Download to read offline



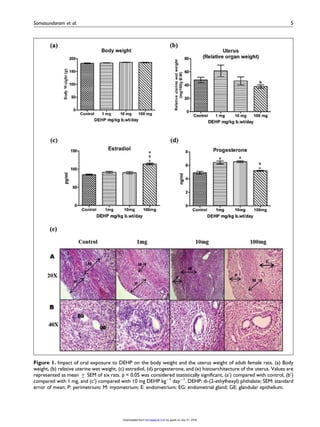
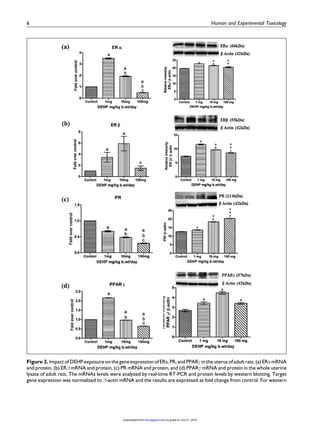



This study examined the effects of di-(2-ethylhexyl) phthalate (DEHP) exposure on the uterus of adult female rats. Rats were orally administered DEHP at doses of 0, 1, 10, and 100 mg/kg body weight daily for 30 days. Key findings include: 1) Serum estradiol levels were unchanged in the 1 and 10 mg groups but marginally increased in the 100 mg group. Progesterone levels increased in the 1 and 10 mg groups. 2) Histological examination found structural abnormalities in the uterus such as decreased diameter and thinning of layers in the 10 and 100 mg groups. 3) mRNA expression of estrogen receptor alpha decreased in the 100 mg



































































