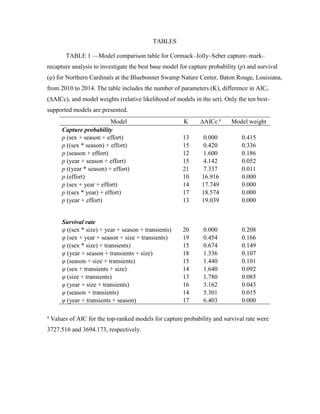
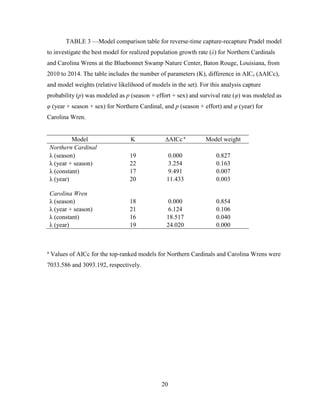
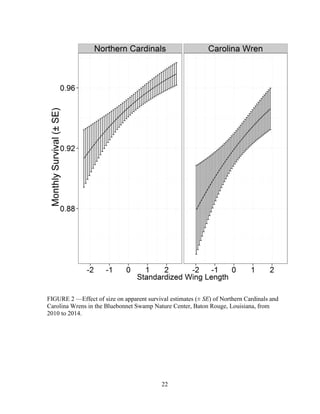
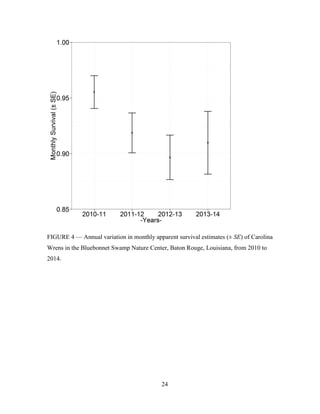
This thesis examines population dynamics of Northern Cardinals and Carolina Wrens in an urban forest fragment using capture-mark-recapture analysis of data collected from 2010-2014. The study aims to determine if the forest fragment acts as a safe refuge or ecological trap for these species. Capture-mark-recapture was used to estimate survival, recruitment, and population growth rates as a function of sex, age, body size, season, and year. Results will help understand how these species persist in small habitat fragments and identify life stages most impacting population persistence to inform conservation efforts.