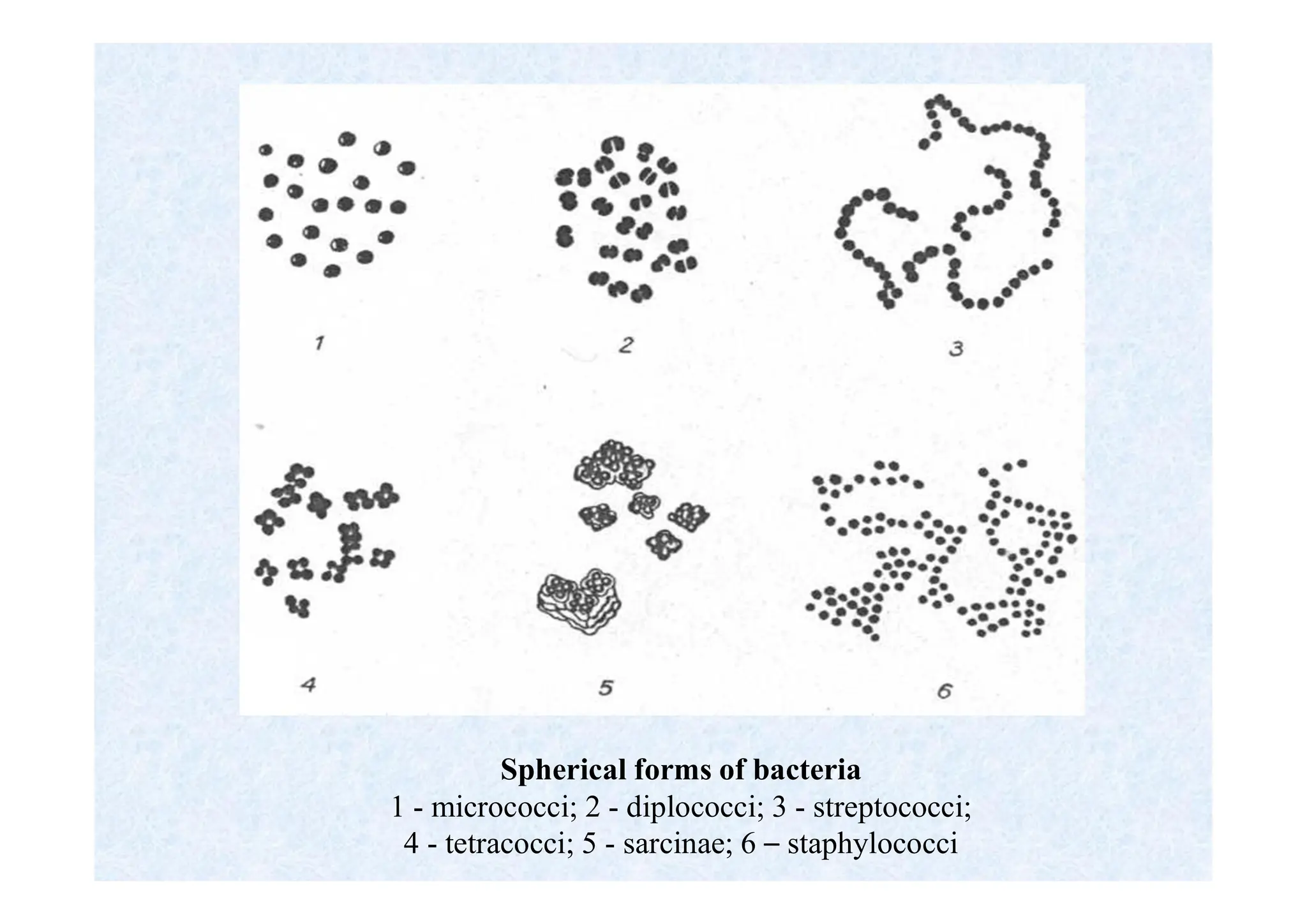
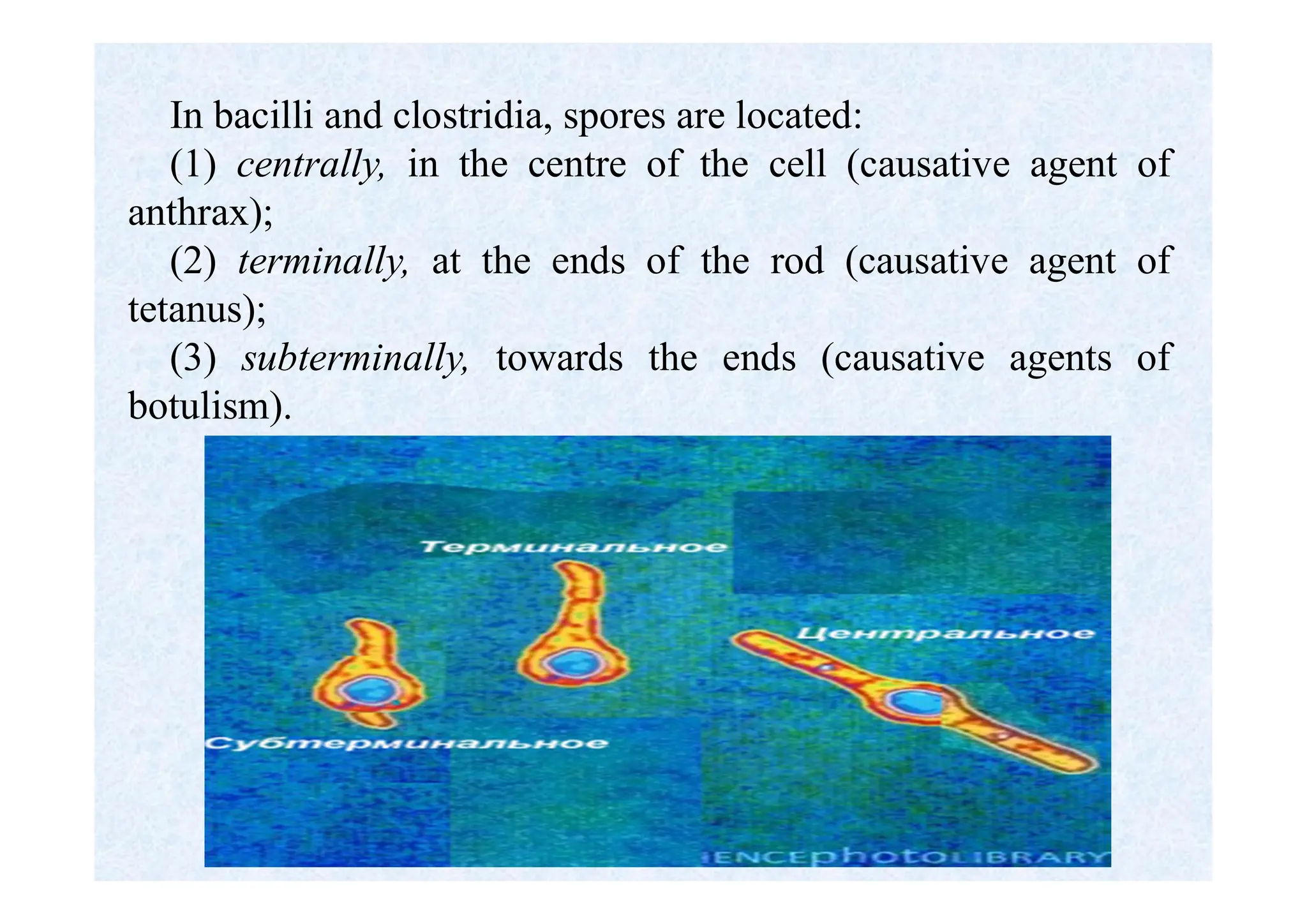
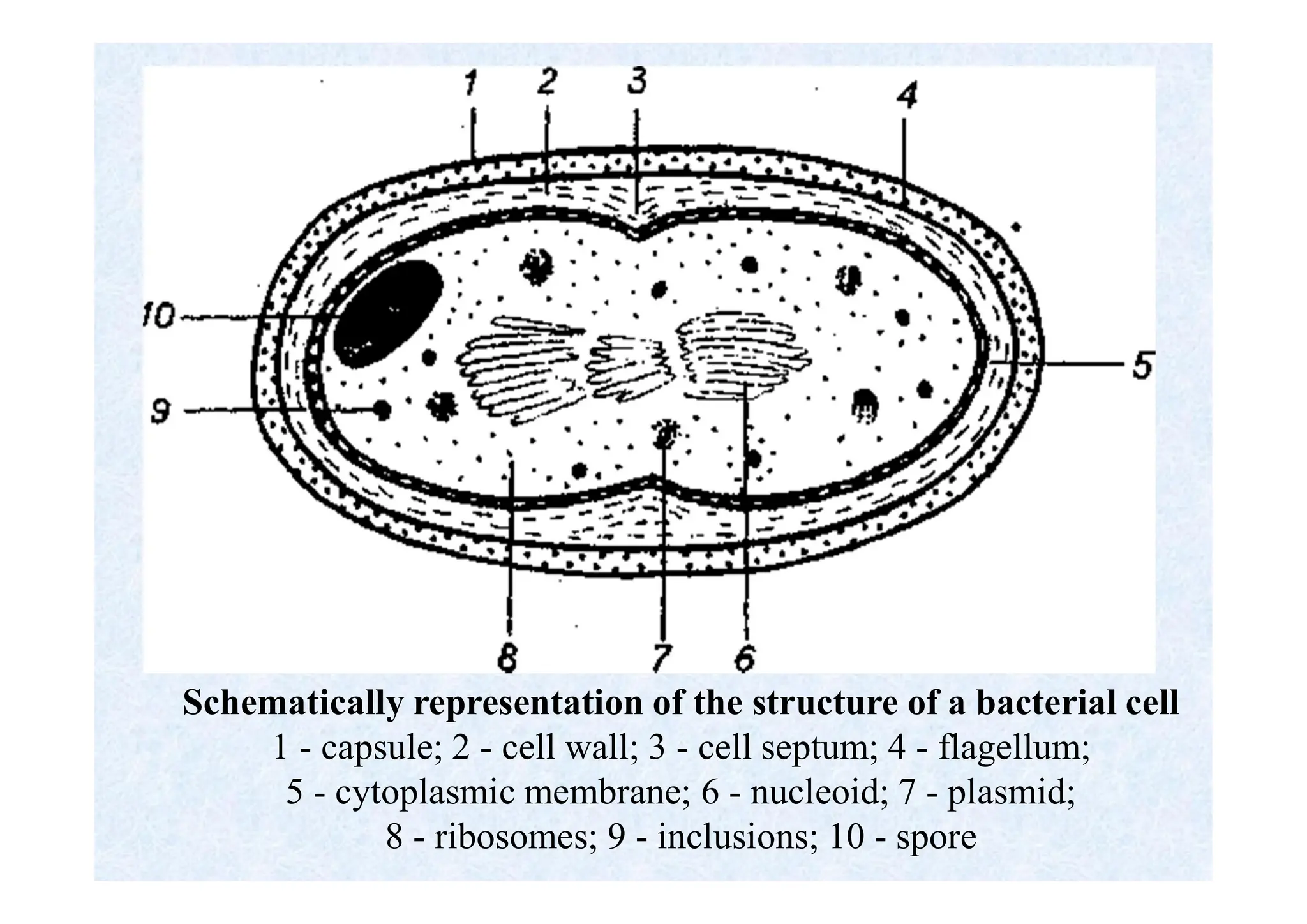
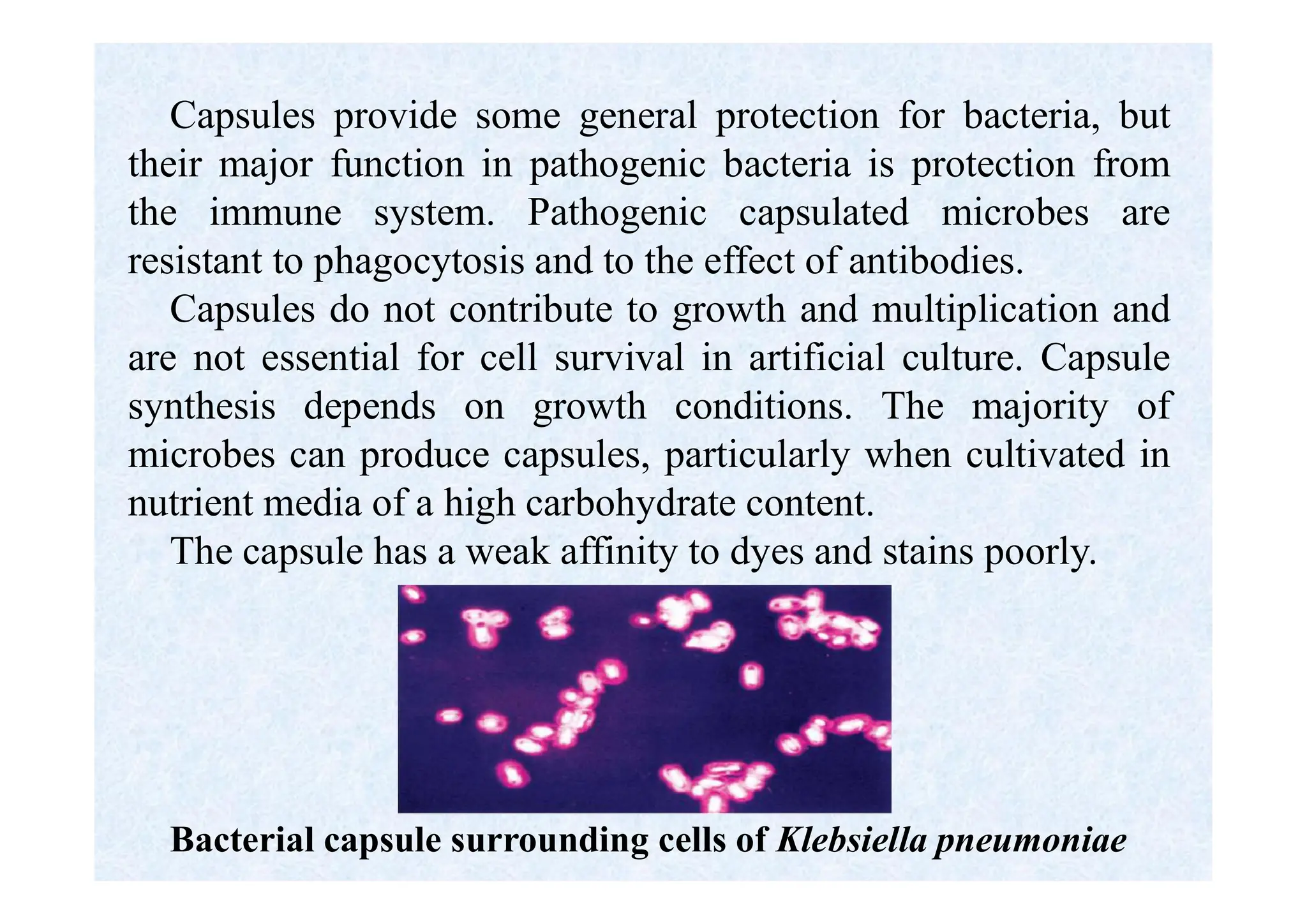
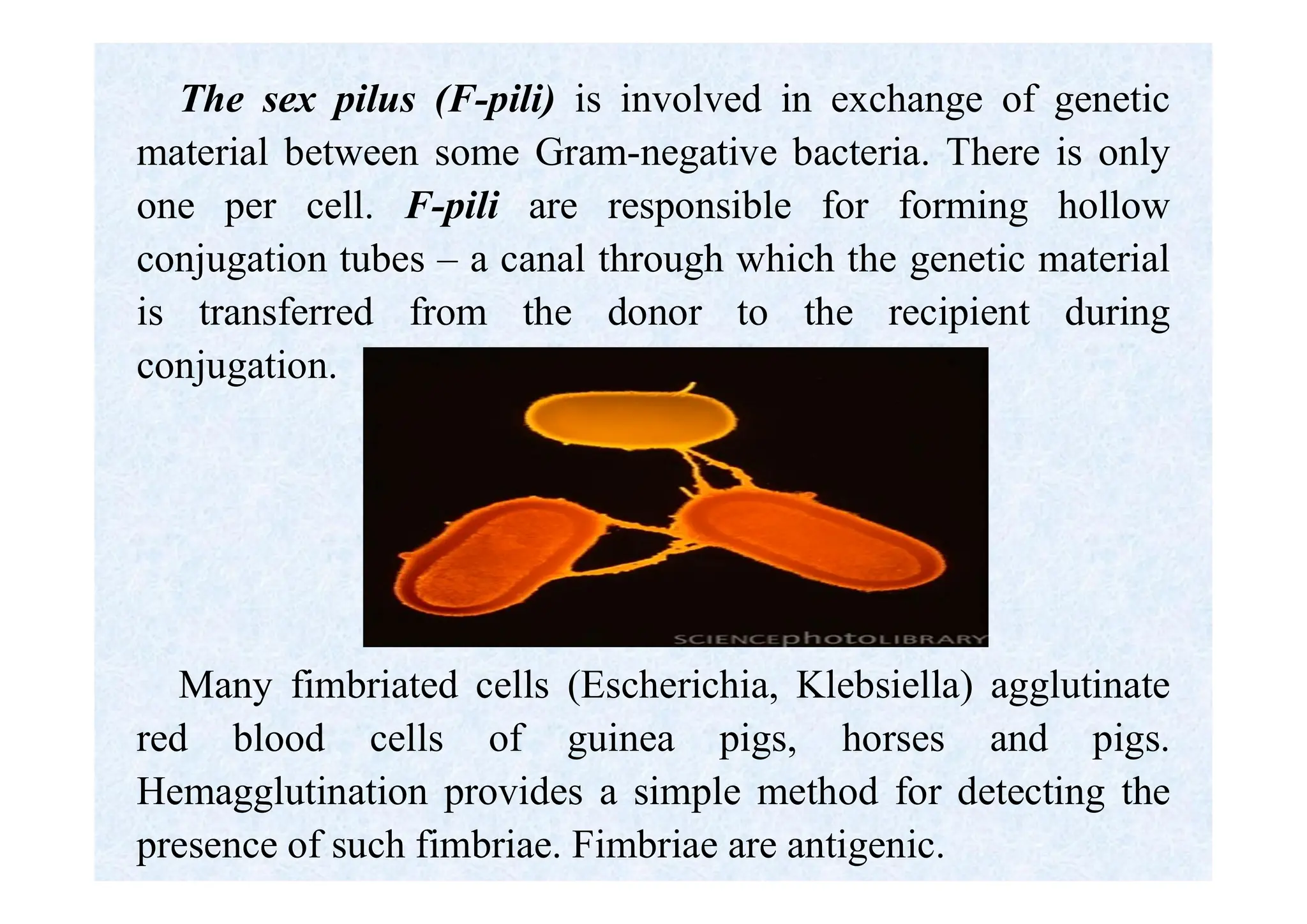
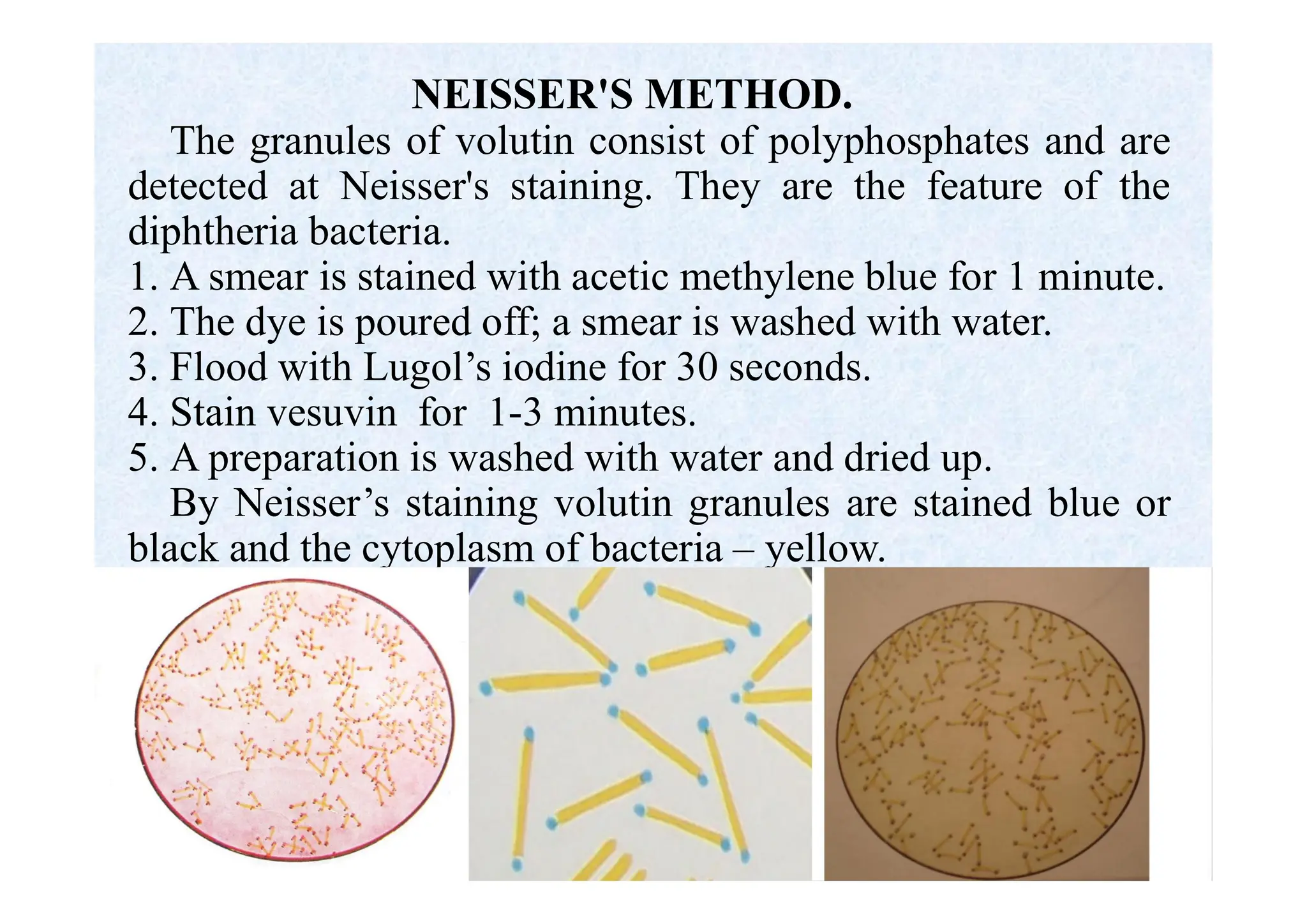
Morphology of
microorganisms.
Bacterial ultrastructure.
Complex staining methods.
Lecturer:
Associate Professor of the MicrThe Subject and Problems of Microbiology.
Microbiology (Gk. mikros small, bios life, logos science Classification and Morphology of MicroorganismsTaxonomy and classifications of microbes