Inability of flowering plants to produce functional pollen.
Male sterility is agronomically important for the hybrid seed production.
Onion crop provides one of the rare examples of very early recognition of male sterility cultivar Italian Red (Jones and Emsweller 1936)
Its inheritance and use in hybrid seed production (Jones
and Clarke 1943).
Since then male sterility is reported in a fairly large number of crops including vegetables.
Molecular control of male fertility for crop hybrid breedingSuresh Antre
Harnessing hybrid vigor or heterosis is a promising approach to tackle the current challenge of sustaining enhanced yield gains of field crops. More than half of the production of major crops such as maize, rice, sorghum, rapeseed, and sunflower comes from hybrid varieties.
1. STABILITY OF MALE STERILE LINES - ENVIRONMENTAL INFLUENCE ON STERILITY - EGMS - TYPES AND INFLUENCE ON THEIR EXPRESSION, GENETIC STUDIES.
2. PHOTO SENSITIVE GENETIC MALE STERILITY AND ITS USES IN HETEROSIS BREEDING
3. TEMPERATURE SENSITIVE GENETIC MALE STERILITY AND ITS USES IN HETEROSIS BREEDING
FERTILITY RESTORATION IN MALE STERILE LINES AND RESTORER DIVERSIFICATION PROG...Rachana Bagudam
1. FERTILITY RESTORATION IN MALE STERILE LINES AND RESTORER DIVERSIFICATION PROGRAMMES.
2. CONVERSION OF AGRONOMICALLY IDEAL GENOTYPES INTO MALE STERILES.
3. GENERATING NEW CYTONUCLEAR INTERACTION SYSTEM FOR DIVERSIFICATION OF MALE STERILES.
Male sterility is the inability of plants to produce or to release functional pollens (microspores) or gametes to produce offspring.
OR
Absence or malformation of male organs (stamens) in bisexual plants or no male flowers in dioecious plants
Anther abortion in intragenic and between hybrids first observed by Koelreuter in 1763.
Inability of flowering plants to produce functional pollen.
Male sterility is agronomically important for the hybrid seed production.
Onion crop provides one of the rare examples of very early recognition of male sterility cultivar Italian Red (Jones and Emsweller 1936)
Its inheritance and use in hybrid seed production (Jones
and Clarke 1943).
Since then male sterility is reported in a fairly large number of crops including vegetables.
Molecular control of male fertility for crop hybrid breedingSuresh Antre
Harnessing hybrid vigor or heterosis is a promising approach to tackle the current challenge of sustaining enhanced yield gains of field crops. More than half of the production of major crops such as maize, rice, sorghum, rapeseed, and sunflower comes from hybrid varieties.
1. STABILITY OF MALE STERILE LINES - ENVIRONMENTAL INFLUENCE ON STERILITY - EGMS - TYPES AND INFLUENCE ON THEIR EXPRESSION, GENETIC STUDIES.
2. PHOTO SENSITIVE GENETIC MALE STERILITY AND ITS USES IN HETEROSIS BREEDING
3. TEMPERATURE SENSITIVE GENETIC MALE STERILITY AND ITS USES IN HETEROSIS BREEDING
FERTILITY RESTORATION IN MALE STERILE LINES AND RESTORER DIVERSIFICATION PROG...Rachana Bagudam
1. FERTILITY RESTORATION IN MALE STERILE LINES AND RESTORER DIVERSIFICATION PROGRAMMES.
2. CONVERSION OF AGRONOMICALLY IDEAL GENOTYPES INTO MALE STERILES.
3. GENERATING NEW CYTONUCLEAR INTERACTION SYSTEM FOR DIVERSIFICATION OF MALE STERILES.
Male sterility is the inability of plants to produce or to release functional pollens (microspores) or gametes to produce offspring.
OR
Absence or malformation of male organs (stamens) in bisexual plants or no male flowers in dioecious plants
Anther abortion in intragenic and between hybrids first observed by Koelreuter in 1763.
Speed Breeding and its implications in crop improvementANILKUMARDASH2
Introduction
History of speed breeding
Methods of speed breeding
Advantages over conventional breeding
Integration with various technologies
Case studies
Opportunities and challenges
Conclusions
Multiple inbred founder lines are inter-mated for several generations prior to creating inbred lines, resulting in a diverse population whose genomes are fine scale mosaics of contributions from all founders.
The term balanced tertiary trisomic has three words of which (1) “trisomic” indicates the presence of extra chromosome, (2) “tertiary” indicates that the extra chromosome is a trans-located chromosome, and (3) “balanced” refers to the breeding behaviour of the trisomic.
Ramage defined the BTT as a tertiary trisomic constructed in such a way that the dominant allele of a marker gene, closely linked with the translocation breakpoint of the extra chromosome is carried on the extra chromosome, and the recessive allele is carried on the two normal chromosomes that constitute the diploid complement. The dominant marker gene may be located on the centromere segment or the trans-located segment of the extra chromosome.
Speed Breeding and its implications in crop improvementANILKUMARDASH2
Introduction
History of speed breeding
Methods of speed breeding
Advantages over conventional breeding
Integration with various technologies
Case studies
Opportunities and challenges
Conclusions
Multiple inbred founder lines are inter-mated for several generations prior to creating inbred lines, resulting in a diverse population whose genomes are fine scale mosaics of contributions from all founders.
The term balanced tertiary trisomic has three words of which (1) “trisomic” indicates the presence of extra chromosome, (2) “tertiary” indicates that the extra chromosome is a trans-located chromosome, and (3) “balanced” refers to the breeding behaviour of the trisomic.
Ramage defined the BTT as a tertiary trisomic constructed in such a way that the dominant allele of a marker gene, closely linked with the translocation breakpoint of the extra chromosome is carried on the extra chromosome, and the recessive allele is carried on the two normal chromosomes that constitute the diploid complement. The dominant marker gene may be located on the centromere segment or the trans-located segment of the extra chromosome.
Earliest Galaxies in the JADES Origins Field: Luminosity Function and Cosmic ...Sérgio Sacani
We characterize the earliest galaxy population in the JADES Origins Field (JOF), the deepest
imaging field observed with JWST. We make use of the ancillary Hubble optical images (5 filters
spanning 0.4−0.9µm) and novel JWST images with 14 filters spanning 0.8−5µm, including 7 mediumband filters, and reaching total exposure times of up to 46 hours per filter. We combine all our data
at > 2.3µm to construct an ultradeep image, reaching as deep as ≈ 31.4 AB mag in the stack and
30.3-31.0 AB mag (5σ, r = 0.1” circular aperture) in individual filters. We measure photometric
redshifts and use robust selection criteria to identify a sample of eight galaxy candidates at redshifts
z = 11.5 − 15. These objects show compact half-light radii of R1/2 ∼ 50 − 200pc, stellar masses of
M⋆ ∼ 107−108M⊙, and star-formation rates of SFR ∼ 0.1−1 M⊙ yr−1
. Our search finds no candidates
at 15 < z < 20, placing upper limits at these redshifts. We develop a forward modeling approach to
infer the properties of the evolving luminosity function without binning in redshift or luminosity that
marginalizes over the photometric redshift uncertainty of our candidate galaxies and incorporates the
impact of non-detections. We find a z = 12 luminosity function in good agreement with prior results,
and that the luminosity function normalization and UV luminosity density decline by a factor of ∼ 2.5
from z = 12 to z = 14. We discuss the possible implications of our results in the context of theoretical
models for evolution of the dark matter halo mass function.
A brief information about the SCOP protein database used in bioinformatics.
The Structural Classification of Proteins (SCOP) database is a comprehensive and authoritative resource for the structural and evolutionary relationships of proteins. It provides a detailed and curated classification of protein structures, grouping them into families, superfamilies, and folds based on their structural and sequence similarities.
What is greenhouse gasses and how many gasses are there to affect the Earth.moosaasad1975
What are greenhouse gasses how they affect the earth and its environment what is the future of the environment and earth how the weather and the climate effects.
Multi-source connectivity as the driver of solar wind variability in the heli...Sérgio Sacani
The ambient solar wind that flls the heliosphere originates from multiple
sources in the solar corona and is highly structured. It is often described
as high-speed, relatively homogeneous, plasma streams from coronal
holes and slow-speed, highly variable, streams whose source regions are
under debate. A key goal of ESA/NASA’s Solar Orbiter mission is to identify
solar wind sources and understand what drives the complexity seen in the
heliosphere. By combining magnetic feld modelling and spectroscopic
techniques with high-resolution observations and measurements, we show
that the solar wind variability detected in situ by Solar Orbiter in March
2022 is driven by spatio-temporal changes in the magnetic connectivity to
multiple sources in the solar atmosphere. The magnetic feld footpoints
connected to the spacecraft moved from the boundaries of a coronal hole
to one active region (12961) and then across to another region (12957). This
is refected in the in situ measurements, which show the transition from fast
to highly Alfvénic then to slow solar wind that is disrupted by the arrival of
a coronal mass ejection. Our results describe solar wind variability at 0.5 au
but are applicable to near-Earth observatories.
Professional air quality monitoring systems provide immediate, on-site data for analysis, compliance, and decision-making.
Monitor common gases, weather parameters, particulates.
This pdf is about the Schizophrenia.
For more details visit on YouTube; @SELF-EXPLANATORY;
https://www.youtube.com/channel/UCAiarMZDNhe1A3Rnpr_WkzA/videos
Thanks...!
Deep Behavioral Phenotyping in Systems Neuroscience for Functional Atlasing a...Ana Luísa Pinho
Functional Magnetic Resonance Imaging (fMRI) provides means to characterize brain activations in response to behavior. However, cognitive neuroscience has been limited to group-level effects referring to the performance of specific tasks. To obtain the functional profile of elementary cognitive mechanisms, the combination of brain responses to many tasks is required. Yet, to date, both structural atlases and parcellation-based activations do not fully account for cognitive function and still present several limitations. Further, they do not adapt overall to individual characteristics. In this talk, I will give an account of deep-behavioral phenotyping strategies, namely data-driven methods in large task-fMRI datasets, to optimize functional brain-data collection and improve inference of effects-of-interest related to mental processes. Key to this approach is the employment of fast multi-functional paradigms rich on features that can be well parametrized and, consequently, facilitate the creation of psycho-physiological constructs to be modelled with imaging data. Particular emphasis will be given to music stimuli when studying high-order cognitive mechanisms, due to their ecological nature and quality to enable complex behavior compounded by discrete entities. I will also discuss how deep-behavioral phenotyping and individualized models applied to neuroimaging data can better account for the subject-specific organization of domain-general cognitive systems in the human brain. Finally, the accumulation of functional brain signatures brings the possibility to clarify relationships among tasks and create a univocal link between brain systems and mental functions through: (1) the development of ontologies proposing an organization of cognitive processes; and (2) brain-network taxonomies describing functional specialization. To this end, tools to improve commensurability in cognitive science are necessary, such as public repositories, ontology-based platforms and automated meta-analysis tools. I will thus discuss some brain-atlasing resources currently under development, and their applicability in cognitive as well as clinical neuroscience.
PRESENTATION ABOUT PRINCIPLE OF COSMATIC EVALUATION
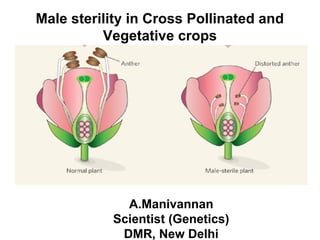
Male Sterility IN Cross Pollinated and Vegetable Crops
1. Male sterility in Cross Pollinated and
Vegetative crops
A.Manivannan
Scientist (Genetics)
DMR, New Delhi
2.
3. Male sterility in Sunflower (Helianthus spp)
Genetic Male sterility (GMS)
Complete male sterility
ms1-ms5 = male sterility in sunflower recessive gene
Two types of g-mst
Type 1-gmst-Bloomington type
Type 2-gmst-Modern type
Cultivated Sunflower variety Karlik-68(Dwarf 68)- two recessive genes
msi1,msi2(Stable and complete male sterile)
Partial male sterility –p mst
4. Source of cms through Interspecific
Hybridization
x H.annuusN1
C1
F1 interspecific cross
H.grossessratus N1
C1
N1
C1
CMS- reccessive ms line
X
5. CGMS
H.petiolaris × H.annuus Repeated backcross of H.annuus
results in cms1 which is extensively
used mst in hybrid seed production
of sunflower all over the world
H.giganteus× H.annuus Cms3( S cytoplasm source)
H.annuus subspp
lenticularis × H.annuus
CV commander
Indiana 1
8. COTTON
Genetic Male Sterility (GMS):
In cotton, GMS has been reported in upland, Egyptian and arboreum cottons.
In tetraploid cotton, male sterility is governed by both recessive and
dominant genes. However, male sterility governed by recessive genes is used in
practical plant breeding
All three types of male sterility occurs (g mst,c mst,gc mst) in cotton
sixteen different genes in tetraploid cottons (13 in G. hirsutum and 3 in G.
barbadense) and two in G. arboreum have been identified for genetic male
sterility.
Sterility is conditioned by dominant alleles at five loci viz, MS4, MS7, MS10,
MS11 and MS12 by recessive allele at other loci viz. msl, ms2, ms3, ms13, ms14
(Dong A), ms15 (Lang A) and ms16 (81 A).
Two male sterile phenotypes viz. ms5ms6 and ms8ms9 are conditioned by
duplicate recessive factors.
G. hirsutum line Gregg (MS 399) from USA is the basic source of
GMS possessing ms5 ms6 gene for male sterility.
11. CMS System
In case of CMS, the originally discovered CMS sources involving G.
arboreum and G. anomalum cytoplasmic systems having interaction
with ms3 locus were not found effective or stable under different
environments.
The only stable and dependable CMS source under varied environment
was developed through the utilization of G. harknessii. The complete
genome of G.hirsutum was transferred into the G. harknessii cytoplasm.
A single dominant gene ‘Rf’ from G.harknessii is essential for fertility
restoration.
Fertility enhancer factor 'E' for this CMS restorer system was obtained
from a G.barbadense stock.
The harknessii system is reported to contribute to good agronomic
properties and attraction to honey bees.
12. Sources of Male sterility in Cotton
Source of ms cytoplasm Nuclear genome
G. anomalum, G. arboreum, G.
harknessii
G. hirsutum
G. anomalum, G. arboreum Heat sensitive , less stable
G. harknessii × G. hirsutum Stable cms all over the
environment
New sources of CMS
G. aridum Skovt. × G. hirsutum (D4)
G. trilobum × G. hirsutum CMS 8 (D-8)
G. sturtianum × G. hirsutum CMS-C1
New sources of CGMS
G. anomalum x G. thurberi Cg-mst
13. Mutation
G. arboreum, the first spontaneous male sterility mutant was identified in
variety DS-5
Chemical based male sterility
FW 450(Sodium B-Dichloro-iso-butyrate)
MH-30 (Maleic hydrazide)
Ethidium bromide
Male sterility based hybrid Production
GMS system. CPH2 (Suguna), First hybrid based on GMS released at
CICR, RS, Coimbatore
G. harknessii based cms with fertility restoration gene sources were used
in developing the hybrid CAHH 468 (PKV Hy-3).
14.
15. T cytoplasm S cytoplasm C cytoplasm
Discovered by Rogers(1944) M.T. Jenkins Beckett(1971)
Developed
from
OP Mexican
maize variety
Gold june
Teopod maize Brazilian Maize
Male sterility Stable Unstable Stable
HMT
Susceptibility
Yes No No
Type of mst Sporophytic Gametophytic Sporophytic
Fertility
restoration
Fr1,Fr2 - Fr4,Fr5, Fr6
CMS
T cytoplasmic mitochondria, HM T pathotoxin causes the
uncoupling of oxidative phosphorylation, inhibiton of oxogluterate
oxidation and causes irreverisble swelling
17. CGMS
Reported by Rhoades (1931) while working with
Peruvian source of maize cytoplasm
Interspecific crosses
Coix lacrymus-jobi X Z.mays
Euchlaena mexican X Z. mays
E.Perennis X Z.mays
Chemcial mst
GA, MH, Mendok, DPX 3778, Mo deficiency
18. A X B
(frfr) (FrFr)
ms mf
AB
(Frfr)
mf
Method of Hybrid seed production
Single cross Hybrid
T- cytoplasm required two Fr genes
C,S-cytoplasm requires one Fr genes
Most of the T cytoplasm posses one Fr gene
19. A X B
(frfr) (frfr)
ms mf
AB
(frfr)
ms
X C
(FrFr)
mf
ABC
(Frfr)
mf
Triple cross Hybrid
20. C X D
(frfr) (FrFr)
ms mf
CD
(Frfr)
mf
A X B
(frfr) (frfr)
ms mf
AB
(frfr)
ms
X
ABCD
1
(Frfr)
mf
1
(frfr)
ms
:
:
:
Double cross Hybrid
21. Simple hybrid with cms and
restoration
Maintainer line (B-line)
N, rfrf
N1
C1
xCMS line (A-line)
CMS, rfrf
N1
C1
Large amounts
of CMS line N1
C1
C2
x N2
Male line (C-line
N and RfRf
C1
Fertile F1 hybrid
CMS, Rfrf
23. CGMS
A1 Tift 23 A ( Most of the world hybrids contains
A1 Blood), Burton,1958
A2,A3 Not stable cytoplasm
A4 Derived from P.glacum subspecies monodii
Does not have effective restorer
Used in forage hybrid production
24.
25. Cytoplasmic male-sterile
Stamen (anther and filament) and pollen grains are
affected
It is divided into:
a. Autoplasmic
CMS has arisen within a species as a result of spontaneous
mutational changes in the cytoplasm, most likely in the
mitochondrial genome
b. Alloplasmic
CMS has arisen from intergeneric, interpecific or occasionally
intraspecific crosses and where the male sterility can be
interpreted as being due to incompatibility or poor co-operation
between nuclear genome of one species and the organellar
genome another CMS can be a result of interspecific protoplast
fusion
26. Genetic Male Sterility
GMS is governed by two genes either recessive or dominant
genes(Kaul,1988)
One more dominant gene is associated with development of male
sterility in B.napus type by means of transgenic male sterility
27. Cytoplasmic Male Sterility
1.Raphanus or ogu system
2.Polima or pol system
3.Shiga-Thompson or nap system
4.Diplotaxis muralis or mur system
5.Tournefortii (tour) system
6. Moricandia arvensis or mori system
7.Chinese juncea or jun system
17 systems are available, only difference is the use of male sterile
cytoplasmic sources differs for each system
Nap system– B.napuus cross b/w winter & spring var.
pol system – B.napus var polima
mur system--Diplotaxis muralis x B.campestris cv Yukina
tour system– B.juncea collections
28. Ogu system:
First discovered in Japanese radish (Raphanus sativus) by Ogura, 1968
B.napus genome was transferred into the back round of R.sativus (mst)
through intergeneric crosses followed by back crossing with B.napus.
CMS seedling under low temperature showed chlorosis , because
chloroplast of R.sativus is sensitive to cold, it is governed by cp-DNA ,
but mst is governed by mt DNA.
Protoplast fusion of R.sativus with B.napus carried out to have normal
green plants with ogu CMS characterisitics
This system now has been used for developing alloplasmic male sterile line
in B.juncea and B.campestris.
Ogu system:
29. B.napaus
F1 interspecific cross
xRhapanus
sativus
F1 Sterile
G-Rs
C-Rs
G-Bn
N-Bn
1/2G-Rs
1/2G-Bn
C-Rs
mftmst
Doubling by colchince
Fertile amphidiploid
1/2G-Rs
1/2G-Bn
C-Rs
mst
Development of Male sterile Brassica napus from Rhapanus sativus
31. Development of Alloplasmic Male sterile Brassica campestris
x
N-Bc
B.campestris
F1 interspecific cross
xG-Bn
S-Rs
G-Bct
N-Bc
1/2G-Bn
1/2G-Bc
S-Rs
mftmst
G-BC
S-Rs
BC4
G-BcG-Bc
Male sterile B.napus
36. Development of Male sterility
Genome transfer
S cytoplasm is in the genome of fr genes
Unreduced Gamete Production
S.tuberosum (2x) × S.tuberosum (4x)
(2x)
F1 (4x)
Protoplast Fusion
S cytoplasm is retained
Unreduced (2x) (2x)
F1 (4x)
40. Maintenance of ms determinant in populations. Maternal
inheritance (mitochondira) the male sterility determinant (red
cytoplasm) and the male fertility determinant (green cytoplasm)
are equally transmitted to the next generation