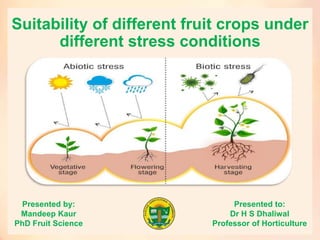
Suitability of different fruit crops under different stress conditions
•
7 likes•1,580 views

Suitability of different fruit crops under different stress conditions

Recommended
Recommended
More Related Content
What's hot
What's hot (20)
Similar to Suitability of different fruit crops under different stress conditions
Similar to Suitability of different fruit crops under different stress conditions (20)
More from _mk_ saini
More from _mk_ saini (20)
Recently uploaded
Recently uploaded (20)
Suitability of different fruit crops under different stress conditions
- 1. Suitability of different fruit crops under different stress conditions Presented by: Mandeep Kaur PhD Fruit Science Presented to: Dr H S Dhaliwal Professor of Horticulture
- 2. What is Stress ? • Stress in biology is any change in environmental conditions that might reduce or adversely change plant growth and development (Levitt, 1972). • Stress is a condition caused by the factors that tends to alter an ‘equilibrium’ (Strasser, 1998). • Jacob Levitt- Father of plant stress physiology. • One stress is often combined with typical set of other stresses, so they do not act individually in nature. • Stresses may be additive or mitigating each other.
- 4. Physiological adaptations under different stress conditions • Plants can modify their life cycles: Eg temperate fruits • Phenotypic changes in leaf structure and behaviour are important stress responses (changes in leaf area, leaf orientation, trichomes, and the cuticle). • Under conditions of water, heat, or salinity extremes, leaves may be narrower or may develop deeper lobes during development. • Plants change stomatal activity in response to dehydration stress. • Plants adjust osmotically to drying soil by accumulating solutes. • Salt exclusion: Prevents the entry of salts into the vascular system. Eg Vitis champini (Cl-ion excluder) and Cleopatra mandarin • Salt elimination: Salt-secreting glands and hairs actively eliminate salts, thus keeping the salt concentration in the leaves beneath a certain threshold. • Heat shock proteins can be induced by different environmental conditions: In response to sudden 5 to 10°C increases in temperature, plants produce a unique set of proteins referred to as heat shock proteins (HSPs). • Many plants have the capacity to acclimate to cold temperature: Several specialized plant proteins, termed antifreeze proteins, limit the growth of ice crystals through a mechanism independent of lowering of the freezing point of water.
- 5. Plant Response to Different Stress • Phytohormones like ABA, SA, JA, and ET accumulates and increase in plant tolerance to various stress. • ABA is responsible for plant defense against abiotic stresses such as drought, salinity, cold, heat stress and wounding. • SA, JA and ET play major roles in response to biotic stress conditions as their levels increase with pathogen infection. • Role of different proteins in stress response: Heat Shock Proteins, Anti free proteins, DREB proteins, LEA proteins and PR proteins. • Osmoprotectants: Osmotin accumulates in salt adapted cells, the overexpression of osmotin induces proline accumulation (a compatible solute).
- 7. A scheme representation of abiotic stress at the cellular level
- 8. Biotic stress at the cellular level
- 9. A scheme for the interaction interface and overlapping signaling pathways of abiotic and biotic stress at the cellular level
- 10. A scheme for the effects of abiotic and biotic stress at the plant level
- 11. Approaches for building combined abiotic and biotic stress Tolerance in plants
- 12. Beyond Chemical Triggers: Evidence for Sound-Evoked Physiological Reactions in Plants • Naturally occurring and artificially generated sound waves contribute to plant robustness. • New information is emerging about the responses of plants to sound and the associated downstream signaling pathways. • This paper provide an overview of the latest findings, limitations, and potential applications of sound wave treatment. • No doubt, chemical triggers improves plant health by enhancing plant growth and resistance. • But, Sound wave treatment is a new trigger to help protect plants against unfavorable conditions and to maintain plant fitness by modulating physiological traits. It shows adaptive advantage in plants. Jung et al (2018) Korea Research Institute of Bioscience and Biotechnology, Daejeon, South Korea,
- 13. Sound production and perception in plants (A) Sound production: Plants produce sound vibrations in their xylem via the generation of tension in the xylem vessel when its diameter decreases. Additionally, gas bubbles produced in xylem vessels during transpiration may produce sound. (B) Sound Perception: Although there are no visible alterations, transcriptional and translational changes occur in plants exposed to sound vibrations. But, levels of stimulus responsive, signaling-related, redox homeostasis, and defense related transcripts are changed in sound-exposed plants. However, the specific organs or proteins used for sound perception have not yet been identified. PRODUCTION (SPEAKING) AND PERCEPTION (LISTENING) OF SOUND IN PLANTS Can Plants Make Sound Vibration? Can Plants Respond to Sound?
- 14. Responses of plants to sounds of different frequencies and magnitudes.
- 15. 1. Sound promotes growth by increasing IAA and GA. 2. Sound pretreatment enhances plant immunity against subsequent pathogen attacks by activating the plant defense hormones SA and JA. 3. Sound treatment triggers drought tolerance by changing the elasticity and flexibility of the cell wall, which affects the ability of plants to absorb water. 4. Ethylene production is delayed by down-regulation of E biosynthesis and expression of signaling-related genes. 5. Sound treatment increases expression of photosynthesis-related genes, such as those encoding fructose 1,6-bisphosphate aldolase and the rubisco small sub-unit, and may induce CO2 fixation Sound waves as a plant stimulant and protectant. Artificial sound treatment can elicit various effects in plants.
- 16. Perspectives And Remaining Questions Sound represents a potential new trigger for plant protection 1. The discovery of an organ or a specific protein in plants that recognizes sound waves would help us maximize the effectiveness of the use of sound treatment in field trials. 2. Second, technology used to engineer sound quality, such as the fine turning, modification, and mixing of sounds, must also be improved to facilitate its use for sound-mediated stress relief and increased plant growth. 3. Third, the analysis of plant biomarkers such as Pathogenesis-Related 1 protein (PR1) (for systemic acquired resistance) will help scientists optimize sounds to maximize sound-specific plant stress relief. 4. Side effects of sound waves. Humans can differentiate and recognize sounds ranging from 20 to 20,000 Hz. If the sound vibration used to treat plants causes damage to animals, humans, or microbes after long-term exposure, a detailed examination and evaluation of the effects of various exposure times and high- frequency (e.g., above 20,000 Hz) will be required. • If the proper electric power supply, speakers, and associated sound-generating equipment are utilized sound treatment can constitutively be applied for long periods of time without additional input. • This unique setup, which has not been tested before, awaits your next experiment.
- 17. SUITABILITY OF DIFFERENT FRUIT CROPS UNDER SALT STRESS CONDITIONS
- 18. Salinity Tolerance of Some Grapevine Cultivars as Affected by Salt Creek and Freedom Rootstocks • Objective: To study variability in salt tolerance of varieties (Flame seedless, Early sweet, Superior seedless and Red Glob) grafted on two rootstocks (Salt creek, Freedom) compared with own rooted vines on growth and nutrient content under varying levels of salinity treatments (1000, 2000, 3000 ppm). • Results: Vine growth significantly decreased with the increase in salinity levels in all vines (Reason: Increasing salt concentration significantly reduced the N, P and K contents in the leaf petioles). • At the highest salinity level (3000 ppm), the survival vine percentage was zero % for own rooted vines, except, the own rooted Flame seedless that recorded 50 percent of vine survival. • Among the scion-rootstock vines, about 70-80% vine survival was observed with the vines grafted on Salt creek and 60-65% in vines grafted on Freedom rootstock. • Conclusion: All cultivars grafted on Salt creek rootstock were more tolerant to salinity. National Research Center, Egypt Desouky et al (2015)
- 19. Effect of saline irrigation water on vines height (cm)
- 20. Effect of saline irrigation water on survival vines percentage
- 21. Relative Salt Tolerance of Different Grape Rootstocks to Different Chloride Salts • Objective: To study the variability in salt tolerance of grape rootstocks (viz., Dogridge, Salt Creek, RS-19, SO4 and 1613-C) to different chloride salts viz., NaCl, KCl, MgCl2 and CaCl2 salts) applied at 4 meq, 8, 16 and 32 meq/lt concentrations. The mean of different concentration of salts was used for salt tolerance assessment. • Observations: The relative salt tolerance was assessed based on growth parameters, dry weight of root to shoot ratio and also on the ability of rootstocks to limit uptake of Na and Cl ions. • Results: Highest root growth parameters measured in terms of root number (59.79), length (42.9cm) and dry weight (24.51g) and also dry weight of root to shoot ratio (0.6) was recorded in Dogridge. • High chloride exclusion ability represents the salt tolerance. Significantly lowest chloride content was recorded with Dogridge (0.87%) rootstock whereas the highest chloride content was recorded with Salt Creek (1.14%). • The Cl accumulation in leaf was significantly higher when applied in the form of NaCl followed by KCl. • Conclusion: Due to low Cl־ content in leaves, high dry weight of root shoot ratio and high vigour, Dogridge is more tolerant to salinity than other rootstocks studied. Saritha et al (2017)Grape Research Station, Rajendranagar, Hyderabad
- 22. Effect of varying chloride salts on root parameters in different grape rootstocks
- 23. Effect of different chloride salts on Cl content (%) in different grape rootstocks Chloridecontent(%)
- 24. Adaptation of grapevine (wild and rootstocks) to salinity • Objective: To establish a possible correlation between salt resistance and mineral content of plants after salt treatments applied in two different culture systems, in vitro culture of meristematic structures and potted plants, respectively. • EXP. 1: The shoot tips were harvested from potted plants and the apexes were cultured on solidified MURASHIGE and SKOOG (1962) medium, supplemented with 17 (V1), 34 (V2) and 43 (V3) mM NaCl. The medium without NaCl content was used as a control for the test. • EXP. 2: Potted plants were watered every two days with solution of 51, 68, 102 mM NaCl, and also without NaCl (as control) for 30 d. Five rootstocks SO4 Kober 5BB 140Ru Fercal Riparia Gloire Six different wild plants from Vitis vinifera L. ssp. sylvestris Vs 1 Vs 10 Vs 9 DM Vs 3 EB Letea 1 Letea 2Popescu et al (2015)Romania
- 25. The influence of NaCl on explants evolution and shoots development, after 5 passages on the same media composition. Example for rootstock 'Fercal 242' and wild plant Letea 1. From the left to the right: control (M), 17 (V1), 34 (V2) and 43 (V3) mM NaCl.
- 26. The effects of different salt concentrations on plant development; potted plants after 15 treatments. From the left to the right: control (M), 51 mM (V1), 68 mM (V2) and 102 Mm (V3) NaCl.
- 27. Effect of saline treatments on nutrients tissue content
- 28. In the potted conditions, the tested rootstocks reacted as “Na includes” while in vitro tests the plants belonging to wild populations reacted as “Na excluders". In vitro conditions favored absorption and retention of K and Na elements in vegetative structures, while potted treatments increased the absorption of P. Different NaCl resistance mechanisms are involved in vitro plant material and potted plants subjected to salt stress. On the whole, the wild grapevine individuals, in comparison to the rootstocks, expressed a higher ability to adapt to the salt stress in both type of treatments. Effect of saline treatments on nutrients tissue content
- 29. Grape Rootstock Response to Salinity, Water and Combined Salinity and Water Stresses A field experiment was conducted at the USDA ‐ California from June 2013 to Sept 2017 to evaluate the effect of 4 irrigation water salinity levels and three water treatments on three grape rootstocks purported to be salt tolerant. Objectives: 1. The salt tolerance (yield) response of Cabernet Sauvignon grafted on to three rootstocks. 2. Response to irrigation quantities, from meeting crop evapotranspiration (ETc) to application of 80% and 60% of crop evapotranspiration (ETc). 3. Vine and fruit yield response to combined effects of salt stress and irrigation quantity. The crop evapotranspiration (ETc) was measured by lysimeters USDA‐Agricultural Research Service, U.S. Salinity Laboratory in Riverside, California Suarez et al (2019)
- 30. 4 irrigation water salinity levels S 0 = 0.7 dS m−1 water S 1 = 1.7 dS m−1 water S 2 = 2.7 dS m−1 water S 3 = 3.7 dS m−1 water Cabernet Sauvignon scions grafted onto SC represents Salt Creek rootstock Ru represents 140 Ruggeri rootstock SG represents St. George rootstock Three water treatments D 0 = 100% ETc Two 1.0 L h−1 and one 0.5 L drippers D 1 = 80% Etc Two 1.0 L h−1 drippers D 2 = 60% ETc One 1.0 L h−1 and one 0.5 L h−1 drippers,
- 34. Conclusion • The 140 Ruggeri scion was the top producer across all treatments including control, followed by Salt Creek, with St. George significantly less productive than 14 Ruggeri across all treatments. • No significant rootstock differences related to water stress was found. • Reductions in yield was observed under reduced water applications. • Under saline conditions, the reduced water applications result in reduced leaching and thus higher soil salinity. • Thus, it is recommend that water application should not be reduced under salinity conditions.
- 35. Salinity Soil Effects on Yield, Fruit Quality and Mineral Composition of Superior Seedless Grapevines Grafted on Some Rootstocks • Objective: To evaluated superior seedless grape cultivar grafted on Paullson 1103, Salt Creek and freedom grapevine rootstocks to soil salinity compared to own rooted superior seedless cultivar. • Methodology: Vegetative growth parameters, chemical analysis, yield and berry characteristics and antioxidant isozymes electrophoresis was estimated. • Results: Superior grape cultivar grafted on three rootstocks gave the best results as compared to own rooted superior grape cultivar for three consecutive seasons. • Superior seedless grape cultivar grafted on Salt Creek rootstock had the highest percentage of bud burst and fruitful buds, improved the best vegetative growth parameters, increment depth of the roots and their distribution in the soil profile. • Cane content of total carbohydrates, leaf content of total chlorophyll and mineral content were increased and leaf prolin content, sodium and chloride were reduced. Additionally, it had a positive impact on the yield and berry quality attributes. • Conclusion: Superior seedless grafted on Salt Creek, Paullson 1103 and freedom grape root stocks were more tolerant to soil salinity than superior seedless cultivar on own rooted root stock, vines grafted on Salt Creek was the most tolerant one. Agriculture Research Center, Giza, Egypt Ahmad (2016)
- 36. Effect of grafting superior seedless cv. on some rootstocks under saline condition on minerals content during (2012, 2013 and 2014) seasons
- 37. Responses of Cherry Plant Grafted onto CAB-6P, MaxMa 14 and Mazzard Rootstocks to Short Term Salinity • Objective: To investigate morphological and physiological responses of a cherry cultivar (0900 Ziraat) grafted onto three rootstocks (CAB-6P, MaxMa 14 and Mazzard) to 35 mM NaCl stress. • After one month, salt stress decreased plant growth of cherry plants. Rootstock, scion and shoot diameters and shoot length significantly decreased as a result of restriction in nutrient uptake due to the salinity that limits photosynthesis consequently decreases plant tissue extension. • Results: In the current study, the growth of Mazzard rootstock was less damaged & showed higher tolerance to short term salt stress as compared to CAB- 6P and MaxMa 14. • Reason: Lower depression in plant growth, less decrease in chlorophyll content and more stability in cell membranes as compared to other rootstocks. • Conclusion: In the short term, cherry can be considered salt tolerant. • But, long term responses of cherry trees to salt stress may be different due to the salt accumulation in leaves. Aras & Esitken (2019)Selcuk University Konya, Turkey
- 38. Effects of salinity on physiological responses of cherry plants * Relative chlorophyll value was measured with a Minolta SPAD-502 chlorophyll meter. * Stomatal conductivity and leaf temperature measurements by leaf porometer. * Membrane permeability by the procedure of electrolyte leakage based on Lutts et al (1996). * Leaf relative water content (LRWC) by the procedure of Smart and Bingham (1974).
- 39. Improved salt-tolerance in Citrus macrophylla mutant rootstocks • Mutagenesis by gamma rays was used to develop salt tolerant mutants. • In previous works, five mutants (MM3B, MM4B, MM5B, MM2A and MM3A) from Citrus macrophylla, obtained through the exposure of seeds to gamma radiation (129 Gy), were selected for their ability to cope with salt stress in vitro. • In the present experiment, the selected mutants, along with Citrus macrophylla shoots as control, were submitted to 60mM NaCl (MM3B, MM4B and MM5B mutants) or to 80mM NaCl (MM2A and MM3A), in their respective culture media. • After 8 weeks of exposure, proliferation rates, shoot length and damaged leaves were recorded to assess the effects produced by salinity in the culture medium. Jiménez and Tornero (2019)Murcia, Spain
- 40. Control in vitro MM2A mutant shoot (A) MM2A mutant shoot exposed to 80mM NaCl (B) ‘Alemow’ mutant shoot exposed to 60mM of NaCl (C) MM5B mutant shoot exposed to 60mM NaCl (D) MM3B mutant shoot exposed to 60mM NaCl (E) MM3A mutant shoot exposed to 80mM NaCl (F) MM4B mutant shoots exposed to 60mM NaCl (G)
- 41. Effect of NaCl exposure on the NO ³־ content in ‘Alemow’ and the mutants MM3B, MM4B, MM5B, MM2A and MM3A. NO ³־ increased in the mutants, as compare to ‘Alemow’. This could be the key to resisting stress, since sustaining the supply with essential nutrients, such as nitrate, is believed to enhance plant salinity tolerance. This is because N is involved in the biosynthesis of many important compounds and its lack would cause a serious imbalance, resulting in reduced growth and lower yield.
- 42. Effect of NaCl exposure on the chlorophyll content in ‘Alemow’ and the mutants MM3B, MM4B, MM5B, MM2A and MM3A. Before salt stress (60mM), higher photosynthetic capacity was present, to produce and store higher amounts of photosyntetates for use in stress conditions. This capacity, along with the other genetic adjustments involved in mutation, could have allowed these mutants to improve proliferation and growth despite the leaf damage, as indicated by the productivity records in salinity.
- 43. Effect of NaCl exposure on the proline content in ‘Alemow’ and the mutants MM3B, MM4B, MM5B, MM2A and MM3A. Proline has been reported to act as a protector that is able to overcome abiotic stresses through the balance of the osmotic potential. But, here, the relation between its accumulation and stress is controversial and remains unclear.
- 44. Effect of NaCl exposure on the soluble sugars in ‘Alemow’ and the mutants MM3B, MM4B, MM5B, MM2A and MM3A. Soluble sugars & starch maintains the osmotic adjustment in the plant. Here, soluble sugars content augmented in MM5B explants from control to salinity, which shows better response to the osmotic stress in this mutant.
- 45. Effect of NaCl exposure on the starch in ‘Alemow’ and the mutants MM3B, MM4B, MM5B, MM2A and MM3A. • Starch increases in salinity conditions in order to regulate osmotical disequilibrium in the plant. • Only MM3B explants activated this mechanism since the levels of starch augmented in salinity, in contrast to what happened in the rest of the mutants and ‘Alemow’. • However, starch did not seem to play any central role in the regulation of the mutants since the performance of MM3B did not stand out of the rest of the mutants.
- 46. Conclusion • In the saline conditions assayed, all the mutants showed better productivity than Citrus macrophylla. • Furthermore, MM5B showed less leaf damage and higher proline levels in the saline stress conditions than Citrus macrophylla. • NO3־ accumulation of was also higher in mutant plants than in Citrus macrophylla in the saline conditions, which conferred an additional advantage against stress.
- 47. Evaluation of MM106 and Omara apple rootstocks for salt tolerance in vitro • Objectives: To study the possibility of using in vitro shoot culture in rooting stage to evaluate salt tolerance of two apple rootstocks (MM106 and Omara). • Single shoots were cultured on MS medium as a rooting media supplemented with 0, 20, 40, 60, 80 and 100mM NaCl for 8 weeks. • Results: All rooting parameters (rooting percentage, roots number, roots length) and plantlets growth (height of rooted shoot) decreased as salt level increased in culture medium, with the reductions generally greater for Omara than MM106 rootstock. • Also NaCl effect resulted in plantlet necrosis and a reduction in total chlorophyll content of both rootstocks. • However, plantlets of MM106 showed less relative root and shoot growth reduction under salt stress compared with Omara, therefore, it appeared to be more salt tolerant in vitro than Omara. Abdel-Hussein (2006)University of Kufa, Iraq
- 48. • Rooting percentage decreased due to inhibitory effects of salt on the metabolic activities which associated with cell division, differentiation and elongation and these processes companied with adventitious root initiation on the base of microcuttings. • Endogenous auxin (IAA) and rooting co-factors levels reduced, which might lead to reduce root initiation, and these substances (especially IAA) play an important regulatory role in root formation and their development Effect of NaCl on rooting of MM106 and Omara apple shoots in vitro after 8 weeks
- 49. Effect of NaCl concentrations on root characters 0f MM106 and Omara apple rootstocks during rooting stage.
- 50. • Biosynthesis of chlorophyll was generally inhibited by the depressive effect of stress. • Under stress conditions, absorption of some ions (Mg and Fe) reduced, which were involved in the chlorophyll formation. • It further leads to chlorophyll suppression in leaves or an increase of some growth inhibitors, such as E or ABA production which enhances senescence. Efects of NaCl on height and total chlorophyll content of MM106 and Omara apple rootstocks plantlets after 8 weeks in culture
- 51. The enhancement of tolerance to salt and cold stresses by accumulating salicylic acid content via the cytosolic malate dehydrogenase gene in transgenic apple plants • Objective: To characterize the role of an apple cytosolic malate dehydrogenase gene (MdcyMDH) in the tolerance to salt and cold stresses and investigated its regulation mechanism in stress tolerance. • Material: ‘Gala’ apple in vitro shoot cultures were used for the gene expression assays, genetic transformation and other analyses. The ‘Gala’ cultures were grown on MS subculture media. • Three-year old pot-cultured WT and transgenic apple trees were used for salt treatment. • Salt stress treatments, that is at 50-mM NaCl for 10 days and subsequent 100- mM NaCl for 9 days. • At 8 °C for 7 days and subsequent 0 °C for 7 days in a 14-h light photoperiod. Wang et al (1997) Shandong Agricultural University, Tai-An, Shandong, China
- 52. STRESS TREATMENTS 1. NaCl (19 days): 50-mM NaCl for 10 days and subsequent 100-mM NaCl for 9 days. 2. Cold (14 days): At 8 °C for 7 days and subsequent 0 °C for 7 days in a 14-h light photoperiod. • Contents of chlorophyll (a + b) of the leaves from the transgenic apple cultures (WT, Line 5 and Line 7) under normal and stress conditions of NaCl & cold treatment • Gene expression analysis was done.
- 53. Conclusion • The MdcyMDH transcript was induced by mild cold and salt treatments, and MdcyMDH-overexpressing apple plants possessed improved cold and salt tolerance compared to wild-type (WT) plants. • A digital gene expression analysis revealed that MdcyMDH overexpression largely altered some biological processes, including hormone signal transduction. • The transgenic plants produced a high H2O2 content, but a low O2- production rate was observed compared to the WT plants. • The transgenic plants accumulated more free and total salicylic acid (SA) than the WT plants under normal and stress conditions. • Taken together, MdcyMDH conferred the transgenic apple plants a higher stress tolerance by producing more ROS and increasing the SA level. • MdcyMDH could serve as a target gene to genetically engineer salt- and cold-tolerant trees.
- 54. SUITABILITY OF DIFFERENT FRUIT CROPS UNDER WATER STRESS CONDITIONS
- 55. Physiological and biochemical responses of young olive trees (Olea europaea L.) to water stress during flowering • Objective: To examine physiological and biochemical changes in three Moroccan varieties (Moroccan Picholine, Menara and Haouzia) of young olive trees (Olea europaea L.) grown under three different water regimes. • T0 (severe water stress ): Another 12 pots were not irrigated during the water treatment period. • T50 (moderate water stress ): Twelve pots received half the water needed to maintain the soil at field capacity. • T100 (control; Well watered plants): Twelve pots of each cultivar were watered twice a week to field capacity (≈800 mL) and represented the control and well- watered plants (T100). • Leaf relative water content (RWC), water potential (Ψw), transpiration rate (E), stomatal conductance (gs), maximum quantum efficiency of PSII (Fv/Fm), the contents of total chlorophyll (TCC), proline (ProC) and soluble sugars (SSC) were measured at the flowering stage during three growing seasons (2015, 2016 and 2017). Yamani et al (2018)Sidi Mohamed Ben Abdellah University, Morocco
- 56. Leaf relative water content (RWC), water potential (Ψw), transpiration rate (E), stomatal conductance (gs), total chlorophyll content (TCC), maximum quantum efficiency of PSII (Fv/Fm), proline content (ProC) and soluble sugars content (SSC). • Among the growing seasons, 2015 flowering displayed the lowest values for RWC, Ψw, E, gs, TCC and Fv/Fm, and the highest for ProC and SSC. (Temperatures was high in 2015. Under such conditions, olive plants can reduce excessive water loss by closing their stomata and accumulating osmolytes to facilitate the retention of water and prevent membrane damage). • Among plant varieties, no significant differences were observed. All are suitable due to high chlorophyll content and low proline accumulation). • Correlation studies highlighted significant associations between most parameters. • Positive relationships were found between RWC, Ψw, E, gs, Fv/Fm and TCC, while all of these parameters were negatively linked to ProC and SSC. Severe Moderate Control
- 57. Leaf mechanisms involved in the response of Cydonia oblonga trees to water stress and recovery • Objective: To identify the strategy by which quince plants cope with water stress and to further elucidate the resistance mechanisms developed in response to water stress and during recovery. • Old quince trees (cv. BA-29) were subjected to two irrigation treatments: • Control (T0) plants were drip irrigated (105% ETo) to ensure non limiting soil water conditions. • T1 plants were irrigated at the same level as used in T0, except that irrigation was withheld for 42 days during the linear fruit growth phase (i.e. 209–251 days of the 2016 year ; DOY) , after which irrigation returned to the levels of T0 (recovery period i.e. 251-266 DOY). • During the experimental period, T0 and T1 received a total of 374 and 143mm water, respectively, including rain water. • Leaf water potential (Ψleaf), turgor potential (Ψp leaf) and leaf conductance (gleaf) values for quince plants was noted. Miguel Hernández Polytechnic University of Elche, Orihuela (Spain) Griñán et al (2019)
- 58. Diurnal course of leaf water potential (Ψleaf), turgor potential (Ψp leaf) and leaf conductance (gleaf) values for quince plants in T0: CONTROL (closed circles) and T1 (open triangles) treatments at three different times during the stress period (DOY 224, 238, 251). Results: From the beginning of water stress to the time of maximum water stress, leaf turgor was maintained, possibly due to active osmotic adjustment (stress tolerance mechanism). This leaf turgor maintenance may have contributed to the high leaf conductance, and, therefore, good leaf productivity.
- 59. Predawn (Ψpd, A) and midday (Ψmd B) leaf water potential values Predawn (Ψppd, C) and midday (Ψpmd, D) leaf turgor potential values T0: CONTROL (closed circles) and T1 (open triangles) treatments
- 60. Midday stem water potential (Ψstem, A) & midday leaf conductance (glmd, B) values. T0: CONTROL (closed circles) and T1 (open triangles) treatments Conclusion: During the development of the severe water stress and recovery periods, quince trees showed stress tolerance mechanism by active osmotic adjustment. This leaf turgor maintenance would allow substantial leaf conductance levels and, therefore, good leaf productivity.
- 61. Rootstock-induced molecular responses associated with drought tolerance in sweet orange as revealed by RNA-Seq • Objective: To investigate the poorly understood molecular responses underlying the rootstock-induced drought tolerance in sweet orange. • RNA-Seq transcriptome analysis was carried out in leaves of sweet orange grafted on ‘Rangpur’ lime subjected to control and drought-stress treatments. • A total of 41,827 unique transcripts were identified, among which 1764 transcripts showed significant variation between the treatments, with 1081 genes induced and 683 repressed by drought-stress treatment. • RNA-Seq data were validated by quantitative real-time PCR (qPCR) analysis. • The transcripts were distributed in 44 different categories of cellular component, molecular function and biological process. • Several genes related to different pathways, were differentially regulated by drought stress. • Conclusion: This study provides a useful reference for further exploration of the functions of candidate genes and applications on the genetic improvement of citrus rootstocks. Gonçalves et al (2019) Universidade Estadual de Santa Cruz, Ilhéus, Brazil
- 62. Functional categorization of the differentially expressed genes into 44 different categories of cellular component, molecular function and biological process in leaves of drought-stressed sweet orange (Citrus sinensis) grafted on ‘Rangpur’ lime. Categorization was performed using BGI WEGO.
- 63. A schematic model of the rootstock-induced transcriptional response associated with drought tolerance in leaves of sweet orange. Dotted lines represent indirect connections and solid lines represent direct connections. Abbreviations: • PKs: protein kinases • PYR/PYL/RCAR: pyrabactin resistance 1/PYR1-like/regulatory components of ABA receptor • PP2C: protein phosphatase type 2C • SnRK2: SNF1- related protein kinase 2 • DREB2A: dehydration-responsive element binding protein 2A • HSP: heat shock protein • MPKs: mitogen-activated kinase • PRP: proline-rich protein; • WAKs: wall-associated kinases • TFs: transcription factors
- 64. Effect Of Drought Stress In In Vitro And Drought-related Gene Expression In Carrizo Citrange • Objective: To investigate the effect of in vitro drought stress on Carrizo citrange and expression levels of drought related genes. • Drought stress experiment was established with plants obtained from germinated seeds of Carrizo citrange. • For this purpose, MS mediums with 0, 1, 2, 4 and 6% PEG 8000 were prepared and the plants were transferred to these media. • 1 mg/L BA was added to promote micropropagation of plants into the MS medium. • The plants were sub cultured 3 times at each 4 weeks. • qRT-PCR Analysis: qRT-PCR analyzes were performed on leaves. • 15 different plant materials from 3 subcultures were used for the total of 5 concentrations. • RNA isolation was performed from the leaves. Expression levels of 3 different genes was noted. Simsek (2018) Cukurova University, Adana, Turkey
- 65. • Decrease in the multiplication coefficient in increasing PEG doses. • Three genes had accelerating expression levels with increasing PEG doses. • With these results, it has been revealed that drought-related genes are triggered.
- 66. 1 SC-PEG 0: Sub culture 1 PEG 0% 1 SC-PEG 6: Sub culture 1 PEG 6%
- 67. 1 SC-PEG 0: Sub culture 1 PEG 0% 1 SC-PEG 6: Sub culture 1 PEG 6%
- 68. 1 SC-PEG 0: Sub culture 1 PEG 0% 1 SC-PEG 6: Sub culture 1 PEG 6%
- 69. Drought Tolerance of Some Wine Grape Cultivars Under In Vitro Conditions • Objective: To determine the drought tolerance of 6 different economically important wine grape cultivars (‘Sultani Seedless’, ‘Çalkarası’, ‘Emir’, ‘Boğazkere’, ‘Öküzgözü’, ‘Narince’) of Turkey under in vitro conditions. • Drought stress was induced on in vitro-grown explants by 3 different PEG (8000) (poly ethylene glycol) doses (2, 4 and 6 %). • Plants were subjected to drought stress for 6 weeks. • Plant fresh weight, dry weight, shoot length, number of shoots, number of leaves, electrolyte leakage, relative water content, proline content were determined. • Being more distinctive at higher doses, PEG treatments yielded significant decreases in fresh weight, dry weight, shoot length, number of shoots and number of leaves & increase in proline content. • Conclusion: Öküzgözü and Sultani Seedless cultivars maximum G & Dvp under drought stress; seemed to be more resistant to drought. • But considering the other parameters (Electrolyte leakage, RWC and proline), it is better to indicate that there were not significant differences in drought tolerance of the cultivars. Neval and Rüstem (2019) Gaziosmanpaşa University, Tasliciftlik, Tokat-Turkey
- 70. The effects of different PEG doses applied to varieties on growth and development parameters
- 71. The effect of different PEG doses applied to the varieties on ion leakage, relative water content & proline accumulation
- 72. Genome-wide characterization and expression analysis of citrus NUCLEAR FACTOR-Y (NF-Y) transcription factors identified a novel NF-YA gene involved in drought-stress response and tolerance • Nuclear factor Y (NF-Y) is a ubiquitous transcription factor found in eukaryotes. It is composed of three distinct subunits called NF-YA, NF-YB and NF-YC. • NF-Ys have been identified as key regulators of multiple pathways in the control of development and tolerance to biotic and abiotic factors. • Objectives: 1. To identify and characterize the complete repertoire of genes coding for NF-Y in citrus. 2. To perform the functional characterization of one of its members, namely CsNFYA5, in transgenic tobacco plants. Pereira et al (2018) Universidade Estadual de Santa Cruz, IlheÂus, Bahia, Brazil
- 73. Experiment No. 1 • Two-year-old sweet orange (Citrus sinensis L. Osb. var. `Westin') plants grafted on Rangpur lime (C. limonia Osbeck) were used in the drought stress experiment. 1. Control (leaf predawn water potential between -0.2 to -0.4 MPa). 2. Drought stress (leaf predawn water potential of -1.5 MPa) treatments. • RNA extraction and quantitative real-time RT-PCR (qRT-PCR) analysis • All procedures of RNA extraction, cDNA synthesis and quantitative real-time RT-PCR (qRT-PCR) analysis were performed. • Identification and sequence analysis of citrus NF-Ys • Data obtained from qRT-PCR was compared with sweet orange genome data available at the Phytozome database. • Results: A total of 22 genes coding for NF-Y were identified in the genomes of sweet orange (Citrus sinensis) and Clementine mandarin (C. clementina), including 6 CsNF-YAs, 11 CsNF-YBs and 5 CsNF-YCs.
- 74. Experiment No. 2 • The plants of Nicotiana tabacum cv. Havana used in Agrobacterium-mediated genetic transformation experiments. • The stress tolerance of wild-type (WT) and transgenic tobacco plants (NF12, NF15, NF16, NF20 and NF22) was examined by in vitro drought stress tolerance assay under control and PEG treatments for 30 days. • CsNF-YA5 expression analysis in response to drought stress • Overexpression of CsNF-YA5 in transgenic tobacco plants contributed to the reduction of H2O2 production under dehydration conditions and increased plant growth and photosynthetic rate under normal conditions and drought stress. • These biochemical and physiological responses to drought stress promoted by CsNF-YA5 may confer a productivity advantage in environments with frequent short-term soil water deficit.
- 75. In vitro drought stress tolerance assay of CsNFYA5-overexpressing transgenic lines. Representative phenotypes of control (WT and PC) and CsNFYA5 transgenic (NF12, NF15, NF16, NF20 and NF22) lines grown under control and PEG treatments for 30 days. Fig. A Seedling biomass & Fig. B root length.
- 76. Representative phenotypes of control (WT and PC) and CsNFYA5 transgenic (NF12, NF15, NF16, NF20 and NF22) lines grown under control and PEG treatments for 30 days. Fig. C Seedling biomass & Fig. D root length.
- 77. Physiological and morphological adaptations of the fruit tree Ziziphus rotundifolia in response to progressive drought stress • Objective: To study physiological basis of drought resistance in Ziziphus rotundifolia Lamk. • Three irrigation regimes were imposed over a 34-day period: • Irrigation treatment (control). • Gradual drought stress treatment (50% of water supplied in the irrigation treatment i.e. control). • Rapid drought stress treatment (no irrigation). • Changes in gas exchange, water relations and solute concentrations of leaves, stems and roots were determined. • Results: The differential rate of stress development in the two drought treatments did not result in markedly different physiological responses, but merely affected the time at which they were expressed. • Conclusion: Z. rotundifolia showed a range of responses to different drought intensities indicating a high degree of plasticity in response to water deficits. Arndt et al (2001) Institute of Ecology and Conservation Biology, Vienna, Austria
- 78. Changes in soil water content in the irrigated, gradual stress and rapid stress treatments. • In the gradual stress treatment, pot soil water content declined steadily, reaching 26% of the initial pre-stress value after 34 days. • In the rapid stress treatment, 76% of the initial water was lost within 16 days. Soil water content continued to decline throughout the study, reaching 16% of the initial water content on Day 34.
- 79. (a) Predawn leaf water potential (b) Predawn osmotic potential of leaf tissue sap (b) Stomatal conductance (gleaf) (c) Intrinsic WUE The initial response to decreasing soil water content was reduced stomatal conductance, effectively maintaining predawn leaf water potential (Ψleaf), controlling water loss and increasing intrinsic water-use efficiency, while optimizing carbon gain during drought.
- 80. Concentrations of starch, proline & sugars in irrigated and drought-stressed Ziziphus rotundifolia trees at three harvests. • As drought developed, osmotic potential at full turgor decreased and total solute concentrations increased in leaves, indicating osmotic adjustment. • Decreases in leaf starch concentrations and concomitant increases in hexose sugars and sucrose suggested a shift in carbon partitioning in favor of soluble carbohydrates. • In severely drought-stressed leaves, increase in proline concentration, suggests an osmoprotective role for proline.
- 81. Physiological, biochemical and molecular responses in four Prunus rootstocks submitted to drought stress • Objective: To study physiological, biochemical and molecular drought responses of four Prunus rootstocks (GF 677, Cadaman, ROOTPAC 20 and ROOTPAC® R) budded with ‘Catherina’ peach cultivar. • Two water treatments: well irrigated and water stress. • Control plants were watered daily and water status was maintained at full field capacity. • Water-stressed plants were also irrigated daily, but adding ~80% of the water evapotranspired the previous day and subjected to progressive water stress during 26 day. • Photosynthetic and gas exchange parameters were determined. Root and leaf soluble sugars and proline content were also measured. • At the end of the experiment, stressed plants showed lower net photosynthesis rate, stomatal conductance and transpiration rate, and higher intrinsic leaf WUE (AN/gs). • Soluble sugars and proline concentration changes were observed, in both root and leaf tissues, especially in an advanced state of stress. Zaragoza, Spain Jiménez et al (2013)
- 82. Daily soil volumetric water content of control and drought-stressed pots containing Prunus rootstocks (Cadaman, CD; GF 677, GF; ROOTPAC 20, R20; and ROOTPAC® R, RR) budded with var. ‘Catherina’.
- 83. Daily scion apical growth of control and drought-stressed four Prunus rootstocks budded with var. ‘Catherina’.
- 84. Midday stem water potential (Ψs) in scion leaves of control and drought-stressed Prunus rootstocks Cadaman (a), GF 677 (b), ROOTPAC 20 (c) and ROOTPAC® R (d) budded with var. ‘Catherina’.
- 85. Scion leaf soluble sugars and proline (mg g−1 DW) concentration (n = 6) in control and drought-stressed Prunus rootstocks after 16 & 26 days of treatment. The accumulation of proline in roots and leaves with drought stress was related to the decrease in osmotic potential and increase in WUE Whereas the accumulation of sorbitol in leaves, raffinose in roots and proline in both tissues was related only to the increase in the WUE. 16 days 26 days
- 86. Expression profiles: • Raffinose synthase (SIP1) • Δ-1-pyrroline-carboxylate synthase (P5SC) • sorbitol 6-phosphate dehydrogenase (S6PDH) gene In leaves & roots Cadaman (CD) and GF 677 (GF) rootstocks’ and submitted to control and drought treatments for 16 days. • Higher expression of P5SC gene in roots was also consistent with proline accumulation in the tolerant genotype GF 677. • Accumulation of sorbitol, raffinose and proline in different tissues and/or the increase in P5SC expression could be used as markers of drought tolerance in peach cultivars grafted on Prunus rootstocks.
- 87. Conclusion • The biochemical responses to drought, mainly accumulation of sorbitol, raffinose and proline, were consistently related to the physiological responses to water stress that confer tolerance. • Initial molecular responses were related to the biochemical responses observed. • Accumulation of leaf sorbitol, root raffinose and root and leaf proline could be implemented as a drought tolerance markers for early selection of Prunus rootstocks for peach trees under controlled conditions. • The differential expression of PSC5 in roots could also be used as drought tolerance marker. • The almond-based rootstocks GF 677 and ROOTPAC® R showed better performance to drought stress with both physiological and biochemical responses. • The different rootstock performance could be related to their different genetic background and vigour. • This study would be the basis for future analysis at the whole-molecular level to illuminate the tolerance mechanisms to drought in Prunus rootstocks.
- 88. Tolerance of citrus plants to the combination of high temperature and drought is associated to the increase in transpiration modulated by a reduction in abscisic acid levels • Objective: To study physiological parameters and the hormonal regulation of Carrizo citrange and Cleopatra mandarin, two citrus genotypes, in response to the combined action of high temperatures and water deprivation. • Four experimental groups of each genotype: • Well-watered plants at 25 °C (CT: control) • Well-watered plants at 40 °C (HS: heat stress) • Drought affected plants at 25 °C (WS: water stress) • Drought affected plants at 40 °C (WS +HS: Combo of both). • Leaf tissue was sampled at 24 h after subjecting plants to both stresses. • Results: Carrizo citrange is more tolerant to the stress combination than Cleopatra mandarin. Zandalinas et al (2016) Universitat Jaume I, Castelló de la Plana, Spain
- 89. Intact sprouts (%) of Carrizo and Cleopatra seedlings subjected to drought and heat stress (40 °C) in combination for 10 days.
- 90. Relative water content (RWC) (a) and proline concentration (b) in Carrizo and Cleopatra plants subjected to drought (WS), heat (HS) and their combination (WS + HS).
- 91. SA levels in citrus plants subjected to different stress treatments • SA accumulated in response to individual stresses but to a higher extent in samples subjected to the combination of heat and drought (showing an additive response). • SA accumulation correlated with the up-regulation of pathogenesis-related gene 2 (CsPR2), as a downstream response.
- 92. • ABA accumulation was higher in water-stressed plants followed by that observed in plants under stress combination. • ABA signaling in these plants was confirmed by the expression of responsive to ABA-related gene 18 (CsRAB18). ABA levels in citrus plants subjected to different stress treatments
- 93. SUITABILITY OF FRUIT CROPS UNDER HIGH TEMPERATURE STRESS CONDITIONS
- 94. High temperatures change the perspective: integrating hormonal responses in citrus plants under co-occurring abiotic stress conditions • Objective: To study physiological, hormonal and molecular responses of the citrus rootstock, Carrizo citrange (Poncirus trifoliata L. Raf. × Citrus sinensis L. Osb.) subjected to wounding or high salinity occurring individually or in combination with heat stress. • Six experimental groups were established: 1. Intact plants growing at 25 ᵒC (control, CT) 2. Intact plants growing at 40 ᵒC (heat stress, HS) 3. Wounded plants growing at 25 ᵒC (wounded, Wo Wounding was performed by drilling 50% of the leaves with a paper puncher ) 4. Wounded plants growing at 40 ᵒC (wounded + heat shock, Wo+HS), 5. Salinized plants growing at 25 ᵒC (NaCl 60mM NaCl) 6. Salinized plants growing at 40 ᵒC (NaCl+HS). • All conditions were applied at the same time for 15 days. • Conclusion: Co-occurring abiotic stress conditions can modify (either enhance or reduce) the hormonal response to modulate specific responses. Balfagón et al (2018) Universitat Jaume I, Castelló de la Plana, Spain
- 95. Leaf RWC of Carrizo plants subjected to wounding (Wo), salt stress (NaCl), heat stress (HS), combination of wounding and heat stress (Wo+HS) and combination of salt and heat stress (NaCl+HS) for 15 days.
- 96. Content of ABA, JA & SA of Carrizo plants subjected to wounding (Wo), salt stress (NaCl), heat stress (HS), combination of wounding and heat stress (Wo+HS) and combination of salt and heat stress (NaCl+HS) for 15 days.
- 97. Relative expression of genes involved in ABA biosynthesis CsNCED1 CsAAO3 Signaling CsAREB1 CsAREB2 CsRD22 CsRD29
- 98. Relative expression of genes involved in JA biosynthesis CsLOX CsAOS CsJAR1 Signaling CsVSP2
- 99. Relative expression of genes involved in SA biosynthesis CsICS1 CsPAL Signaling CsPR2
- 100. Involvement of ascorbate peroxidase and heat shock proteins on citrus tolerance to combined conditions of drought and high temperatures • Objective: To study stress tolerance mechanism in two citrus rootstocks Carrizo citrange (Poncirus trifoliata L. Raf. X Citrus sinensis L. Osb.) and Cleopatra mandarin (Citrus reshni Hort. ex Tan.) with contrasting tolerance to drought and heat stress. • Four experimental groups of each genotype: • Well-watered plants at 25 °C (CT: control) • Well-watered plants at 40 °C (HS: heat stress) • Drought affected plants at 25 °C (WS: water stress) • Drought affected plants at 40 °C (WS +HS: Combo of both). • Leaf tissue was sampled 24 h after the stress imposition. • Results: Carrizo citrange is more tolerant to the stress combination than Cleopatra mandarin. Zandalinas et al (2016) Universitat Jaume I, Castelló de la Plana, Spain
- 101. Stress-induced leaf damage Carrizo citrange (A, C and E) Cleopatra mandarin (B, D and F) A and B: Intact leaves C and D: Chlorotic leaves E and F: Partially necrotic leaves.
- 102. Leaf damage & stomatal conductance (gs) in Carrizo and Cleopatra plants subjected to different stress Less leaf damage in Carrizo than in Cleopatra under combined conditions of drought and heat stress. Stomatal conductance increased in heat stress due to HSP.
- 103. APX activity and APX2 gene expression in Carrizo and Cleopatra plants. • APX activity was less under water stress conditions, moderate under WS+HS conditions, but highest under HS conditons. • And it was comparatively higher in Carrizo than in Cleopatra under stress combination. • Higher APX2 gene expression in Carrizo, contributes to the higher tolerance of Carrizo plants to the combination of stresses. • Carrizo is better rootstock than Cleopatra (traditionally used in areas with scare water supplies).
- 104. APX2, HSP101 and HSP17.6 levels in Carrizo and Cleopatra plants subjected to stress. Top: Quantification graphs Bottom: Representative protein blots and loading controls
- 105. Conclusion • The pattern of accumulation of three proteins (APX, HSP101 and HSP17.6) involved in abiotic stress tolerance shows that they do not accumulate under water stress conditions individually applied. • Contents of APX and HSP101 are higher in Carrizo than in Cleopatra under stress combination • Whereas HSP17.6 has a similar behavior in both types of plants. • This, together with a better stomatal control and a higher APX activity of Carrizo, contributes to the higher tolerance of Carrizo plants to the combination of stresses. • Carrizo is better rootstock than Cleopatra (traditionally used in areas with scare water supplies) under the predictable future climatic conditions with frequent periods of drought combined with high temperatures. • This work also provides the basis for testing the tolerance of different citrus varieties grafted on these rootstocks and growing under different field conditions.
- 106. Effect of high temperature stress on the reproductive growth of strawberry cvs. ‘Nyoho’ and ‘Toyonoka’ • Objective: To examined the effect of two day/night temperature regimes on fruit set and fruit growth in two cultivars, ‘Nyoho’ and ‘Toyonoka’. • 10 plants of each cultivar were two growth chambers kept at either 23/18 C (control) or 30/25 C (high) day/night temperatures. • The percentage of fruit set in ‘Nyoho’ was not significantly different between the two temperature treatments, while that in ‘Toyonoka’ was much lower at 30/25 C than at 23/18 C. • Days to ripening was shorter at 30/25 C than at 23/18 C, and no cultivar differences were observed. • Fresh weight & diameter of fruits reduced at 30/25 8C in both cultivars • Percentage of fruit set at 30/25 C was also significantly lower in ‘Toyonoka’ than in ‘Nyoho’. ‘Nyoho showed resistance to high temperature. • These results indicate that high temperature stress negatively affects the reproductive process in strawberry and that plant response to high temperature stress is cultivar specific too. Ledesma et al (2008)University of Tokyo, Japan
- 107. Fully ripe fruits obtained from ‘Nyoho’ (A) and ‘Toyonoka’ (B) strawberry plants grown at 30/25 C. In ‘Toyonoka’, low fruit set caused malformations and reductions in the sizes of fruits. Mean number of inflorescences Fresh weight of fruits
- 108. A Study on the High Temperature Treatment to Three Grapevine Varieties Grown Under In vitro Conditions • Objective: To determine temperature grades and exposure times to stresses in plants of vine varieties grown in vitro conditions. • Three varieties: Çalkarası, Öküzgözü and Narince). • The durations of the stress conditions were determined with the preliminary study carried out with the Çalkarası variety. • Then, the three varieties used in the study were exposed to stress for the same durations. • Plants with appropriate sizes (5-6 leaves) were placed in a plant growth cabinet at 25ºC with a photoperiod of 16 h. • Heat stress at temperatures of 35ºC and 40ºC were applied. Altıncı et al (2018) Gaziosmanpaşa University Tokat, TURKEY
- 109. In vitro plants of Çalkarası exposed to 35 ºC stress at different times (24, 36, 48 hours) In vitro plants of Çalkarası exposed to 40 ºC stress at different times (9, 12, 15 hours) Exposure time 36 hours at 35˚C and 12 hours at 40˚C did not lose the vitality of the plants, but plants seriously entered the stress.
- 110. 35 ºC stress application for 36 hours in Çalkarası, Öküzgözü and Narince 40 ºC stress application for 12 hours in Çalkarası, Öküzgözü and Narince More damage was found at 40 ºC than 40 ºC stress application. Among three varieties, least damage was found in Öküzgözü.
- 111. Climate warming enhances flower formation, earliness of blooming and fruit size in plum (Prunus domestica L.) in the cool Nordic environment • Objective: To analyse data from a 31-year series of observations (1985–2016) on spring phenology and flowering and fruiting performance of three plum cultivars in an experimental orchard at Ås in southeast Norway (59° 40′N; 10° 50′E). • As temperature increased in March and April, the date of full bloom (FB) reduced than earlier low temp. On average for all cultivars, blooming was advanced by 10 days over the study period. • August and September temperature, which also increased significantly during the study period, was closely positively correlated with the amount of flowering in the subsequent spring and also interacted with early spring temperature in advancing blooming time. • Conclusion: Increasing March and April temperatures during the last 30 years has advanced blooming and spring phenology in plum and the resulting extension of the growing season has led to increasing fruit size at harvest. • Climate warming appears to have been positive for plum production in the cool Nordic environment. Woznickia et al (2019) Norwegian University of Life Sciences, Ås, Norway
- 112. Trends in March, April and May temperatures at Norway, and the corresponding trends in FB from 1 January for the three studied plum cultivars over the years 1985–2016.
- 113. Scatterplot and trend line for the mean fruit size at harvest for each year across the 1985–2016 period for the three studied plum cultivars.
- 114. Late blooming was negatively correlated with fruit size at harvest for all cultivars. Early blooming had significant positive correlation between fruit size at harvest and the number of days from FB to harvest for all cultivars. Since the period between blooming and fruit maturity (harvest) increased during the observed 31- year period, fruit size at harvest also increased across the years for all cultivars.
- 115. SUITABILITY OF DIFFERENT FRUIT CROPS UNDER LOW TEMPERTURE STRESS CONDITIONS
- 116. Evaluation of susceptibility of pear and plum cultivars to winter frost • Objective: To investigate winter frost damage to pear and plum buds in Mashhad, after a period of unusually warm weather for a few days in March 2013 followed by a sudden drop in temperature to -12°C. • Two separate experiments included. • Twelve cultivars of pear: William’s, Bell de june, Ida, Spadona, Koshia, Domkaj, Dare Gazi, Mohamad Ali, Boheme, Asian pear, Shekari and Ghodumi. • Eight plum cultivars (European: Prunus domestica L. Stanley, Early Santa Rosa, Late Santa Rosa, Shablon, Black Diamond; Japanese: P. salicina L. Shiro, Shams, Computi) • Frost damages of vegetative and reproductive buds and spur-part below the buds of pear and plum cultivars were investigated based on visual observations. • After 24 h keeping at laboratory temperature, a hundred vegetative and reproductive bud samples of each cultivar were visually evaluated for brown discoloration. Khorshidi et al (2014) Ferdowsi University Of Mashhad, Mashhad, Iran
- 117. • Minimum & maximum temperatures from February 19th to March 20th, 2013. • For most of the time the lowest temperature was not below zero in February and March until March 7th, when it reached -5.2°C, and the lowest one was -12.2°C on March 9th.
- 118. Vegetative bud of pear var. ‘Ghodumi’ suffered the most damage. Reproductive bud of ‘Ghodumi’ suffered the least damage. The EC and proline content of ‘Dare Gazi’ were the highest despite it did not show high resistance. (Reason: Proline increase doesn’t increase cold resistance).
- 119. Percentage of frost damage of pistil and male organs of some plum cultivars • In all plum cultivars the pistil suffered more damage than the male part. • Percentage of cold injury in flower bud was higher than other tissues, and the cold hardiness of pistil was weaker than stamen.
- 120. • All parts of ‘Late Santa Rosa’ were the most susceptible in plum cultivars. • ‘Computi’ had the most resistant reproductive bud and no damage was observed in the other parts. It had the lowest proline content. • Conclusion: Proline index is not appropriate for evaluating frost damage, but there was a high correlation was between EC and frost damage.
- 121. Metabolic Responses to Low Temperature of Three Peach Fruit Cultivars Differently Sensitive to Cold Storage • Refrigerated storage is widely applied in order to maintain peach quality but it can also induce chilling injuries (CIs) such as flesh browning and bleeding. • Objective: To study changes in both metabolome and volatilome induced by cold storage in the mesocarp of three peach cultivars (‘Red Haven’, RH, ‘Regina di Londa’, RL, and ‘Flaminia’, FL). • Fruits of ‘Red Haven’ (RH, yellow fleshed), ‘Flaminia’ (FL, yellow-fleshed, C35 days from RH harvest), ‘Regina di Londa’ (RL, white fleshed, C55 days from RH harvest) were harvested at flesh firmness values of 37, 42, and 56 N for RH, FL, and RL, respectively, • Immediately after transfer to the lab, fruit were incubated in cold chambers at two low temperature conditions (0.5 and 5.5 C) and reference (control) at 20 C. • Peaches were sampled at harvest (T0) and after 1, 2, 3, and 4 weeks of cold storage, whereas for the control at 20C the 4th week sampling was not performed due to over-ripening and decay. • Considering that CI symptoms appear after fruit removal from cold storage, fruit were additionally kept for 3 days at room temperature to evaluate the post-storage behavior and the shelf-life (SL). Brizzolara et al (2018) Istituto di Scienze della Vita, Scuola Superiore Sant’Anna, Pisa, Italy,
- 122. Flesh firmness (A) & TSS (B) measured at end of 3 week in control & at 4 week in cold treatment. Further cold storage fruits kept for three additional days under shelf-life conditions (20 C, Shelf Life; dotted lines) and on peaches sampled immediately after cold storage (no SL; solid lines). Black diamonds: Control fruit 20 C Blue circles: 0.5 C Red squares: 5.5 C RL showed highest firmness at 0.5 C. No SL (solid lines) had more firmness than 20 C SL (dotted lines). FL & RH showed highest TSS at 5.5 C. But RL showed highest TSS at 0.5 C. 20 C SL (dotted lines) showed highest TSS than No SL (solid lines).. Compared to RL and FL, RH showed a marked less reduction in firmness & TSS
- 123. Incidence of CIs (flesh bleeding, A & flesh browning, B) measured at end of 3 week in control & at 4 week in cold treatment. Further cold storage fruits kept for three additional days under shelf- life conditions (20 C, Shelf Life; dotted lines) and on peaches sampled immediately after cold storage (no SL; solid lines). Black diamonds: Control fruit 20 C Blue circles: 0.5 C Red squares: 5.5 C Flesh bleeding revealed high incidence following 0.5C storage. Flesh browning appeared to be more related to 5.5C storage. Compared to RL and FL, RH showed a marked lower incidence of CIs.
- 124. Conclusion • RH peaches indeed differ from RL and FL in particular when considering data from samples collected at the end of the cold storage. • In all three cultivars, glucose-6 P, xylose, sorbitol, epicatechin, catechin, and putrescine markedly increased during cold storage, while glucuronic, mucic and shikimic acids decreased. • Among volatile organic compounds (VOCs), aldehydes and alcohols generally accumulated more under low temperature conditions while esters and lactones evolved during subsequent shelf life. • The lower levels of flesh browning and bleeding displayed by RH peaches were related to compounds with antioxidant activity, or acting as osmotic protectants and membrane stabilizer. • Indeed, RH showed higher levels of amino acids and urea, together with a marked increase in putrescine, sorbitol, maltitol, myoinositol and sucrose detected during storage and SL.
- 125. Cold Hardiness and Options for the Freeze Protection of Southern Highbush Blueberry • Southern highbush blueberries are a low chill species of blueberry that are commercially grown in sub-tropical climates. • Due to the nature of SHB, the flowering and fruit set occur in mid-winter to early spring and are susceptible to freeze damage. • Of the various methods used in frost protection, applying overhead irrigation water is the most promising. • During a freeze event, an application of 6.3 mm/ha of water per hour is required to protect blueberries from 2.8 C (27 F) temperature with winds from 0 to 16 km/h (0 to 10 miles/h). • This is 25.4 kL/h/ha of water. • This paper will review methods of freeze/frost protection, importance of weather patterns, and critical temperatures based on phenology of flowering to fruit. • Conclusion: Understanding weather patterns, flower phenology, and the capacity of the freeze protection system are the basis for mitigating freeze damage to southern highbush blueberry. Smith(2019) University of Georgia, Tifton Campus, Tifton, USA
- 126. Tight bud (stage 1) Bud swell (stage 2) Bud break (stage 3) Tight bud and bud swell can tolerate cold temperatures of −12 to −9 °C (10–15 °F). Bud break is more sensitive to cold, where at −7 °C (20 °F), damage can occur.
- 127. Tight cluster (stage 4) Early pink bud (stage 5) Tight cluster can tolerates cold to −7 to −5 °C (20–23 °F). Early pink bud can tolerate cold to −4.4 to −2.8 °C (23–27 °F).
- 128. Pink buds (stage 6) Late pink bud (stage 7) Pink buds are cold tolerant to −5 to −4 °C (23–25 °F). Late pink bud are cold tolerant to −4.4 to −2.8 °C (24–27 °F).
- 129. Full bloom (stage 8) Petal fall (stage 9) Full bloom are cold tolerant to −2.2 °C (28 °F). Petal fall are cold tolerant 0 °C (32 °F). Cold tolerance decreased as growth proceed from tight bud to petal fall. Once the petals fall (stage 9), freeze damage will occur at 0 °C. Even small green fruit is sensitive to 0 °C.
- 130. Comparative metabolic profiling of Vitis amurensis and Vitis vinifera during cold acclimation • Objective: To investigate accumulation of metabolites in V. amurensis during cold acclimation (CA) which is wild Vitis plant that can withstand extreme cold temperatures • Plantlets of V. amurensis and V. vinifera cv. Muscat of Hamburg were treated at 4 °C for 24 and 72 h, and changes of metabolites in leaves were detected. • Most of the identified metabolites, including carbohydrates, amino acids, and organic acids, accumulated in the two types of grape after CA. • Sugars: Galactinol, raffinose, fructose, mannose, glycine, and ascorbate were continuously induced by cold in V. amurensis, but not in Muscat of Hamburg. • Twelve metabolites increased in V. amurensis during CA. • More galactinol, ascorbate, 2-oxoglutarate, and putrescine, accumulated in V. amurensis, but not in Muscat of Hamburg, during CA. • All these metabolites are responsible for the excellent cold tolerance in V. amurensis. • The expression levels of the genes encoding β-amylase (BAMY), galactinol synthase (GolS), and raffinose synthase (RafS) were evaluated by qRT-PCR. • In conclusion, these results show the inherent differences in metabolites between V. amurensis and V. vinifera under CA. Chai et al (2019)Chinese Academy of Sciences, Wuhan, P.R. China
- 131. Metabolites that specifically accumulated in V. amurensis and V. vinifera cv. Muscat Hamburg under the cold stress condition.
- 132. qRT-PCR results for BAMY gene family members in V. amurensisand V. vinifera cv. Muscat Hamburg. The expression BAMY (VIT_02s0012 g00170) was primarily responsible for the accumulation of maltose.
- 133. qRT-PCR results for GolS gene family members in V. amurensisand V. vinifera cv. Muscat Hamburg. The expression GoIS (VIT_14s0060g00760) was primarily responsible for the accumulation of galactinol.
- 134. qRT-PCR results for RafS gene family members in V. amurensisand V. vinifera cv. Muscat Hamburg. The expression RafS (VIT_05s0077 g00840) was primarily responsible for the accumulation of raffinose.
- 135. ADAPTATION OF DIFFERENT FRUIT CROPS TO BIOTIC STRESS
- 136. Resistance gene analogues in mango against mango malformation • Plant resistant gene analouges (RGA): Resistant gene having conserved domains that plays specific roles in pathogens resistance. These are receptors like proteins. • H-8-11 (A×LS) and Bhadauran showed a more than 50% sequence similarity to other resistance gene analogues. • So far malformation has not been observed in these two cultivars. • These sequences can be used for further studies in the mango malformation resistance breeding programs. • The cloning of resistance gene sequences provides the possibility of using the transgenic approach to increase disease resistance of plants Ebrahim et al (2013)Indian Agricultural Research Institute, New Delhi, India
- 137. Evaluation of Different Papaya Cultivars for Yield, Quality and Papaya Ring Spot Disease under Pune Conditions • Objective: To access the suitability of different papaya cultivars to Pune conditions. • Nine papaya cultivars (Washington, CO 2, CO 6, Pusa Dwarf, Pusa Giant, Pusa Nanha, Pusa Delicious, Pusa Majesty and Red Lady) were evaluated for growth, yield, quality and PRSV incidence. • Results: Cv. Pusa Giant recorded significantly maximum fruit yield/plant (66.1kg) • Cv. Red Lady showed early flowering (70.4 Days after planting; DAP) which was at par with cv. CO 2 (73.5 DAP), Pusa Nanha (73.7 DAP) and Pusa Dwarf (73.6 DAP). • Regarding quality parameters viz. pulp thickness and TSS were maximum (3.1cm and 11.71 o Brix, respectively) in cv. Red Lady Chalak et al (2016) College of Agriculture, Pune-411005, India.
- 138. Evaluation of Papaya Cultivars
- 139. Reaction to papaya ring spot disease (PRS) Among these cultivars, the magnitude of infestation was lowest DPI (66.0 as Percentage Disease Index) in Pusa Giant at 10 months after planting. The maximum infestation was observed in widely adopted cv Red Lady (98.7) at the same age. From present investigation cv Pusa Giant was found as a good yielder and also tolerant to ring spot disease among these papaya cultivars.
- 140. Resistance of transgenic papaya plants to Papaya ringspot virus • Objective: To generate transgenic papayas resistant against Papaya ringspot virus, • The full-length coat protein (CP) gene was amplified from the RNA of PRSV isolated from Ratchaburi province, Thailand by RT-PCR. • A binary vector containing the coat protein gene under the control of a 35S promoter, was constructed and transformed into somatic embryos of papaya cultivar Khak Dum by microprojectile bombardment. • Eight transgenic lines were identified. • Integration of the transferred genes into papaya calli was verified by PCR amplification of the coat protein gene, • Although the coat protein gene was detected in all transgenic lines, only line G2 was found to be highly resistant to virus. Because coat protein gene was expressed by this line. Kertbundit et al (2007) Mahidol University, Bangkok, Thailand
- 141. ‘HoneySweet’ (C5), the First Genetically Engineered Plum pox virus–resistant Plum (Prunus domestica L.) Cultivar • ‘HoneySweet’ originated as a seedling from the open pollination of ‘Bluebyrd’ plum (Scorza and Fogle, 1999). The pollen parent of ‘HoneySweet’ is unknown. • ‘HoneySweet’ was originally selected in vitro shoots regenerated from a hypocotyl slice of Bluebyrd plum seedling. • Transformed with Agrobacterium tumefaciens carrying the plasmid PPV-CP- 33 (some prunus sp) Transgenic shoots, coded as C5 along with other transgenic shoots, was rooted in vitro and transferred to a greenhouse. . • Asexually propagated by bud grafting on different rootstock of plum and peach. • Aphid inoculations with the different strains of Plum pox virus (PPV Testing (PCR, ELISA). • Field evaluation of transgenic plants for almost twelve years in aphid vectored conditions in different countries later known as Honey Sweet (E). U.S Kearneysville Scorza et al (2016)
- 142. Three-year old ‘HoneySweet’ tree in Bistrita, Romania (left), with a weighed sample of ‘HoneySweet’ fruit (right).
- 143. Transgenic Virus Resistant Papaya: From Hope to Reality for Controlling Papaya Ringspot Virus in Hawaii • Papaya ringspot is the most important disease limiting the production of papaya worldwide. • Symptoms of PRSV on Papaya tree (a) and fruit (b) • Symptoms: Severe leaf mosaic, reduction of leaf canopy as the disease progresses, stunting of the plant, water-soaked oily streaks on the petiole and upper part of the trunk, and ringed spots on the surface of the fruits. Gonsalves et al (2004)U.S. Pacific Basin Agricultural Research Center, Hawaii
- 144. Effect of rapid virus spread through commercial fields. A healthy commercial papaya field in Puna in 1940. Viral epidemic in 1992 & all plants got infected. PRSV was first reported in Hawaii in the mid-1940s, it did not become a major problem to the Hawaiian papaya industry until the 1950s (Gonsalves, 2004).
- 145. How transgenic papaya formed ? • Isolation of coat protein (CP) gene of the mild mutant PRSV HA 5-1. • Insertion of coat protein gene in vector pGA482GG • Resulting plasmid DNA delivered via high velocity micro-projectiles gun into the papaya cells (Sunset and Kapoho) • As a result, sufficient number of transgenic papaya plants were obtained. • R0 micropropagated plants, designated as 55-1, when tested showed excellent resistance against PRSV. • Furthur, Line 55-1 was crossed with non-transgenic Sunset under green house condition with 50 % progenies being transgenic to produce R1 plants. • The homozygous line 55-1 named as ‘SunUp’, was red fleshed transgenic cultivar, the dominant cultivar growing in Hawaii. • The cross of transgenic SunUp and non-transgenic Kapoho was made to obtain the yellow fleshed hybrid ‘Rainbow’.
- 146. • Kapoho field trial started in 1995, showing a solid block of PRSV-resistant Rainbow growing well while the surrounding susceptible non-transgenic Sunrise is severely infected with PRSV. • Picture taken 19 months after start of the field trial.
- 147. Transgenic papaya line 55-1 showing resistance to PRSV HA compared to infected non-transgenic papaya.
- 148. Conclusion & Future prospects • The understanding of the underlying mechanisms of response and the existing interactions among abiotic & biotic stressors will provide valuable information for crop improvement. • Breeding for resistance to exposure to combined abiotic and biotic stress is needed to be done. It can be done either by incorporation of genetic components regulating the response or by gene pyramiding to both stresses. • Gene cloning: Resistance genes to particular stress should be cloned. The cloning of resistance gene sequences provides the possibility of using the transgenic approach to increase stress resistance in plants without the transfer of undesirable characters to the offspring.
- 149. THANK YOU
Editor's Notes
- Fig. 1 The complexity of the plant response to abiotic stress. Primary stresses, such as drought, salinity, cold, heat and chemical pollution are often interconnected, and cause cellular damage and secondary stresses, such as osmotic and oxidative stress. The initial stress signals (e.g. osmotic and ionic effects, or temperature, membrane fluidity changes) trigger the downstream signaling process and transcription controls which activate stress-responsive mechanisms to re-establish homeostasis and protect and repair damaged proteins and membranes. Inadequate response at one or several steps in the signaling and gene activation may ultimately result in irreversible changes of cellular homeostasis and in the destruction of functional and structural proteins and membranes, leading to cell death
- P is used to maintain the integrity of tissue structures and proper functioning of cell membranes under saline stress. Potassium concentration into plant material during salt solution treatments is recognized to have a beneficial role in osmotic adjustment, and its accumulation in parallel with high Na uptake is considered to be a parameter for plant tolerance to NaCl
- The decrease in TCC is a typical symptom of oxidative stress and could be due to pigment degradation, chlorophyll synthesis deficiency and changes in thylakoid membrane structure [52-53]. The degradation of the photosynthetic apparatus in the leaves exposed to extended water deficit aimed to mobilize resources for the production of new acclimated leaves Important role of soluble sugars in osmotic adjustment and cell protection by inducing direct detoxification processes of reactive oxygen species or by stimulating indirectly the antioxidative defense system. Cultivars with low photosynthetic activity tend to synthesize more soluble sugars in order to maintain the translocation of assimilates such as sugars to the growing parts