Recommended

Recommended
More Related Content
Viewers also liked
Viewers also liked (11)
Similar to Epidural Spinal Cord Stimulation for Modulating Small Sensory Nerve Fibers to the Pancreas: A Pilot Study
Similar to Epidural Spinal Cord Stimulation for Modulating Small Sensory Nerve Fibers to the Pancreas: A Pilot Study (20)
Recently uploaded
Recently uploaded (20)
Epidural Spinal Cord Stimulation for Modulating Small Sensory Nerve Fibers to the Pancreas: A Pilot Study
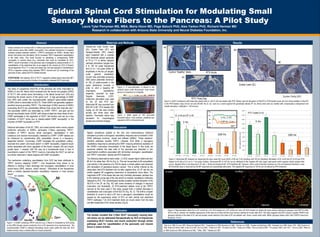
- 1. Epidural Spinal Cord Stimulation for Modulating Small Sensory Nerve Fibers to the Pancreas: A Pilot Study Laura Tyler Perryman MS, MBA, Maria Hizon MD, Page Baluch PhD, Alex Yarkov PhD, Richard Herman MD Research in collaboration with Arizona State University and Neural Diabetes Foundation, Inc. Abstract Materials and Methods Discussion References Obese animals and humans with or without glucose/insulin dysfunction often exhibit small sensory nerve fiber (SSNF) neuropathy. One potential mechanism is impaired transient receptor potential vanilloid 1 (TRPV1) expression on SSNFs. Studies have shown that spinal cord stimulation (SCS) increases TRPV1-dependent vasodilatation in the lower limbs. This study focused on identifying a corresponding SSNF neuropathy in visceral tissue (e.g., pancreas) that could be modulated by SCS. TRPV1 neuron innervation of the pancreas was investigated by measurements of: (1) vasodilatation of the abdominal skin (a surrogate for the viscera) by SCS of thoracic neural segments T9 to T11 in lean and obese rats, and rats exposed to resiniferatoxin (RTX) a capsaicin analog which depletes TRPV1 neurons and (2) morphology of the pancreas of lean, obese and RTX treated animals. HYPOTHESIS: 5Hz biphasic SCS of T9-T11 segments creates higher blood flow (BF) to the pancreas preserving the function of neuropathic stressed ISLET beta cells. Zucker Fatty (ZF) Zucker Lean (ZL) Results Twenty-one male Zucker Lean (ZL), Zucker Fatty (ZF), and Sprague-Dawley (SD) rodents were implanted with a unipolar SCS electrode placed epidurally at T9 to T11 to deliver biphasic cathodal stimulation (ranges from 5 to 100 Hz pulse frequencies and 0.2 to 1 ms pulse widths for amplitudes in the low A ranges) under general anesthesia. Current was controlled producing EMG action potentials (minimum of 100 V peak-to-peak) in the upper abdominal muscles in order to elicit a baseline BF (neurogenic vasodilatory) response, recorded by laser Doppler flowmetry (LDF). Antidromic stimulation was used on the ZF and RTX rats. Abdominal BF was recorded from 80% MT to MT. RTX-treated (250 mg/kg, sc) SD rats were studied acutely and three days after injection. Pancreatic tissue was harvested for morphological studies and confocal imaging. Figure 2. A neurostimulator is placed in the epidural space under fluoroscope. Inset shows device in comparison to a fingertip. Figure 4. ISLET architecture with beta-cells stained dark in: (4A-C) SD rats treated with RTX. Please note the disruption of ISLETS in RTX-treated acute rats and virtual abolition of ISLETS in the RTX-treated 3-day chronic rat and (4D-4E) the ZL rat, used as a control against the genetically altered ZF rat, whose beta cells are marked with compensatory enlargement and cellular disruption. Calibration = 100 microns. The Islets of Langerhans (ISLETS) of the pancreas are richly innervated by SSNFs (C and A - fibers) which emanate from the dorsal root ganglion (DRG) of T9-T11 with central axons terminating in the lateral dorsal horn (DH) and traversing the dorsal column of the spinal cord 1 . A high percentage of SSNF from DRG cells contain the neurotransmitter calcitonin gene-related peptide (CGRP) which is transmitted to ISLETS. These SSNFs are generally capsaicin-sensitive neurons encoding TRPV1. The phenotype of DRG neurons of SSNFs innervating ISLETS are considerably different than those that innervate skin 2 . The pancreatic SSNFs are dominated by CGRP, TRPV1, and glial cell line derived neurotrophic factor (GDNF) with values at least two times those of skin. CGRP participates in the regulation of ISLET cell function and can be a key modulator of ISLET stress due to obesity-related SSNF neuropathy 3 or the induction of SSNF neuropathy by RTX 4 . Electrical stimulation of the DC, DRG, and mixed sensori-motor nerves causes antidromic activation of SSNFs, particularly C-fibers expressing TRPV1. Excitation of TRPV1 neurons elicits neurogenic vasodilatation in both cutaneous and visceral microvessels, mediated by CGRP 5 . CGRP release can be influenced by noradrenaline (NA) sympathetic nerves in a frequency-dependent fashion 6 . Conversely, CGRP modulates NA sympathetic outflow 7 , neuronal nitric oxide 8 , and insulin action 9 . In SSNF neuropathy, impaired insulin action associated with down regulation of the insulin receptor (IR) and insulin growth factor 1 receptor (IGF-1R) may be a fundamental factor in loss of neurotrophic support of the TRPV1 neurons innervating the ISLET as well as be a factor observed in obese subjects with SSNF neuropathy 3 . The mechanism underlying vasodilatation from SCS has been attributed to TRPV1 neurons releasing CGRP 10,11 . Low frequencies have shown to be effective in eliciting C-fiber discharge and neurogenic vasodilatation in most neural tissue (DRG, sensory, mixed nerves) 13 . Stimulation at low frequencies elicits a marked capsaicin-sensitive vasodilatory response in most sensory tissue 6 . Introduction 1 Won et al 1998; Su et al 1987; Wang and Westlund 2001. 2 Fasanella et al 2008; Bennett et al 1996. 3 Herman et al 2006. 4 Szallasi and Blumberg 1996. 5 Kawasaki et al 1988; Holzer 1991, 1995, Escott et al, 1995; Khalil and Helme 1996; Croom et al 1997; Wu et al 2006 . 6 Khalil et al 1997. 7 Oh-hashi et al 2001. 8 Holzer et al 1995a. 9 Sima and Kamiy 2006. 10 D ’Luzansky 2000; Cook 1973. 11 Wu et al 2008. 12 Mehri et al 1998; Escott et al 1995; Santicioli et al 1992. 13 Willis, 1999. 14 Wardle et al 1997. Figure 3. EMG signal at 100 microvolts threshold criteria. Once reached, amplitude was adjusted to 80% to threshold levels. RTX Treated (n=5) Algesic substances applied to the skin and transcutaneous electrical stimulation provoke a neurogenic vasodilatory response due to excitation of the SSNF pathways involving dorsal root reflexes (DRRs) 3 . These capsaicin-sensitive pathways express TRPV1. Likewise, SCS elicits a neurogenic vasodilatory response by activating the DRR 13 inducing antidromic excitation of the SSNFs innervating microvessels of the target tissue. In this study we examined whether the beta cells of the pancreas are disturbed in two experimental conditions associated with diminished neurogenic vasodilatation and TRPV1 innervation- obesity and RTX-treatment. The following observations were made: (1) SCS caused higher abdominal skin BF at 5 Hz rather than 100 Hz (Fig. 5). This can be ascribed to NA sympathetic over activity in the presence of a 100 Hz signal; a similar pattern was observed with stimulation of peripheral sensory nerves 6 . This is further inferred by the observation that RTX-treatment did not alter negative flow; (2) ZF rats did not exhibit negative BF suggesting impairment of sympathetic nerve fibers. The magnitude of BF in the obese rats was only minimally decreased, perhaps due to the relatively young age of the rats and/or to residual vasodilatory molecules released by SCS. The morphological studies revealed marked disruption of the ISLETS in the ZF rat (Fig. 4E) with some evidence of changes in neuronal innervation (not illustrated); (3) RTX-treatment deletes most of the TRPV1 neurons at the dose used in this study caused both a marked decrease in vasodilatation and a disruption of the ISLETS (Fig. 4C, 5). The shift to a higher threshold of current to elicit a MT and a neurogenic vasodilatation would be caused by the suppressive action of RTX on both central and peripheral TRPV1 pathways. 14 (4) SCS treatment elicits an insulin action from the beta cell after treatments of 45 minute duration (Fig 6). The studies revealed that C-fiber ISLET neuropathy causing beta-cell stress can be addressed therapeutically by SCS at frequencies modulating ISLET function and insulin production through similar pathways used for vasodilatation of the pancreatic and visceral tissue in obese animals. Figure 5. Abdominal BF analyzed by measuring the area under the curve (AUC) of 90 sec 5 Hz (exciting) and 100 Hz (blocking) stimulation of ZL (n=6) and ZF (n=9) and RTX treated (n=5) rats at 0.2 ms or 1 ms pulse duration. Decreased BF at 100 Hz can be attributed to the negative BF (see upper right panel) which appears before positive flow occurs. Negative flow is not observed in ZF rats..) At the pre-treatment MT, RTX abolishes BF. However, when current intensity increases by 2-4x pre-treatment threshold, a new MT threshold value is observed but the BF responses are substantially attenuated. The negative BF responses at 100 Hz are not altered by RTX-treatment. 4A 4B 4C 4D 4E ZL (n=6) and ZF (n=9) Figure 1. A SSNF containing TRPV1 neurons (e.g. C-fibers) is modulated by SCS acting on the DRG through volume conduction from a DC placement of an electrode lead. The neurotransmitter CGRP is released stimulating insulin action within the beta cell. SCS treated animals show a marked effect on insulin production. 5 Hz Conclusion Figure 6. Representative islets from a ZL control rat, a ZL SCS treated rat, a ZF control rat, and a ZF SCS treated rat. Notice the more obvious staining for glucagon and PGP 9.5 after SCS in the ZL animals, the healthier appearance of the islet as a whole and the more obvious staining for insulin after SCS . This data suggests that SCS results in greater PGP9.5 and glucagon staining in the islets of ZL rats and greater insulin staining in the islets of ZF pre‐diabetic rats . Green: insulin (beta cells); White: glucagon (alpha cells); Red: PGP9.5 (neurons); Scale bar: 75 um. 4E Control “Healthy” Islets RTX Acute RTX Three Day