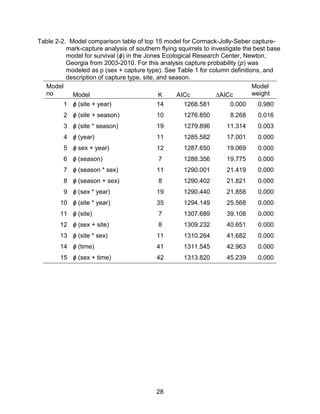
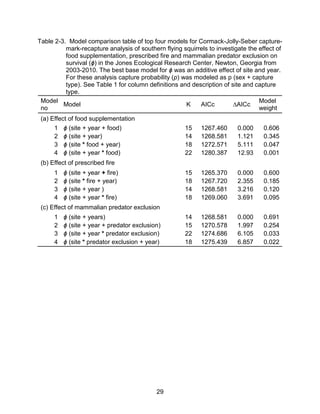
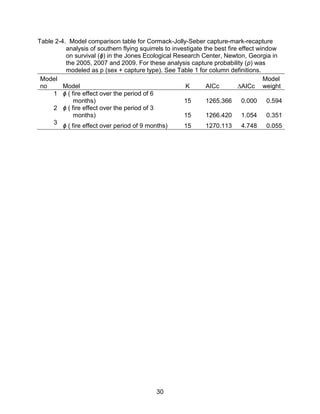
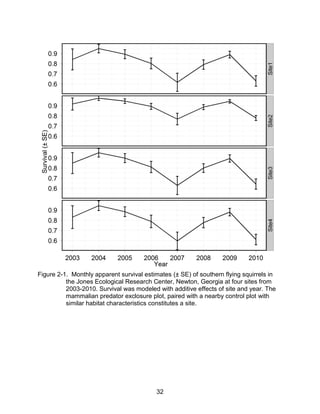
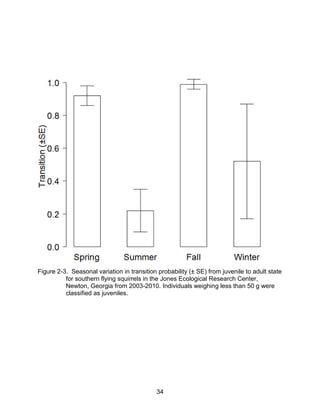
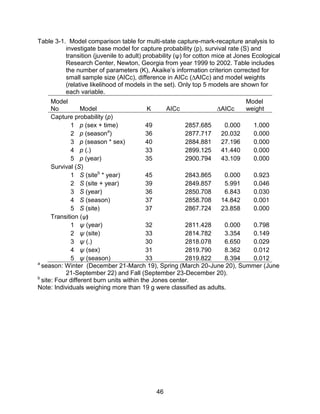
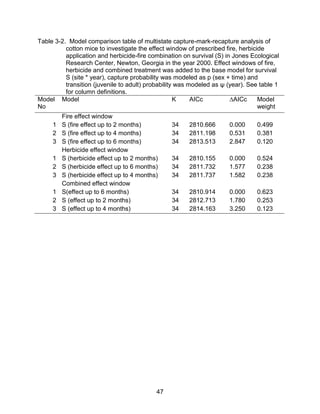
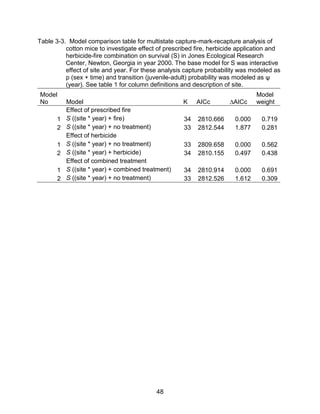
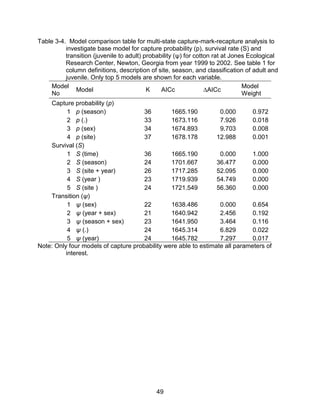
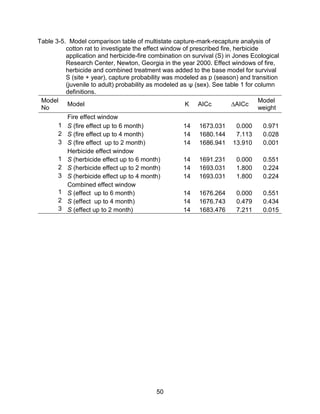
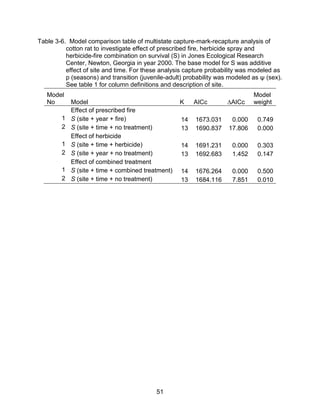
This thesis examines the effects of food, fire, herbicide, and predation on small mammal populations in a longleaf pine ecosystem through capture-mark-recapture studies of the southern flying squirrel and cotton mice/rat. For southern flying squirrels, apparent monthly survival was estimated to be 0.84 on average, with higher survival for adults than juveniles. Prescribed fire had a positive effect on survival, increasing it for a period of up to 6 months after a burn. Food supplementation and predator exclusion did not significantly influence survival. Studies of cotton mice and rats found their survival was impacted by prescribed fire and herbicide application.