researchers were so surprised that they spontaneously shouted out in German
"Das ist ja toll!" which translates as "That's great!".
[The Nobel Prize in Physiology or Medicine 1995:Edward B. Lewis, Christiane
Nüsslein-Volhard, Eric F. Wieschaus]](https://image.slidesharecdn.com/tolllikereceptors-modified-130212110903-phpapp01/75/Toll-like-receptors-4-2048.jpg)



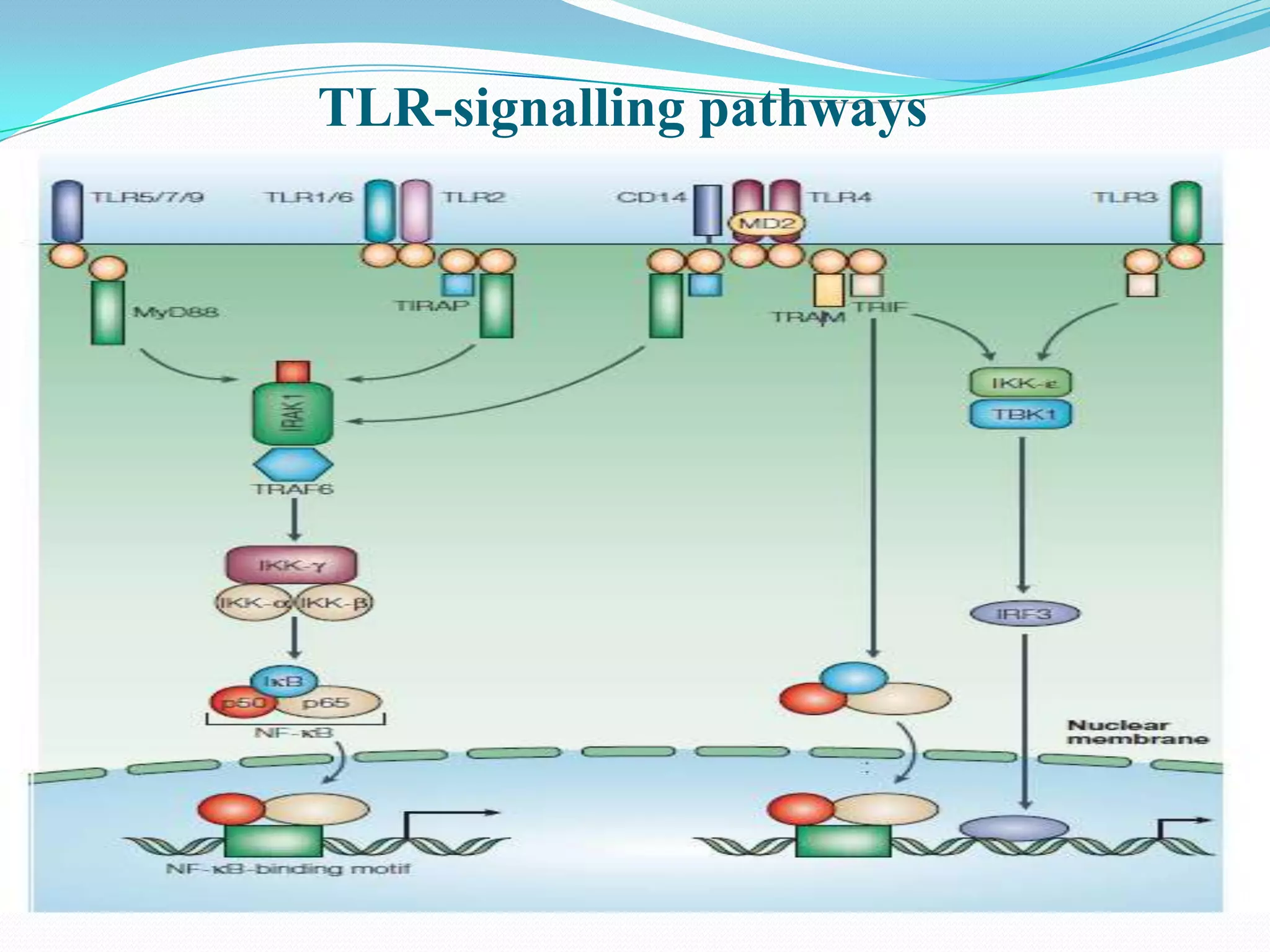
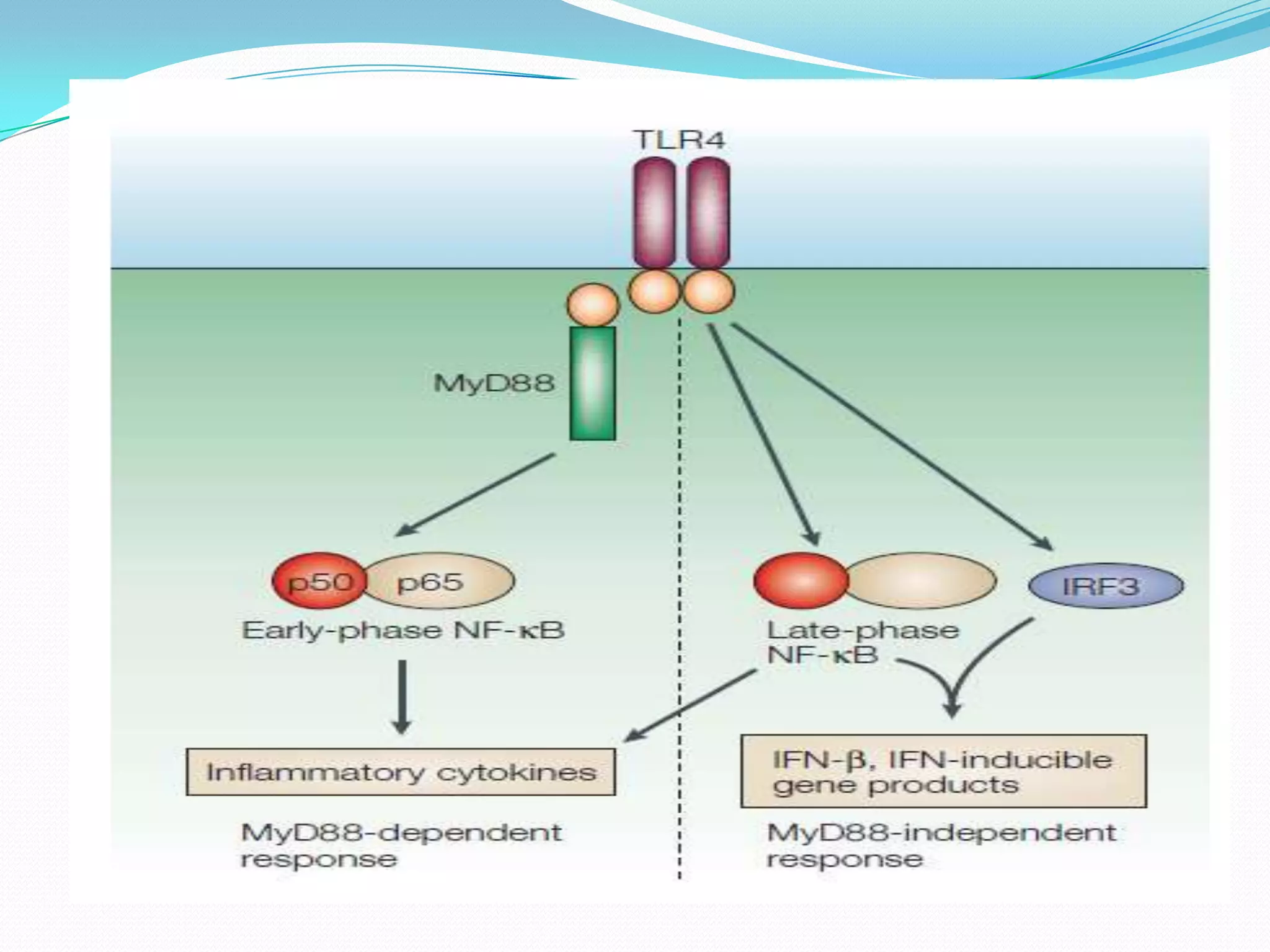
![TLR Signaling Pathway
1]MyD88 (myeloid differentiation primary-response protein 88)
Forms homodimers through DD–DD and TIR-domain–TIR-domain interactions and exists
as a dimer when recruited to the receptor complex.
Functions as adaptor linking TLRs/IL-1Rs with downstream signalling molecules that have
DDs.
2] IRAK family (IL-1-receptor-associated kinases)
Four IRAKs — IRAK1,IRAK2,IRAK4 and IRAK-M identified
The kinase activity of IRAK1 increases strongly following TLR/IL-1R stimulation, and its
kinase domain is essential for signalling through nuclear factor-κB (NF-κB)
3] TRAF6 (tumour-necrosis-factorreceptor- associated factor 6)
Signalling mediator for both the TNF-receptor superfamily and the TLR/IL-1R
superfamily, interacting directly with members of the TNF-receptor superfamily
4] NF-κB (nuclear factor-κB)
Promote expression of genes and synthesis of cytokines](https://image.slidesharecdn.com/tolllikereceptors-modified-130212110903-phpapp01/75/Toll-like-receptors-18-2048.jpg)


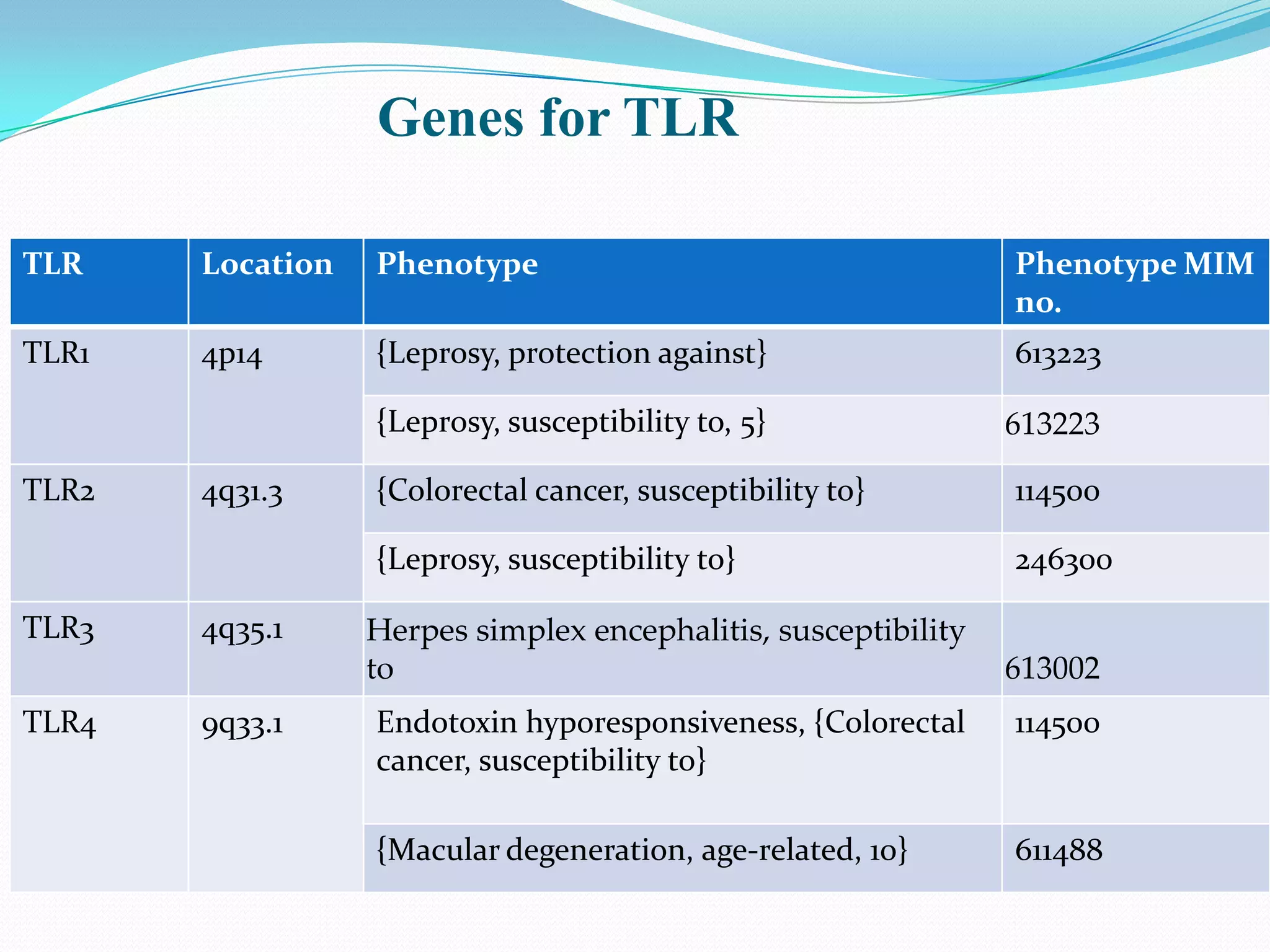
![TLR 1
1]PROTECTION AGAINST LEPROSY
SNP in TLR1, 1805T-G, that results in an ile602-to-ser (I602S) substitution
at the junction of the transmembrane and intracellular domains of TLR1.[
Johnson et al. 2007, Misch et al.2008]
2] SUSCEPTIBILITY TO LEPROSY
Association of an asn248-to-ser (N248S) SNP in the TLR1 gene and leprosy
(LPRS5; 613223) in a Bangladeshi population consisting of 842 patients
and 543 controls.[Schuring et al. (2009)]
Homozygosity for S248 significantly associated with leprosy (OR = 1.34)
& heterozygosity was found to be protective against leprosy (OR = 0.78)
Homozygous N248 genotype was equally distributed among patients and
controls
Patients with erythema nodosum leprosum reactions were more likely to
have the N248 allele (68%) than were patients who had no reactions (46%)](https://image.slidesharecdn.com/tolllikereceptors-modified-130212110903-phpapp01/75/Toll-like-receptors-24-2048.jpg)
![TLR2
1]Susceptibility to Leprosy
Arg677-to-trp polymorphism (R677W; 603028.0001) in the intracellular domain
of TLR2 in 10 (22%) of 45 Korean lepromatous leprosy patients [Kang and Chae
2001]
R677W leads to poor cellular immune response associated with lepromatous
leprosy.[Bochud et al. 2003]
R677W was undetectable in the Japanese patients [Mikita et al. 2009], similar to
the findings in Indian patients reported by [Malhotra et al. 2005]
Bochud et al. (2008 )analyzed 3 TLR2 polymorphisms in 441 patients and 187
controls in 3 Ethiopian groups.
597C-T SNP was associated with reduced susceptibility to reversal reaction
Homozygous for a 280-bp microsatellite marker had an increased risk of reversal
reaction](https://image.slidesharecdn.com/tolllikereceptors-modified-130212110903-phpapp01/75/Toll-like-receptors-25-2048.jpg)
![TLR2
1] Lyme Disease
Monocytes and lymphocytes from healthy subjects produced more TNF
and IFNG, respectively, in response to high concentrations of Borrelia
lysate than did healthy subjects heterozygous for an arg753-to-gln (R753Q)
SNP. [Schroder et al. 2005]
R753Q SNP may protect from development of late-stage Lyme disease due
to reduced signaling via TLR2/TLR1.
2] Tuberculosis
Increased risk of TB in carriers of a nonsynonymous 2258G-A SNP in the
TLR2 gene, which causes the R753Q substitution [Ogus et al. (2004)]
3] Colorectal Cancer
Boraska Jelavic et al. (2006) GT microsatellite repeat polymorphism in
intron 2 of the TLR2 gene in sporadic colorectal cancer patients
Frequency of TLR2 alleles with 20 and 21 GT repeats was decreased &
that of 31 GT repeats was increased in patients versus controls.](https://image.slidesharecdn.com/tolllikereceptors-modified-130212110903-phpapp01/75/Toll-like-receptors-26-2048.jpg)
![TLR3
1] Herpes Simplex Encephalitis
Zhang et al. (2007) detected a heterozygous P554S mutation in the
TLR3 gene. The mutation occurred on different TLR3 haplotypes in
the children.
TLR3 is vital for natural immunity to HSV-1 in the CNS and
neurotropic viruses have contributed to the evolutionary maintenance
of TLR3.
2] Age-related macular degeneration
T allele of rs3775291 in the TLR3 gene, which results in a L412F
substitution, is protective against the development of geographic
atrophy or advanced dry age-related macular degeneration](https://image.slidesharecdn.com/tolllikereceptors-modified-130212110903-phpapp01/75/Toll-like-receptors-27-2048.jpg)
![TLR4 (The human homolog of Drosophila Toll)
1] ENDOTOXIN HYPORESPONSIVENESS
Arbour et al. (2000) showed that 2 common cosegregating missense mutations
(asp299 to gly and thr399 to ile) that affect the extracellular domain of the TLR4
receptor are associated with blunted response to inhaled lipopolysaccharide in
humans.
Susceptibility to Gm negative sepsis.
2] D299G allele - Lower Levels Of Certain Proinflammatory Cytokines, More
Susceptibility To Severe Bacterial Infections, Lower Risk Of Carotid
Atherosclerosis, And A Smaller Intima-media Thickness In The Common Carotid
Artery. [Kiechl et al. (2002)]
3] gly299 allele of the TLR4 gene was more frequent in colorectal cancer patients
than controls
4] D299G and T399I variants of TLR4 as contributors to susceptibility to age-related
macular degeneration
5] D299G polymorphism– metastasis after breast cancer surgery
6] D299G polymorphism in TLR4 may influence the immunologic component of
anthracycline-based chemotherapy in human cancer.](https://image.slidesharecdn.com/tolllikereceptors-modified-130212110903-phpapp01/75/Toll-like-receptors-28-2048.jpg)
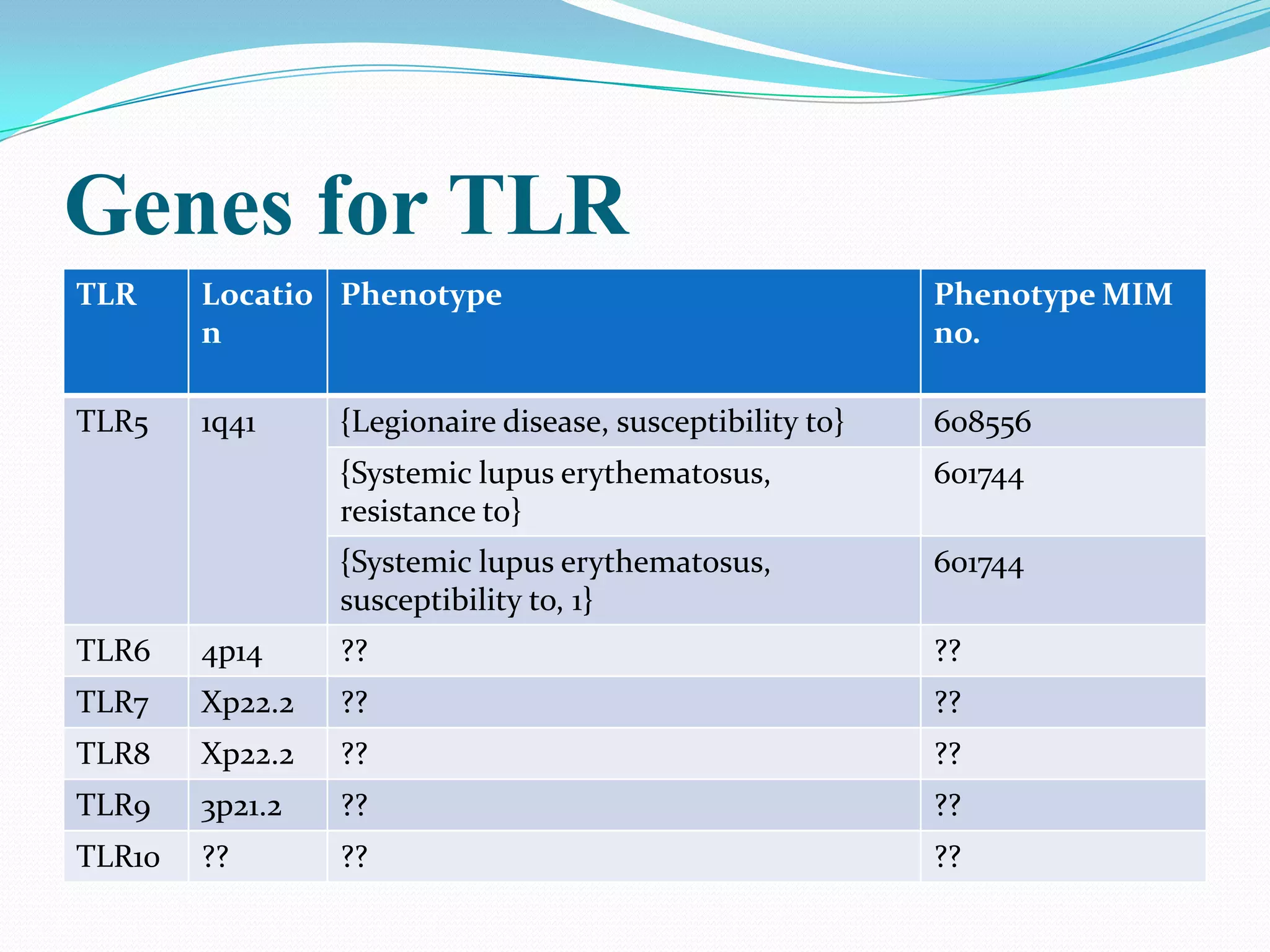
![TLR5
1] Susceptibility to Legionnaire disease
C-to-T transition at nucleotide 1174, changed arg392 to a stop codon
(R392X) [
Hawn et al. 2003]
2] Resistance to systemic lupus erythematosus
allele 1174C of TLR5, but not allele 1174T, which encodes the premature
stop codon, was preferentially transmitted to SLE-affected
offspring. [Hawn et al. 2005]](https://image.slidesharecdn.com/tolllikereceptors-modified-130212110903-phpapp01/75/Toll-like-receptors-29-2048.jpg)





![Neurological diseases with possible link to TLR
pathway
1]Leprosy 9]Bacterial meningitis
2]Herpes Simplex Encephalitis 10] Alzheimer’s Disease
3]Entero and flaviviral 11] Prion Diseases
encephalitis 12] Amyotrophic Lateral
4]Malaria Sclerosis.
5]Toxoplasmosis 13] Parkinson’s Disease
6]Trypanosomiasis 14]Perinatal brain injury
7]Lyme disease 15] Multiple sclerosis
8]Neurocysticercosis and ?????????????????](https://image.slidesharecdn.com/tolllikereceptors-modified-130212110903-phpapp01/75/Toll-like-receptors-35-2048.jpg)








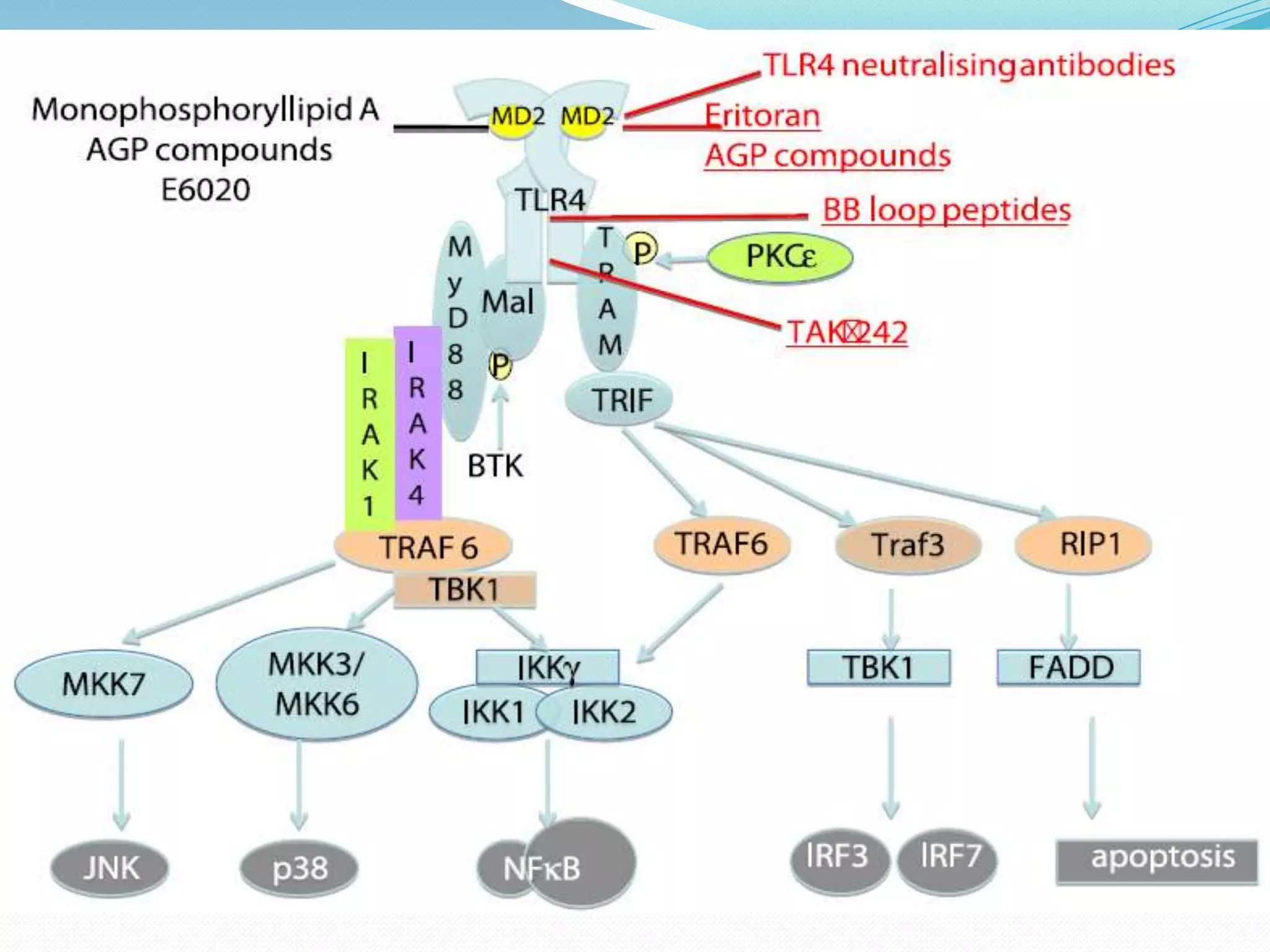




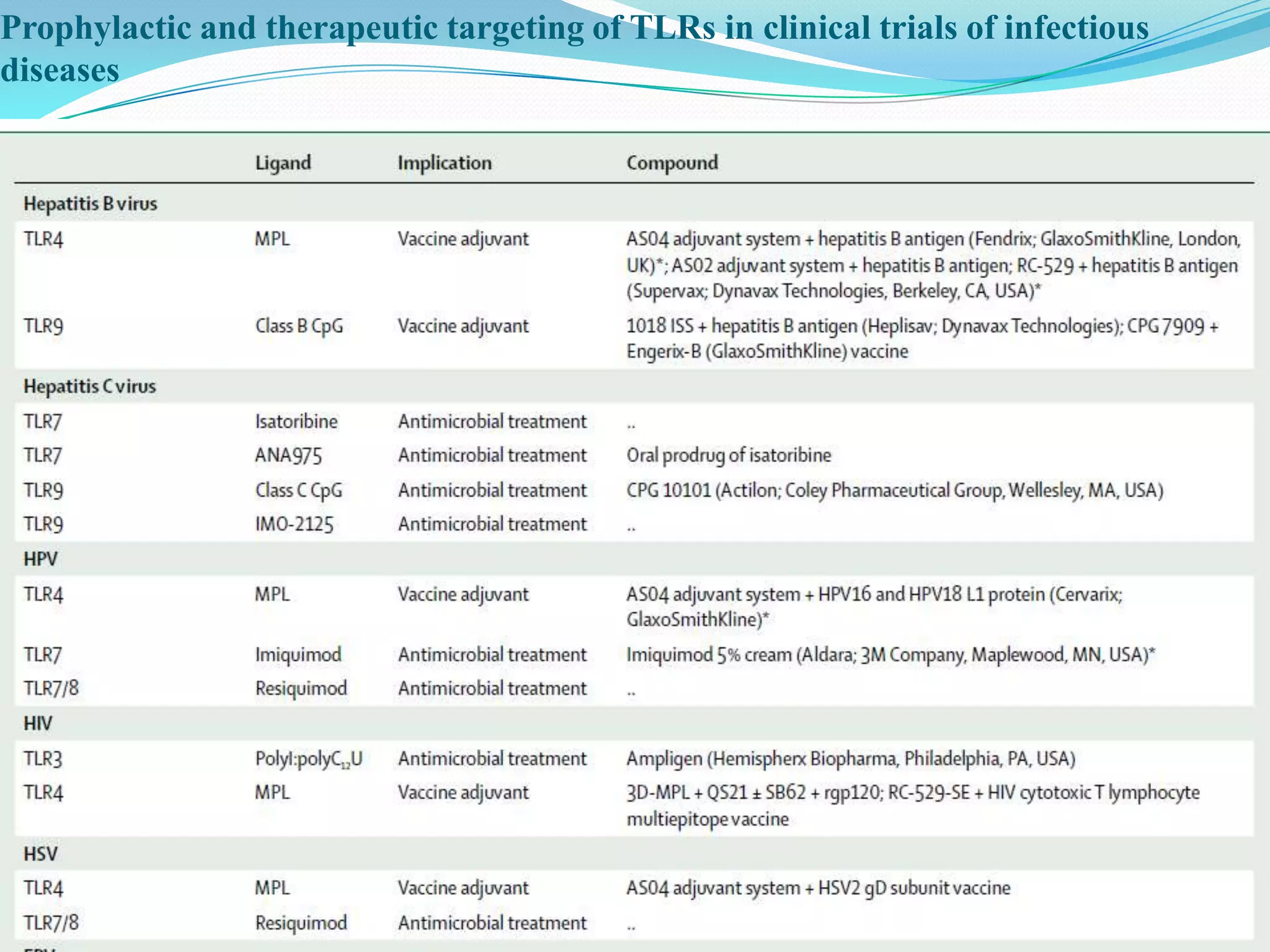
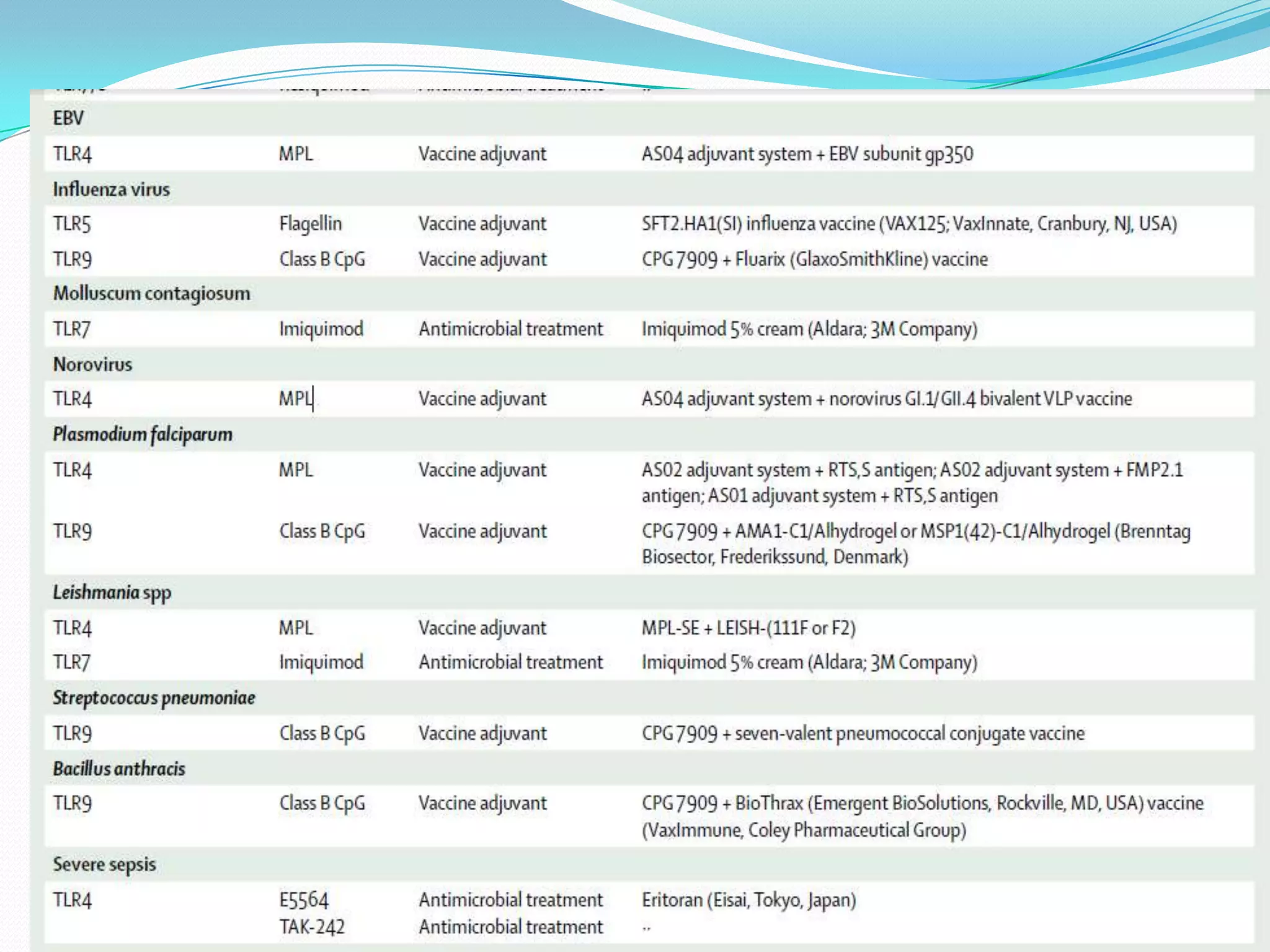




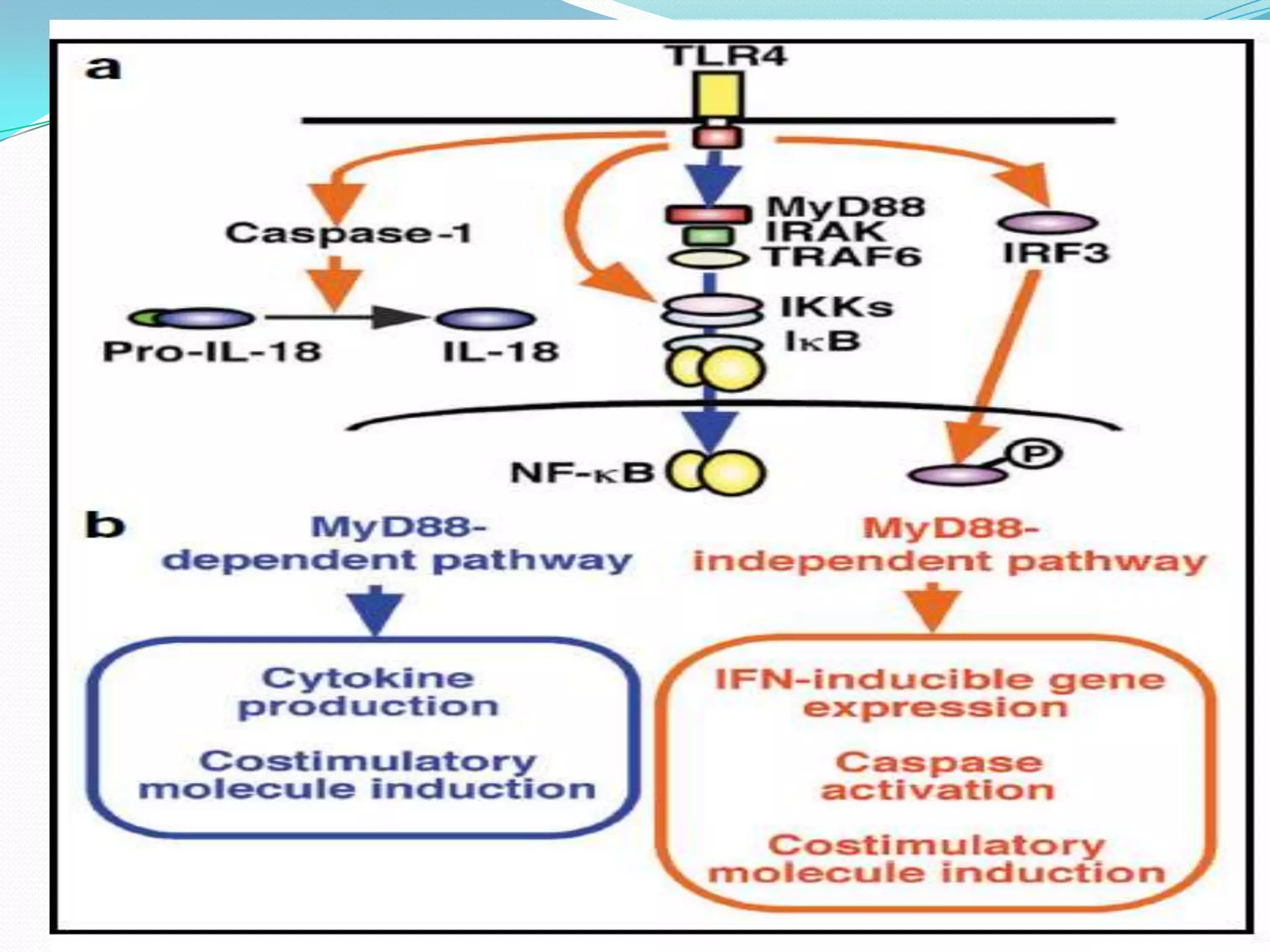
This document summarizes key information about Toll-like receptors (TLRs): - TLRs are pattern recognition receptors that recognize pathogens and activate immune responses. They play a role in both innate and adaptive immunity. - TLRs recognize specific microbial ligands and signal through either MyD88-dependent or MyD88-independent pathways to induce inflammatory responses. - Genetic variations in TLRs have been linked to susceptibility or resistance to various diseases like leprosy, tuberculosis, and cancer. Targeting TLR pathways may offer therapeutic approaches for neurological diseases like Alzheimer's disease.













































































