The document describes research investigating how the protein amyloid-beta (Aβ) may amplify itself in Alzheimer's disease. It finds that Aβ inhibits the enzyme ADAM10, reducing cleavage of amyloid precursor protein (APP) and allowing more amyloidogenic processing and Aβ production. It also finds that the protein netrin-1 can disrupt this process, reducing Aβ levels both in vitro and in mouse models. Testing netrin-1 and netrin-mimetic peptides as potential Alzheimer's therapeutics is discussed.


![224 P.R. Spilman et al. / Netrin-1 Disrupts Aβ Amplification
A1-42 and A1-40, and ICV delivery of netrin-1 to PDAPPSwe/Ind
mice increased sAPP␣, decreased A, and improved
working memory. Finally, to support further study of netrin-1’s potential as a therapeutic for Alzheimer’s disease, pilot gene
therapy studies were performed and a netrin mimetic peptide synthesized and tested that, like netrin, can increase sAPP␣
and decrease A1-42 in vitro. Taken together, these data provide mechanistic insights into A self-amplification and the ability
of netrin-1 to disrupt it.
Keywords: A1-42, APP, amplification, CED, inducible, mimetic, netrin-1, sAPP␣
INTRODUCTION
The number of cases of Alzheimer’s disease
(AD), the most prevalent age-associated dementia,
is expected to increase rapidly as society ages, and
currently there are no pharmaceutical treatments
that prevent onset or alter the course of the dis-
ease. AD is a progressive neurodegenerative disorder
characterized by the presence of senile plaques, com-
posed mainly of amyloid- peptide (A) [1, 2],
and the development of neurofibrillary tangles of
hyperphosphorylated tau (ptau) in brain tissue [3, 4].
AD and many other neurodegenerative diseases are
considered proteinopathies sharing biophysical and
biochemical characteristics with prion diseases [5] in
that key proteins implicated in the pathophysiology
of the disease apparently self-amplify. Understanding
the mechanisms underlying A’s initial production,
self-amplification, accumulation, and spreading—as
well as endogenous factors that may abrogate such
increases—may well inform new therapeutic strate-
gies for AD.
Previous studies have shown that addition of A
peptides to cells [6] results in an overall increase
in A concentration, supporting a hypothesis that
A is self-amplifying. In vivo studies confirm this,
revealing that inoculation of AD-susceptible mice
with natural or synthetic A increases and acceler-
ates A1-40 and A1-42 accumulation [7, 8]. While
the exact mechanism(s) for A increases under these
conditions are unknown, it likely involves increased
amyloidogenic processing of full-length APP (FL
APP) or reduced degradation of A. FL APP
may be processed proteolytically via two mutually
antagonistic pathways. Cleavage by BACE1 (BACE)
at the  site produces sAPP and CTF, the
latter of which can then undergo ␥ cleavage to pro-
duce A of various species (lengths) [9–11]. FL
APP can also undergo caspase cleavage, generat-
ing APPneo (APP C31) and C31 [12–16]. The
afore-mentioned peptides mediate neurite retraction,
synapse loss, and ultimately programmed cell death.
In contrast, cleavage by ADAM10 (a disintegrin and
metalloprotease) at the ␣ site results in production
of trophic peptides sAPP␣ and ␣CTF that mediate
neurite extension, synaptic maintenance, and anti-
apoptotic events [12, 17]. Thus APP may function
as a molecular switch involved in plasticity.
The “decision” between these two APP cleavage
pathways may be influenced, in part, by ligand bind-
ing. We and others have previously shown that the
A peptide interacts with APP in the A cognate
region [18, 19], and that this results in an increase in
the caspase cleavage of APP at Asp664 [20]; thus,
A acts as a ligand and directs APP to a particular
cleavage pathway. Of direct relevance here, previous
work has also shown that the axonal guidance and
trophic factor netrin-1 can also act as a ligand [14,
21]. The binding of netrin-1 to APP was revealed
by co-immunoprecipitation (IP) and recruitment of
netrin-1 to APP-expressing (but not APP null)
B103 cells. This interaction was confirmed in physi-
ological systems using primary neuronal cultures and
endogenously expressed proteins. Further IP experi-
ments narrowed the binding region to between the ␣-
and -cleavage sites of APP, or the N-terminal 17
amino acids of A. This earlier work also revealed
that netrin-1 exerts a functional effect on APP pro-
cessing in the form of enhancing Fe65 binding to
APP and AICD-dependent gene transcription.
The reasons for undertaking the studies presented
here were twofold; first to elucidate a mechanism
for A self-amplification and second to determine if
netrin-1, known to bind both FL APP and A itself
[14], could disrupt this self-amplification in vitro and
in vivo. To achieve these goals, a variety of in vitro
and in vivo studies were performed, with in vitro
studies focused on the effects of exogenously sup-
plied A, netrin-1, and ADAM10 on APP-derived
peptides, particularly A, in a variety of cell lines;
and in vivo studies in mice designed to determine
effects of netrin-1 increased by either transgenic over-
expression or delivery of exogenous protein on APP
processing, and to ascertain further its potential as an
AD therapeutic. With the same goal of studying the
therapeutic potential of netrin-1, we also performed](https://image.slidesharecdn.com/9d4f4bed-5158-4741-be4b-0954b89c28d3-161106220247/85/Spilman-Corset-Netrin-paper-2-320.jpg)
![P.R. Spilman et al. / Netrin-1 Disrupts Aβ Amplification 225
pilot studies of convection-enhanced delivery (CED)
of an adeno-associated virus 2 (AAV2) netrin-1-cmyc
construct and designed, synthesized, and tested sev-
eral cyclic peptide netrin mimetics in vitro.
In future studies, CED of carefully-designed con-
structs will allow us to ascertain netrin effects in a
variety of AD models, and perhaps even in models
of other neurodegenerative diseases such as amy-
otrophic lateral sclerosis wherein A has recently
been implicated [22]. We will also determine the in
vivo effects of the netrin peptide mimetic described
here as well as similar peptides, since mimetics—due
to their relative ease of delivery—may hold the most
promise as therapeutics.
MATERIALS AND METHODS
Cell culture and transfection conditions
B103 (rat neuroblastoma) cells were cultured
in Dulbecco’s modified Eagle’s medium (DMEM)
containing 10% fetal bovine serum and 1% peni-
cillin/streptomycin. Transient transfection of B103
cells with APPSwe/Ind was performed using the
Lipofectamine LTX reagent (Life Technologies) for
48 h according to the manufacturer’s instructions.
24 h before assay, the cells were cultured without fetal
bovine serum and recombinant human A1-42 pep-
tidewasaddedtothemedia.H4(humanneuroglioma)
cells were cultured similarly, but without the 48-h
transfection.
Plasmids
The construct for full length APPSwe/Ind
(pcDNA3- APPswi) was provided by the Glad-
stone Institutes (UCSF) and netrin-1 (pGNET1-myc)
was from the laboratory of Patrick Mehlen [23]. The
MBP-APPC125constructwasobtainedbyfusionof
the C-terminal 125 residues of APP Swedish vari-
ant with the C terminus of maltose binding protein
(MBP) in Escherichia coli expression vector pMALc
(New England Biolabs).
Recombinant protein and peptides
Recombinant human netrin-1 was purchased
from Apotech, while recombinant human peptides
A1-42, A42-1, FAM-Biotin A1-42, and rat A1-40
were purchased from Anaspec. ADAM10 and TIMP1
were purchased from Sigma-Aldrich. The cyclic pep-
tides were prepared at Anaspec using solid phase
synthesis. First, the linear tripeptide netrin-loop
sequence with flanking N and C-terminal cysteines
were prepared on resin. The linear peptide synthe-
sis was terminated with a dansyl N-terminal group.
After cleavage of the peptide from the resin, it was
cyclized in solution as described in Eichler and
Houghten [24]. The internal disulfide bond was mon-
itored using Ellman’s reagent. The cyclic peptide was
then purified using standard reverse phase HPLC
to obtain the cyclic peptides in >95% purity. The
cyclic peptides had the following sequences: pep-
tide 1 - dansyl-Cys-Ile-Asp-Pro-Cys (with S-S bond);
peptide 2 - dansyl-Cys-Val-Ala-Gly-Cys (with S-S
bond);peptide3-dansyl-Cys-Pro-His-Phe-Cys(with
S-S bond); peptide 4 - dansyl-Cys-Pro-Asp-Phe-Cys
(with S-S bond); peptide 5 - dansyl-Cys-Val-Asn-
Ala-Cys (with S-S bond); and peptide 6 - dansyl-
Cys-Ile-Pro-Asp-Cys (with S-S bond).
Immunoblotting and immunoprecipitation
B103 cells were lysed in RIPA buffer in the pres-
ence of complete protease inhibitor cocktail (Roche)
on ice for 60 min, followed by removal of cell debris
by centrifugation. Electrophoresis and immunoblot-
ting were performed using lysates as previously
described [14], and anti-APP C-terminal epitope
(Sigma-Aldrich), anti-sAPP␣ (IBL), anti-netrin-1
(R&D Systems), and anti-actin (Chemicon) anti-
bodies. Co-immunoprecipitations from B103 cell
supernatants were performed using anti-sAPP␣
or anti-APP N-terminal epitope (Sigma-Aldrich)
antibodies. Densitometry was performed using the
ChemiImager 4400 (Alpha Innotech) with Image J
software.
Primary neuronal culture
Embryos from PDAPP (J20) mice described else-
where [25] were used to generate primary neuronal
cultures. Hippocampi from E18.5 embryos from
PDAPP (J20) × PDAPP (J20) crosses were iso-
lated. Genotyping was performed a posteriori to
control for the percentage of embryos positive for the
transgene in each culture. Hippocampi were chem-
ically dissociated using trypsin (Sigma-Aldrich),
DNA digested with DNaseI (Sigma-Aldrich) and
neurons cultured in Neurobasal media supple-
mented with B27, 1 mM Glutamax and 1%
Penicillin/Streptomycin (Life Technologies), in 6
well plates pre-coated with 10 g/ml poly-L-lysine.
Cultures were maintained for 8 days. At day 1](https://image.slidesharecdn.com/9d4f4bed-5158-4741-be4b-0954b89c28d3-161106220247/85/Spilman-Corset-Netrin-paper-3-320.jpg)
![226 P.R. Spilman et al. / Netrin-1 Disrupts Aβ Amplification
(1 day after dissection), medium was replaced, and
each day following, human netrin-1 (6 nM) or rat
A1-40 (440 nM) were added. At day 8, media
supernatants were collected, and sAPP␣ and human
A1-42 were measured.
sAβPPα and Aβ determination
For all but H4 cells, sAPP␣ was quantified by
immunoblot using the specific 2B3 antibody (IBL)
normalized against actin or APP loading controls.
AlphaLISA (Perkin-Elmer) was used to measure
sAPP␣ in H4 supernatents. A was determined by
ELISA (Life Technologies) in all experiments. For
primary hippocampal neuronal cultures, in order to
measure total human A1-42, protein from super-
natantswasprecipitatedwithmethanolandthepellets
resuspended in 5M guanidine and assayed by ELISA.
A1-42 was also measured by ELISA for cell lysates.
ADAM10 in vitro assays
All ADAM10 digests were performed for 1 h at
37◦C in ADAM10 reaction buffer, pH7.4, according
to the manufacturer’s instructions (Sigma-Aldrich).
Recombinant peptides A1-42, A42-1, and FAM-
Biotin A1-42 were resuspended in PBS with 0.06%
NaOH before use. ADAM10 and TIMP1 were
resuspended according to the manufacturer’s instruc-
tions (Sigma-Aldrich). MBP-APPC125 cleavage
was visualized by immunoblot using an anti-sAPP␣
2B3 antibody (IBL), and the intensity of the cleavage
product band was quantified using using the Chemi-
Imager 4400 (Alpha Innotech) and Image J software.
ADAM10 cleavage of recombinant FAM-Biotin
Aβ1-42
Recombinant FAM-Biotin A1-42 was used to
determine cleavage of A1-42 by ADAM10. The
biotin residue is located in the C-terminus. The reac-
tion is the same as described above. The efficiency
of ADAM10 cleavage was controlled by quantifying
the cleavage of MBP-APPC125 by immunoblot as
described above. An ELISA assay was developed to
quantify the cleavage of FAM-Biotin A1-42. Fol-
lowing cleavage, the reaction mix was loaded into
wells of high-sensitivity streptavidin 96-well plates
(Pierce). FAM-Biotin A1-42 was allowed to bind for
3 h at room temperature with shaking. Then, the plate
was incubated with the 6E10 antibody (Covance),
which only recognizes the uncleaved FAM-Biotin
A1-42, as the epitope is on the N-terminus. Sec-
ondary anti-mouse HRP antibody (Santa Cruz) was
then added and the amount of uncleaved FAM-Biotin
A1-42 was quantitated by colorimetric assay using
SIGMAfast OPD on a SoftMax Pro Spectramax 190
plate reader (Molecular Devices).
MBP-AβPPC125 fusion protein production and
purification
Constructs for maltose binding protein (MBP)
fused to the C-terminus of the APPSwe C-terminal
125aa sequence were cloned into the expression vec-
tor pMalc (New England Biolabs) and E. coli were
transformed. The MBP-APPC125 fusion protein
was induced in bacterial cultures and purified by
affinity chromatography using amylose-agarose. The
bound proteins were eluted with 10 mM maltose in
20 mM Tris, pH 7.5, 1% Triton X-100, 0.1M NaCl,
diluted 1:1 with 6 M guanidine, and stored in frozen
aliquots (0.1 mg/ml fusion protein in 10 mM Tris, pH
7.5, 0.2% Triton X-100, 0.15 M guanidine-HCl) as
previously reported [26].
Generation of transgenic mice and netrin
induction
All mouse experiments were performed in accor-
dance with an Institutional Animal Care and Use
Committee (IACUC) approved protocol at the Buck
Institute for Research on Aging. The netrin-inducible
line was created by co-injection of two DNA con-
structs into ova; the first was inducible Cre [27]
wherein the rtTA was under the control of the
neuron-specific enolase (NSE) promoter and the
second construct comprised a prion (Prp) promoter-
loxp-Luciferase-stop-loxp-Netrin-1. Treatment with
doxycycline (Dox) and expression of CRE allows
excision of the luciferase sequence, recombination,
and therefore read-through and expression of netrin-
1. In the netrin-inducible line used here, the genes
stably sort together.
Study design
The bi-transgenic mice were crossed with
PDAPP J20 mice described elsewhere [25, 28]
which express human APP with both the Swedish
and Indiana mutations, the former increasing BACE
cleavage of APP and the latter increasing A1-42
production relative to other A species. Such
crosses generate non-transgenic, single transgenic](https://image.slidesharecdn.com/9d4f4bed-5158-4741-be4b-0954b89c28d3-161106220247/85/Spilman-Corset-Netrin-paper-4-320.jpg)
![P.R. Spilman et al. / Netrin-1 Disrupts Aβ Amplification 227
(APP), bi-transgenic (Net), and tri-transgenic mice
(APP/Net). As non-transgenic and bi-transgenic
mice do not express huAPP, they were not used in
the studies presented here. Single transgenic APP
and triple transgenic (APP/Net) mice were sorted
into groups and received either non-Dox or Dox-
containing chow (200 mg per kg food, Bio-Serv)
starting at two months of age for two weeks. Several
rounds of cross-breeding were required to generate
enough mice for each group to power the study to
reach statistical significance. For A1-40 and A1-42
analysis,APPdoxn = 9,APP-Netdoxn = 9,APP
n = 8, and APP-Net n = 9. In sAPP␣ analysis, n
remained the same for APP dox and APP-Net dox,
but dropped in to n = 6 and n = 7 for APP and APP-
Net, respectively as very low outliers were removed.
In pilot studies, the increase in netrin-1 in APP-
Net dox as compared to APP dox was determined
by immunoblot. Brain homogenates were prepared in
PBS with complete protease inhibitors (Roche). Lim-
iting dilutions (30, 15, and 7.5 l) were loaded onto
Bis-Tris gels (Novex) which were then transferred to
PVDF membrane, probed with anti-netrin-1 (R&D
Systems), and after incubation with HRP-conjugated
secondary antibody, developed with chemilumnes-
cent substrate and exposed to film.
Biochemistry in brain tissue
Hemibrains minus cerebellar tissue were sonicated
by weight into 5 M guanidine, 50 mM Tris, pH 8
buffer and rotated for three hours at room tempera-
ture. huA1-42 and huA1-40 levels were ascertained
by ELISA assay (Biosource, Life Technologies). For
sAPP␣ determination, 100 l of the Gdn sonicate
was precipitated in ice-cold methanol for 4 h and
centrifuged at 5000 rpm for 15 min to pellet the pro-
teins. Proteins were then sonicated in 1X AlphaLISA
Hiblock buffer (Perkin-Elmer) and assessed in the
AlphaLISA assay for sAPP␣ (Perkin-Elmer).
Acute intracerebroventricular injection of mice
Adult non-transgenic mice were anesthetized with
isoflurane and injected intracerebroventricularly into
the right lateral ventricle at bregma 1.2 and 1 mm
from the mid-line. Incision sites were closed with
two sutures and surgical adhesive. A total of 5 l
of 1 g/l netrin-1 (R&D Systems) in saline was
injected over several minutes. Mice were euthanized
by ketamine/xylazine injection at 5 min, 1 h, 6 h,
and 24 h after injection; 3 mice were used for each
time point. Brains were removed and snap frozen
for immunofluorescent labeling. For immunofluo-
rescence, 14 m sections were cut using a cryostat
and allowed to dry for 2 min before post-fixation
in 4% paraformaldehyde for 30 min. Non-specific
antibody binding was blocked with 5% normal
goat serum (NGS, Vectorlabs) in phosphate buffered
saline (PBS) for 1 h followed by incubation with pri-
mary antibody, rabbit anti-netrin-1 (R&D Systems)
at 1:200 in PBS with 5% NGS overnight at room
temperature. Slides with labeled brain tissue sections
were washed with PBS and then incubated with don-
key anti-rabbit AlexaFluor488 at 1:200 in PBS with
5% NGS for 2 h. Sections were washed again and
Prolong Gold with DAPI (Vectorlabs) was used to
mount the slides. Images were taken on a Nikon 800
upright fluorescent microscope at 20X using Act-1
imaging software.
Chronic netrin-1 delivery study design
Netrin-1 was delivered for 28 days by Alzet pump
with the ICV cannula placed in the left lateral ventri-
cle. Pumps were filled with 1 g/l netrin-1 (R&D
Systems) in physiological saline with 1% mouse
serum albumin (MSA, Calbiochem). Mice were
anesthetized with isoflurane and an incision made
between the shoulder blades and the pump assem-
bly, comprising the pump, 1.4 cm of tubing and the
cannula were implanted subcutaneously. The wound
sites were closed with multiple sutures. Mice were
placed on a heating pad during recovery and anal-
gesia administered as needed. Mice were carefully
monitored during the course of the study. Groups
included non-transgenic mice receiving vehicle, J20
PDAPP AD model mice receiving vehicle and J20
mice receiving netrin-1. While each group started
with 12 mice, some mice died during the course of the
study. At the time of behavioral analysis at 2 weeks,
for NTg, n = 11, J20 vehicle, n = 11, and J20 netrin,
n = 10; and at the time of behavioral analysis at 4
weeks and for biochemistry, n = 11, n = 10, and n = 9,
for NTg, J20 vehicle and J20 netrin, respectively.
Novel Object Recognition (NOR) testing
Two and four weeks after the implantation of
the pumps, mice underwent working object memory
testing using the NOR paradigm. The method used
followed that of Bevins and Besheer [29] wherein
the mice were acclimated to the black rectangular
30 × 24 × 22 cm arena with bedding for 15 min/day](https://image.slidesharecdn.com/9d4f4bed-5158-4741-be4b-0954b89c28d3-161106220247/85/Spilman-Corset-Netrin-paper-5-320.jpg)
![228 P.R. Spilman et al. / Netrin-1 Disrupts Aβ Amplification
for two days before the study. During the study, mice
were placed in the arena with two identical objects
for 10 min (acquisition) and then removed for 1 h.
The mice were then exposed to one familiar and
one novel object for 10 min. The movements of the
mice were recorded by video and processed using
Noldus Ethovision software. The distance traveled
was analyzed by the software and the number of inter-
actions (the nose coming within 2 cm of the object)
with each object was scored by an observer of the
video blinded to the genotype and treatment of the
mice.
Determination of sAβPPα and Aβ in mouse
brain tissue after chronic delivery
At the end of the study, mice were euthanized
by ketamine/xylazine anesthesia and brains were
removed, halved down the mid-line, and snap frozen
on powdered dry ice. As the netrin-1 was delivered
unilaterally into the left ventricle in the chronic study,
the left hemibrains were processed and analyzed. To
make it possible to perform both immunofluorescent
labeling of tissue sections and biochemical analysis
of sAPP␣ and A1-40 and A1-42 in the left brain,
frozen, unfixed tissue was cut coronally into 14 m
sections, and starting at the emergence of the ven-
tricle nearest the olfactory bulb. Every 10th section
was mounted on a slide for immunofluorescence and
an equal number of intervening sections placed in a
tube for biochemical analysis. For biochemical anal-
ysis, 100 l of 5M guanidine buffer was added to the
tissue sections from the same bregma and sonicated.
The assays for sAPP␣ and A1-40 and A1-42 were
performed as described above.
Convection-enhanced delivery of an AAV2-
netrin-1-cmyc construct
AAV2 vectors containing complementary DNA
sequences for human netrin-1 (AAV2-netrin-1-cmyc)
under the control of the cytomegalovirus promoter
were packaged by the AAV Clinical Vector Core
at Children’s Hospital of Philadelphia as previously
described [30, 31]. AAV2-netrin-1-cmyc stocks were
diluted immediately before use to equivalent titers
of 1–1.2 × 1012 vector genomes/ml in phosphate-
buffered saline with 0.001% (vol/vol) Pluronic F-68.
Under surgical anesthesia [ketamine:xylazine (1:1);
2 ml/kg], three adult (>3 months of age) received
CED delivery of a CMV-AAV2-netrin1-cmyc
construct. Each mouse was placed in a stereo-
tactic frame (Stoelting), an incision made in the
skin overlying the skull and a hole made in the
skull at the infusion site. A custom-made silica
cannula was used to infuse 2 l of AAV2-netrin1-
cmyc (1 × 1013 vector genomes/ml) unilaterally into
the right hippocampus by programmable microsy-
ringe pump (World Precision Instruments; flow rate
0.5 l/min; stereotactic coordinates from bregma:
AP –2 mm, ML +1.5 mm, DV –2 mm). After
CMV-AAV2-netrin1-cmyc administration, the scalp
was closed with sutures, and 30 days later mice
were euthanized by ketamine:xylazine anesthesia.
Brains were removed and snap frozen on pow-
dered dry ice, sectioned at 14 m and post-fixed in
4% paraformaldehyde for immunolabeling. Sections
were incubated with rabbit monoclonal anti-cmyc
antibodies(Abcam,ab32072)andafterwashing,anti-
rabbit Alexafluor488 secondary. Microscopy and
imaging were performed as described above for acute
netrin-1 injection.
Treatment of CHO-7W cells with netrin cyclic
peptides
CHO-7W cells stably transfected with huAPPwt
and grown in Dulbecco’s Modified Eagle’s Medium
(DMEM) supplemented with 10% fetal bovine serum
and 1% penicillin-streptomycin, were seeded at
a density of 5 × 104 cells/well in 96-well plates
overnight and 24 h later, treated with netrin cyclic
peptides 1–6 at 1 M. As the original stock solution
was in DMSO, both DMSO and no DMSO controls
were also run. After treatment for 24 h, cell media
were collected and assayed for sAPP␣ using the
Perkin-Elmer AlphaLISA and for A1-42 using Invit-
rogen’s ultrasensitive A1-42 ELISA kit according to
the manufacturer’s instructions. The ratio was cal-
culated by dividing the sAPP␣ reading in arbitrary
units (AU) by the A1-42 reading in pg/ml for indi-
vidual wells.
Determination of ERK phosphorylation by netrin
mimetic cyclic peptides
Deleted in colorectal cancer (DCC)-expressing
B103 rat neuroblastoma cells were transfected
with a vector expressing APP (pcDNA3-human
APP695wt) using Lipofectamine 2000 reagent
(Invitrogen) according to the manufacturer’s instruc-
tions. The following day, cells were treated with
netrin-1 and cyclic peptides at 150 ng/ml for 5 min.
Cells were then harvested and lysed, and equal](https://image.slidesharecdn.com/9d4f4bed-5158-4741-be4b-0954b89c28d3-161106220247/85/Spilman-Corset-Netrin-paper-6-320.jpg)
![P.R. Spilman et al. / Netrin-1 Disrupts Aβ Amplification 229
amounts of total protein were loaded onto 15% poly-
acrylamide gels for electrophoresis. Western blotting
was performed using anti-APP, anti-ERK1/2, and
anti-phospho ERK (Cell Signaling) antibodies. In a
follow-up experiment, peptide 1 alone was used and
the same experiment performed, but in comparison to
transfection with an empty pcDNA3 vector and with
0, 5, and 15 min time points.
Surface plasmon resonance: Binding of
netrin-1-derived peptides to TRX-eAβPP575-624
SPR data was obtained with a Biacore T100.
CM5 chips were cross-linked with either TRX-
eAPP575-624 or TRX (thioredoxin) [19]. Two
different pHs were tested for binding: pH 6.7 and
pH 7.4. Sensograms were obtained for peptide 1 for
0–16.7 M peptide pumped at 20 l per min in 1%
DMSO, 20 mM sodium phosphate pH 6.7, 125 mM
sodium chloride, 0.05% Tween 20 over a flow chan-
nel containing TRX-eAPP575-624 (20 kDa) and the
responses at equilibrium for all three peptides were
compared. The response for peptides 1 and 2 was
not significantly different from the flow channel con-
taining TRX for concentrations below 10 M. The
responses were calculated using a double subtraction
method using a mock cross-linked reference chan-
nel and buffer standards. Because of the relatively
small responses, buffer standards were created for
each of the steps of the serial dilution in order to
get a better estimate of the effect of small deviations
in the DMSO concentrations between the sample
and the running buffer. PRISM (GraphPad Inc.) was
used to fit a single-site saturation binding model in
which the background and the non-significant bind-
ing contributions were constrained to be the same for
all three proteins, but the KD was allowed to vary.
The KD was estimated to be 0.018 M with a 95%
confidence interval of 0.015–0.021 M for peptide
1. The calculated KDs for peptides 2 and 3 were
greater than 10 M consistent with a lack of signal
for these peptides in the concentration range used for
the experiment. R2 was greater than 0.95 for pep-
tide 1. Sensograms were also obtained for peptide
1 pumped at 20 l per min in 1% DMSO, 20 mM
sodium phosphate pH 7.4, 125 mM sodium chlo-
ride, 0.05% Tween 20 for concentrations 0–14 M,
as was the response at equilibrium for peptide 1
at pH 7.4. The calculated KD was estimated to
be approximately 30 M, which suggests that the
binding of peptide 1 to TRX-eAPP575-624 is pH
dependent.
RESULTS
In vitro
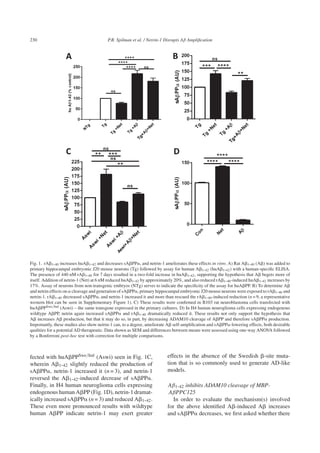
rAβ1-40 increases huAβ1-42 in primary mouse
hippocampal neurons
In order to determine the effect of A peptide
on the processing of APP, we added rat A1-40
(rA1-40) to primary cultures of hippocampal neu-
rons derived from J20 PDAPPSwe/Ind transgenic
mouse embryos. J20 mice express human APP
under the control of the platelet-derived growth factor
promoter with both the “Swedish” (KM670/671NL)
and “Indiana” V717F mutations, the former increase
BACE1 cleavage of APP and the latter increases
production of A1-42 compared to other A species
(lengths). After addition of rA1-40 to the primary
neurons, we then assayed human A1-42 with an
ELISA assay that is specific for huA1-42. As shown
in Fig. 1A, the presence of rA1-40 at a concen-
tration of 440 nM for 7 days led to a greater than
two-fold increase in huA1-42 (n = 9), supporting the
hypothesis that A begets more of itself. However,
exposure of neurons to 6 nM netrin-1 over the same
time period reduced huA1-42 by 20%, and lessened
the rA1-40–induced increase in huA1-42 by ∼17%,
although not significantly so. These findings confirm
and extend our previous results showing that netrin-
1 reduces net A production (here, just a trend) in
organotypic slice cultures from J20 mouse brain [14].
Thus, the addition of exogenous A (in this case,
rA1-40) increases net A production from APP
(here, huA1-42) and netrin-1 shows a trend to reduce
huA1-42 production.
Aβ1-42 reduces, and netrin-1 increases, sAβPPα
in primary neurons, huAβPPSwe/Ind-transfected
B103 cells, and H4 cells
As noted above, FL APP may be processed in
two mutually antagonistic ways, one of which results
in the formation of A and the other of which results
in the formation of sAPP␣. Moreover, since the ␣
cleavage site lies within the A peptide region of
APP, cleavage at that site precludes the production
of A. Therefore, we next evaluated the effects of
A and netrin-1 on sAPP␣ production. As shown
in Fig. 1B, in primary hippocampal neuronal cul-
tures derived from J20 AD model mice, rA1-40
decreased sAPP␣ by ∼15%, netrin-1 significantly
increased it (n = 9), and netrin-1 increased sAPP␣
levels in the presence of ratA1-40. Results were
very similar with B103 rat neuroblastoma cells trans-](https://image.slidesharecdn.com/9d4f4bed-5158-4741-be4b-0954b89c28d3-161106220247/85/Spilman-Corset-Netrin-paper-7-320.jpg)
![232 P.R. Spilman et al. / Netrin-1 Disrupts Aβ Amplification
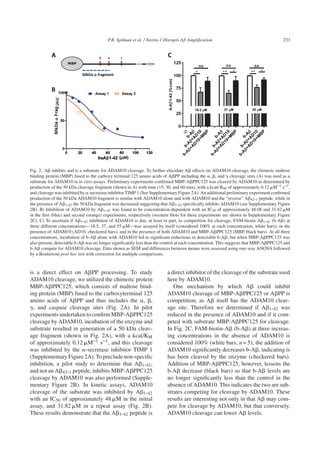
In vivo
Induced netrin-1 expression decreases Aβ in
triple transgenic mice
In order to test the ability of netrin-1 to interrupt
A amplification in vivo, a tet-on double trans-
genic mouse line was established wherein gene 1
comprises the rtTA expressed under control of the
neuron-specific enolase (NSE) promoter and Cre
recombinase (Cre) under the control of the CMV pro-
moter (Fig. 3A). Upon treatment with doxycycline
(dox), Cre is expressed and then excises a luciferase
reporter sequence bracketed by lox p elements in
gene 2 (Fig. 3B), allowing recombination, read-
through, and expression of netrin-1 under the control
of the prion promoter. These mice were crossed
with J20 mice (Fig. 3C) that express human APP
(huAPPSwe/Ind) with KM670/671NL and V717F
mutations. A levels are low in these mice before 3-4
months of age, and then increase exponentially from
4 months on, resulting in plaque formation in most
mice by 7 months of age [25]. To ascertain impact
on A amplification, mice were studied during this
pre-plaque amplification window.
Crosses of the double transgenic mice to J20
PDAPP mice potentially produce mice positive for
huAPP (APP), for APP and inducible netrin-1
(APP-Net), for Net only (Net), and non-transgenic
mice. As the biochemical assays require the pres-
ence of huAPP, only those mice positive for
huAPP were used in studies. To ascertain effects
of netrin induction, some mice received doxycy-
cline for two weeks in chow starting at two months
of age, and control mice did not. All mice were
euthanized at 5 months of age. Quantification of
western blots performed in pilot studies to con-
firm increased expression of netrin-1 in dox-treated
APP-Net mice showed 3 to 4-fold increases as com-
pared to control (Fig. 3D). In study mice, A1-40
and A1-42 brain levels were measured by ELISA
and sAPP␣ levels by AlphaLISA, as described
in “Methods”. sAPP␣ was unchanged in APP-
Net dox mice as compared to all other groups, but
both A1-40 and A1-42 were decreased signifi-
cantly (Fig. 3E, F, and G, respectively). Increases
in the A1-40/A1-42 (slight), sAPP␣/A1-40, and
sAPP␣/A1-42 ratios—all thought to reflect a more
trophic state—were seen in APP-Net dox mice as
compared to other groups (Fig. 3H, I, and J, respec-
tively) supporting the in vitro finding that netrin-1
inhibits A amplification.
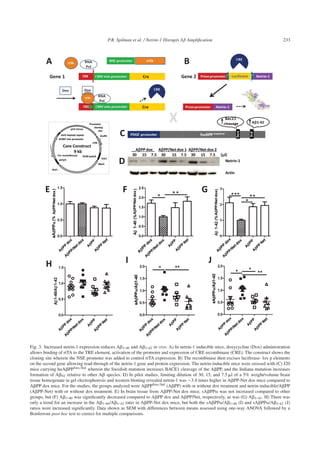
Recombinant netrin-1 diffuses into tissue after
acute ICV delivery
Before commencing chronic pump delivery stud-
ies, it was important to determine if netrin-1, a
relatively“sticky”proteinpronetoaggregation,could
pass through the ventricle wall into brain tissue.
Therefore, a study was performed wherein 5 l of
recombinant netrin-1 at 1 g/l in a saline-BSA vehi-
cle was injected acutely into the right lateral ventricle
of mice. This study played the role that a pharmacoki-
netic study would for a chemical compound, serving
to show if the potential therapeutic can enter the target
organ.ThesiteofinjectionisshowninFig.4A(insert,
and red arrow). Injected mice were euthanized at var-
ious time points, and at 5 min post-injection, netrin-1
can be seen lining the ventricle wall (Fig. 4A). At 1 h,
it could be seen diffusing into the surrounding tissue
(Fig. 4B, area imaged red box in insert) and this dif-
fusion was the greatest, of the time points measured,
6 h after injection both in the lateral and third ven-
tricles (Fig. 4C and D, respectively, area imaged red
box in insert). In the ventricle contralateral (left) to
the injected ventricle, at 6 h, netrin-1 was only seen
lining the walls (Fig. 4E). By 24 h post-injection, only
residual netrin-1 remained lining the ventricle walls
(Fig. 4F). Again, while the images of netrin-1 diffu-
sion after acute injection do not represent the level
of netrin-1 at any given time during chronic infusion,
they gave evidence that netrin-1 when delivered to
the ventricle can enter the tissue, and therefore that
chronic infusion studies would be worthwhile.
Chronic ICV netrin-1 delivery improves working
object memory
After J20 mice received ICV delivery of 1 g/l
netrin-1 by Alzet osmotic pump at 0.11 l/h for 2
weeks, they underwent working memory assessment
in the NOR testing paradigm. As part of NOR test-
ing, mice are exposed to two identical objects for
10 min in the acquisition phase. This serves not only
to introduce objects to remember, but validates the
method in that mice should interact equally with both
objects (“right” and “left”), as the mice in this study
did (Fig. 4G). The total interactions with both objects
also reflect activity level and disinhibition. There was
a trend to a decrease in the hyperactivity that is part
of the J20 phenotype as reflected by the total interac-
tions with identical objects in testing (Fig. 4H). The
hyperactivity of J20, and many other AD model mice,
may impact performance in this, and many memory
assessment paradigms, and thus a therapeutic that can](https://image.slidesharecdn.com/9d4f4bed-5158-4741-be4b-0954b89c28d3-161106220247/85/Spilman-Corset-Netrin-paper-10-320.jpg)
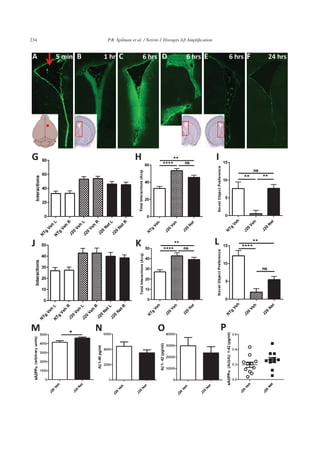
![P.R. Spilman et al. / Netrin-1 Disrupts Aβ Amplification 235
lower activity may well increase performance. Also
at 2 weeks, working object memory as reflected by
increased novel object preference in the NOR task
was significantly improved in netrin-treated J20 mice
(Fig. 4I). After 4 weeks of delivery, there was still a
trend toward lowered activity in netrin-treated J20
mice (Fig. 4K), but the improvement in memory in
these mice was no longer significantly different from
vehicle-treated J20s (Fig. 4L). Post-study analysis of
brain tissue for evidence of netrin-1 and of residual
volume in the pumps revealed that netrin delivery had
failed in some mice with time, likely resulting in the
loss of efficacy seen in those mice.
Chronic ICV netrin-1 delivery increases sAβPPα
and decreases Aβ1-42 and Aβ1-40
Although netrin-1 delivery may have been
impaired with time in some mice, at the end of the
study, netrin-1 treated J20 mice had significantly
increased sAPP␣, decreased (although not signifi-
cantly so) A1-40 and A1-42, and therefore a higher
sAPP␣/A1-42 ratio (Fig. 4M, N, O, and P, respec-
tively). The decreases in A were not significant due
to individual mouse variation. It should be noted what
appear to be minor increases (10%) in sAPP␣ here
result in great improvement in performance in NOR.
Nonetheless, as direct delivery of recombinant netrin-
1 may continue to provide technical challenges, we
also performed pilot studies on a more readily deliv-
erable netrin-1 gene construct and netrin-1 cyclic
mimetic peptides.
CED of an AAV2-netrin-cmyc construct results
in excellent tissue distribution
Exogenously delivered proteins and peptides are
relatively short-lived in brain tissue and ICV/pump
delivery can be associated with pump failure, tis-
sue damage, or other technical issues. Generation
of transgenic mice expressing a protein of interest
is important for ascertaining protein effects, but is
not an option as a human treatment modality. Deliv-
ery of a gene construct, however, may allow not only
more facile testing of protein—here, netrin—effects
in a variety of AD/neurodegenerative disease mod-
els, it holds promise as a treatment modality for
patients [32–34]. Convection-enhanced (slow, pump-
driven) delivery of a drug-, protein-/peptide-, or gene
construct-containing fluid results in greater distribu-
tion volume than conventional injection and use of
the custom-designed cannula results in reduced tis-
sue damage and less regurgitation up the injection
track. In the study performed here, CED of the con-
struct shown in Fig. 5A to the hippocampus (Fig. 5B)
resulted in excellent tissue distribution and netrin
expression in the target region, particularly the den-
tate gyrus and hippocampus (Fig. 5C, D).
Netrin-1 cyclic mimetic peptide 1 binds to
AβPPα and shows some initial biochemical
effects that are similar to netrin-1
As another approach to harness the effects of
netrin-1 as an AD therapeutic, an initial set of six
cyclic netrin-1 mimetic peptides was designed and
Fig. 4. Netrin-1 diffuses into tissue after acute delivery, and improves behavior and biochemical readouts after ICV delivery. To determine
if netrin-1 could transverse the ventricle wall, acute injection of 5 l of 1 g/l recombinant netrin-1 was performed. A) Netrin-1 can be
seen lining the ventricle wall 5 min after injection into the right lateral ventricle (red arrow) at approximately Bregma 0.74 mm, 1 mm from
the midline (insert, red dot). B) By 1 h, it has diffused into the surrounding tissue. The insert shows the imaged area boxed in red. C) Of the
time points measured, the greatest tissue distribution was seen at 6 h post-injection in lateral ventricle near the injection site. D) Netrin-1
can also clearly be seen in the third ventricle 6 h post-injection. The insert again shows the imaged area boxed in red. E) Also at 6 h, some
netrin-1 can be seen lining the lateral ventricle contralateral (left) to the injection site (see insert). F) By 24 h, only residual netrin-1 is seen
lining the walls of the ventricle. All images were taken at 10X and only represent diffusion after acute, not chronic, delivery, but provide
evidence that netrin-1 can exit the ventricle into surrounding brain tissue and therefore support undertaking of subsequent chronic pump
delivery studies. J20 mice received chronic ICV delivery of 1g/l netrin-1 (Net) at a rate of 0.11 l/h for 4 weeks; an additional J20 cohort
and non-transgenic (NTg) mice received vehicle-only (Veh). Working object memory was determined at 2 and 4 weeks. G) At 2 weeks, in
the acquisition phase of training wherein mice were exposed to two identical objects, mice in each group interacted equally with the left (L)
and right (R) objects. H) The total interactions with both objects reveals the increased activity of J20 Veh as compared to NTg Veh mice,
and indicates there was a slight reduction in this activity as a result of netrin-1 treatment of J20 mice (J20 Net). I) Also at 2 weeks, working
object memory as reflected by greater interaction with the novel object (novelty preference) was significantly lower for J20 Veh mice as
compared to NTg Veh mice, but J20 Net mice had improved memory as compared to J20 Veh mice. J) At 4 weeks, while mice in each
group interacted with the two familiar objects equally, the decrease in hyperactivity in J20 Net mice was again only a trend (K). L) Also
at 4 weeks, improvement in memory in J20 Net as compared to J20 Veh was just short of significance; however, the degree of significance
between NTg Veh and J20 Veh mice was greater than that between NTg Veh and J20 Net mice. At the end of 4 weeks of chronic netrin-1
delivery by Alzet pump and implanted ICV cannulae, (M) sAPP␣ was significantly increased in brain tissue and both (N) huA1-40 and
(O) A1-42 decreased, although not significantly; (P) the sAPP␣/A1-42 ratio was increased, but again without significance. Data shown as
SEM with differences between two means were assessed using unpaired two-tailed Student’s t-test, and differences between multiple means
were assessed using one-way ANOVA followed by a Bonferroni post-hoc test to correct for multiple comparisons.](https://image.slidesharecdn.com/9d4f4bed-5158-4741-be4b-0954b89c28d3-161106220247/85/Spilman-Corset-Netrin-paper-13-320.jpg)
![236 P.R. Spilman et al. / Netrin-1 Disrupts Aβ Amplification
Fig. 5. Good distribution achieved after intrahippocampal injection of AAV2-netrin-cmyc in mice. An AAV2-netrin-1-cmyc construct under
the control of the CMV promoter (A) was injected into the hippocampus (red arrow, B) of C57Bl6/J mice using convection-enhanced delivery
(CED). A image representative of fluorescent immunolabeling of the cmyc tag on netrin-1 from the three injected mice from the area boxed
in (B) is shown in (C). It, and the higher magnification image of the boxed area in (C) shown in (D), reveals good expression in the dentate
gyrus and CA3 region of the hippocampus.
synthesized by solid phase peptide synthesis to pro-
vide a mimetic of netrin-1. The structure of netrin-1
mimetic cyclic peptide 1, designed from a loop in the
N-terminal domain of netrin-1, is shown in orange
in Fig. 6A. Cyclic peptides were designed from
netrin loops to mimic the site of active interaction
with the molecule of interest, in this case APP. In
pilot studies of the peptides tested, only peptide 1
phosphorylated ERK in an APP-dependent man-
ner (Supplementary Figure 3A, B) and increased the
sAPP␣/A1-42 ratio in CHO-7W cells stably trans-
fected with human wildtype APP (Supplementary
Figure 3C) similarly to netrin-1.
In addition, as netrin-1 has been shown to bind
APP directly [14] it was of interest to us to see
in cyclic netrin mimetic peptide 1 did as well. The
surface plasmon resonance (SPR) of cyclic peptide 1
obtained with the Biacore T100 is seen in Fig. 6B-E.
Figure 6B shows a sensogram obtained for peptide 1
for0–16.7 Mpeptidepumpedat20 lperminin1%
DMSO, 20 mM sodium phosphate pH 6.7, 125 mM
sodiumchloride,0.05%Tween20overaflowchannel
containing TRX-eAPP575-624 (20 kDa). Compari-
sonoftheresponseatequilibriumforallthreepeptides
tested at pH 6.7 is seen in Fig. 6C. The response for
peptides 2 and 3 was not significantly different from
the flow channel containing TRX for concentrations
below 10 M. PRISM (GraphPad Inc) was used to fit
a single-site saturation binding model in which the
background and the non-significant binding contri-
butions were constrained to be the same for all three
peptides tested. The sensogram obtained for peptide 1](https://image.slidesharecdn.com/9d4f4bed-5158-4741-be4b-0954b89c28d3-161106220247/85/Spilman-Corset-Netrin-paper-14-320.jpg)
![P.R. Spilman et al. / Netrin-1 Disrupts Aβ Amplification 237
Pharmacophore
Peptide 1
EGF Laminin Domains 283-447
(Homology Model using 1 npe)
HMMSTR-Rosetta Model of
Netrin N-terminal domain
All models are from HMMSTR-Rosetta
N-terminal Domain 47-282
Netrin Domain
448-604
N-terminal tail 24-46
Fig. 6. Netrin cyclic peptide 1 binds APP similarly to netrin-1. A) The molecular model of netrin-1 is shown, and below it, cyclic peptide 1
(orange). B) The sensogram obtained for peptide 1 for 0–16.7 M peptide pumped at 20 l per min in 1% DMSO, 20 mM sodium phosphate
pH 6.7, 125 mM sodium chloride, 0.05% Tween 20 over a flow channel containing TRX-eAPP575-624 (20 kDa) is shown. C) Comparison
of the response at equilibrium for the all three peptides is shown. The response for peptides 2 and 3 was not significantly different from the
flow channel containing TRX for concentrations below 10 M. The responses were calculated and graphed as described in the Methods. The
KD was estimated to be 0.018 M with a 95% confidence interval of 0.015–0.021 M for peptide 1. The calculated KDs for peptides 2 and 3
were greater than 10 M, consistent with a lack of signal for these peptides in the concentration range used for the experiment. R2 was greater
than 0.95 for peptide 1. D) The sensogram obtained for peptide 1 pumped at 20 l per min in 1% DMSO, 20 mM sodium phosphate pH 7.4,
125 mM sodium chloride, 0.05% Tween 20 for concentrations 0–14 M is shown. E) The response at equilibrium for peptide 1 at pH 7.4 is
shown. The calculated KD was estimated to be approximately 30 M, which suggests that the binding of peptide 1 to TRX-eAPP575-624 is
pH dependent.
pumpedat20 lperminin1%DMSO,20 mMsodium
phosphate pH 7.4, 125 mM sodium chloride, 0.05%
Tween 20 for concentrations 0–14 M is shown in
Fig. 6D, and the response at equilibrium for peptide
1 at pH 7.4 is shown in Fig. 6E. The calculated KD
wasestimatedtobeapproximately30 M,whichsug-
gests that peptide 1 does indeed bind APP and that
this binding to TRX-eAPP575-624 is pH dependent.
Thus,peptide1sharesseveralkeycharacteristicswith
netrin-1 and is undergoing further study.
DISCUSSION
The relationship of self-amplification mechanisms
to normal physiology is incompletely defined, but
it is possible that the phenomenon may be a gen-
eral one that is related to molecular switching that
requires positive feedback loops and may be medi-
ated by multiple mechanisms, both conformational
and otherwise. In the case of A, amplification
involves, in part, protease inhibition and competition
for ␣-secretase cleavage; however, this does not pre-
clude the possibility that other mechanisms are also
involved, such as effects on transcription or post-
translational modifications such as phosphorylation.
Multiple neurodegenerative diseases display fea-
turessuchaspositivefeedbackloopsinwhichspecific
proteins or peptide fragments trigger increased
production of themselves leading to imbalanced sig-
naling. In addition, certain protein isoforms have
a propensity to aggregate and to induce further
aggregation when transmitted cell-to-cell, and as
these aggregates often have a longer half-life, this
results in increases of the protein in tissue. In recent
publications, tauopathies [35–37], synucleinopathies
[38–40], and amyotrophic lateral sclerosis [41, 42]
have all been shown to involve proteins (phosphory-
lated tau, ␣-synuclein, and SOD1, respectively) that
apparently self-amplify by either de novo increases in
production or propagation of aggregates. The mech-
anisms underlying these phenomena may differ, but
elucidating each of them is of great importance to
understanding disease progression and treatment, and
it is likely that many similarities will be found.](https://image.slidesharecdn.com/9d4f4bed-5158-4741-be4b-0954b89c28d3-161106220247/85/Spilman-Corset-Netrin-paper-15-320.jpg)
![238 P.R. Spilman et al. / Netrin-1 Disrupts Aβ Amplification
Fig. 7. A and netrin-1 effects on APP processing. A) In the trophic state, ␣-secretase ADAM10 cleaves full-length (FL) APP at the
plasma membrane, generating sAPP␣ and ␣CTF. B) Alternatively, -secretase BACE1 can bind FL APP forming a dimer that is then
endocytosed to an acidic compartment (C) wherein BACE cleavage occurs, producing sAPP and CTF. CTF can then be cleaved by
␥ secretase (D) to generate A and the APP intracellular domain (AICD). The exact compartment wherein ␥ cleavage occurs may vary.
E) As A contains the ADAM10 cleavage site (green line), it may inhibit ADAM10 cleavage of FL APP by competing for cleavage.
Alternatively, it may interact with FL APP at the A cognate region and block ADAM10 cleavage (not shown). F) Conversely, netrin-1 can
bind the A cognate region of FL APP between the ADAM10 and BACE cleavage sites (purple line), and this may block dimer formation,
endocytosis and ultimately cleavage by BACE1. Thus A interaction decreases sAPP␣ production and leaves more substrate available for
BACE cleavage, resulting in increased A production; and netrin-1 opposes this action by preventing BACE cleavage, leaving more substrate
for ADAM10 cleavage and sAPP␣ production. This schematic does not preclude other effects, such as a direct interaction between netrin-1
and A leading to sequestration of A (F, insert).
Inmanyprotein-basedneurodegenerativediseases,
increased local protein concentration contributes to
protein oligomerization and aggregation resulting
in ‘seeding’ and amplification of amyloid forma-
tion that is age-dependent [43]. Then, from these
areas of increased protein concentration, proteins
with the ability to self-amplify spread, following neu-
roanatomical pathways and networks in addition to
fluiddrainagechannelsandthevasculartransportsys-
tem [44–46]. AD fits this model, as A pathologies
appear first where protein expression is the highest
and then spread from these areas [47].
Additional similarities in mechanisms underlying
pathological protein amplification include the finding
that small, soluble A species (oligomers) are partic-
ularly potent inducers of -amyloidosis [48], similar
to the strong infectivity of small, non-fibrillar prion
particles[49].Proteinisoformand/oroligomericstate
are likely key to A and other proteins’ ability to
self-amplify or to persist due to reduced turnover.
To confirm which state is the most like to induce
increases or persistence of the protein, experimental
paradigms outlined here can be used for in vitro anal-
ysis, while those outlined in Stohr [7] and Hamaguchi
[45] for in vivo analysis.
Biological mechanisms disrupting A amplifica-
tion are of therapeutic interest. In Tian et al. [50],
it was shown that ␣CTF, a product (in addition
to sAPP␣) of ADAM10 cleavage of FL APP
(Fig. 7A) can inhibit ␥-secretase. ␣CTF comprises
the ␥-cleavage site but is described as a relatively poor
substrate for ␥ secretase, yet nonetheless interacts
with it and decreases cleavage of CTF. In addi-
tion, sAPP␣, which contains an intact -site, has
been shown by us and others to interact with BACE1
[51, 52] and reduce BACE1 cleavage of FL APP.](https://image.slidesharecdn.com/9d4f4bed-5158-4741-be4b-0954b89c28d3-161106220247/85/Spilman-Corset-Netrin-paper-16-320.jpg)
![P.R. Spilman et al. / Netrin-1 Disrupts Aβ Amplification 239
What these studies and those presented here reveal
is what may be a common theme to the trophic or
anti-trophic state of APP cleavage: cleavage prod-
ucts themselves modulate the pathways for cleavage
by either inhibiting or competing for cleavage. As
seen in Fig. 7B, in an anti-trophic state, FL APP
(or a FL APP dimer) interacts with BACE1 on the
cell surface and is transported to an acidic endoso-
mal compartment (Fig. 7C) wherein it is cleaved by
BACE1, generating sAPP and CTF. This is fol-
lowed by ␥-secretase cleavage, producing A and
AICD (Fig. 7D). We show here that A, or A
oligomers, can inhibit and/or compete for ADAM10
cleavage of FL APP (Fig. 7E); thereby increas-
ing FL APP substrate available for interaction with
BACE1, ultimately amplifying A production.
The triggers of an initial increase in A production
that may ultimately lead to amplification are many.
As stated above, aggregation, seeding or “nucleation”
areage-dependent,butasnotallagedpersonsdevelop
neurodegenerative disease, other factors, both envi-
ronmental and genetic, are likely to be involved. In
AD, the most common genetic risk factor is the pres-
ence of the ApoE 4 allele. Possession of the 4 allele
has been shown to increase A production in part by
increasing endocytosis of BACE1-bound FL APP
substrate [53]. Other risk factors associated with A
increases include diabetes [54], brain injury [55], and
hypoxia [56], just to name a few. The mechanism(s)
by which these risk factors induce an increase in A
production may vary, but they are likely to share the
effect of shortening the time necessary to reach a level
of A production that triggers self-amplification.
Much about the mechanism of A amplification
is revealed here by the ability of netrin-1, to some
extent, to interrupt it, making this not only useful
in elucidating the mechanism, but providing further
evidence of netrin-1’s potential as a possible AD ther-
apeutic. Netrin-1 interacts with FL APP at the A
cognate region near the BACE1 cleavage site [14]
(Fig. 7F) and therefore may either prevent BACE1-
APP complex formation or inhibit cleavage in the
endosomal compartment (not shown). It could also
be that netrin-1, known to bind A itself, sequesters
A and prevents interaction with ADAM10 (Fig. 7F
insert). In this/these way(s), netrin-1 may disrupt A
amplification.
Netrin-1 has a variety of other effects that are likely
to be of benefit in AD. The best characterized is
netrin-1’s role as an axonal guidance factor, which it
manifests by interacting with its receptors, deleted in
colorectal cancer (DCC) and uncoordinated gene 5H
(UNC5H). When unbound by netrin-1, these recep-
tors can induce caspase activation and apoptosis.
Of great interest here, it has recently been shown
that a mutation of the netrin receptor gene UNC5C
increases risk for late-onset AD and was found in
vitro to increase cell death, particularly in response
to A and glutamate [57].
In addition, with trophic deprivation, APP can
undergo BACE1 cleavage to generate sAPP and
further N-terminal cleavage generating N-APP,
which interacts with death receptor 6 (DR6), ulti-
mately leading to loss of axons [58], and netrin-1
may provide a trophic factor that interrupts this pro-
cess. Further studies need to be performed to clarify
this possibility. Other effects of netrin-1—seen when
it is overexpressed in a rat stroke model—include an
increase in neurogenesis [59] and cerebral vascular-
ization [60]. Netrin-1 has also been shown to increase
dendritic arborization and complexity, as well as
synapse formation and adhesion, by reorganizing
cytoskeletal structures through Src family kinase
signaling and mTOR-dependent protein translation
[61]. A role for netrin-1 in local protein synthesis
was recently confirmed in Kim et al. [62], where
it was shown that netrin-1/DCC signaling leads to
translation of mRNA translocated to synapses, a pro-
cess necessary for experience-dependent plasticity.
Again, APP may be important in these netrin-
mediated processes as it has been recently shown that
APP is part of a complex with DCC acting as a co-
receptor, enhancing axonal guidance and other effects
[21].
Netrin-1’s anti-A amplification and other anti-
AD effects make it a promising AD therapeutic.
Furthermore, it may potentially be of utility in
the treatment of traumatic brain injury (TBI), not
only as it may abrogate A amplification in TBI
[63], but also as it may restore blood-brain barrier
integrity by increasing expression of tight junction-
associated proteins [64]. Here, to further investigate
its therapeutic potential, we induced netrin-1 expres-
sion in transgenic mice also carrying APP with
familial AD mutations, and saw an increase in the
sAPP␣/A1-42 ratio. We also delivered netrin-1
ICV to AD model mice to confirm its efficacy. In
parallel, we designed, synthesized, and performed
in vitro analysis of netrin-1 mimetic cyclic peptides,
as they offer a more readily deliverable option. In
our ongoing efforts to develop netrin mimetics with
greater potency than mimetic peptide one, we will
continue the optimization of cyclic peptide 1 and
screening of our chemical library to identify netrin-1](https://image.slidesharecdn.com/9d4f4bed-5158-4741-be4b-0954b89c28d3-161106220247/85/Spilman-Corset-Netrin-paper-17-320.jpg)
![240 P.R. Spilman et al. / Netrin-1 Disrupts Aβ Amplification
mimetics that increase sAPP␣ and/or decrease
BACE1 cleavage and A production. The establish-
ment of the CED method for delivery of a netrin
construct described here will allow us to ascertain
effects in a variety of AD and other neurodegenera-
tive disease models, and can also be used to deliver
protein, peptides, mimetics, and compounds that have
poor brain penetrance.
The development of a protein or peptide as an AD
therapeutic has many challenges that are arguably
greater than those faced in development of a chemical
therapeutic. Other (non-antibody) biologic potential
AD therapies include the tumor necrosis factor ␣
(TNF␣) inhibitor etanercept, nerve growth factor,
brain-derived neurotrophic factor, and insulin-like
growth factor 2 [65–68], some of which have been
taken as far as pilot clinical trials using gene therapy;
this is one of our future goals for netrin-1. Here, using
a variety of approaches, we have revealed some of
netrin-1’s promise as an AD therapeutic and provided
further support for targeting ADAM10 and sAPP␣
enhancement in AD.
ACKNOWLEDGMENTS
We would like to thank Rowena Abulencia for help
with the manuscript, and Harris Spilman, Jakob Dorf-
man, Tina Bilousova, Olivier Descamps and Jeremy
Lambert for technical assistance.
Dr. Bredesen/The Buck Institute holds a patent for
the use of netrin-1 in Alzheimer’s disease: US Patent
No. 8,329,653.
This work was supported by the NIH (AG12282 to
D.E.B.), the Douglas and Ellen Rosenberg Founda-
tion, the Joseph Drown Foundation, and BioMarin,
Inc.
Authors’ disclosures available online (http://j-alz.
com/manuscript-disclosures/15-1046r2).
SUPPLEMENTARY MATERIAL
The supplementary material is available in the
electronic version of this article: http://dx.doi.org/
10.3233/JAD-151046.
REFERENCES
[1] Ikeda S, Wong CW, Allsop D, Landon M, Kidd M, Glenner
GG (1987) Immunogold labeling of cerebrovascular and
neuritic plaque amyloid fibrils in Alzheimer’s disease with
an anti-beta protein monoclonal antibody. Lab Invest 57,
446-449.
[2] Hardy J, Allsop D (1991) Amyloid deposition as the cen-
tral event in the aetiology of Alzheimer’s disease. Trends
Pharmacol Sci 12, 383-388.
[3] GoedertM(1993)Tauproteinandtheneurofibrillarypathol-
ogy of Alzheimer’s disease. Trends Neurosci 16, 460-465.
[4] Dickson D (1997) Discovery of new lesions in neurode-
generative diseases with monoclonal antibody techniques:
Is there a non-amyloid precursor to senile plaques? Am J
Pathol 151, 7-11.
[5] YinRH,TanL,JiangT,YuJT(2014)Prion-likemechanisms
in Alzheimer’s disease. Curr Alzheimer Res 11, 755-764.
[6] Yang AJ, Chandswangbhuvana D, Shu T, Henschen A,
Glabe CG (1999) Intracellular accumulation of insoluble,
newly synthesized abetan-42 in amyloid precursor protein-
transfected cells that have been treated with Abeta1-42. J
Biol Chem 274, 20650-20656.
[7] Stohr J, Watts JC, Mensinger ZL, Oehler A, Grillo SK,
DeArmond SJ, Prusiner SB, Giles K (2012) Purified and
synthetic Alzheimer’s amyloid beta (Abeta) prions. Proc
Natl Acad Sci U S A 109, 11025-11030.
[8] Eisele YS, Bolmont T, Heikenwalder M, Langer F, Jacobson
LH, Yan ZX, Roth K, Aguzzi A, Staufenbiel M, Walker LC,
Jucker M (2009) Induction of cerebral beta-amyloidosis:
Intracerebral versus systemic Abeta inoculation. Proc Natl
Acad Sci U S A 106, 12926-12931.
[9] Selkoe DJ, Yamazaki T, Citron M, Podlisny MB, Koo EH,
Teplow DB, Haass C (1996) The role of APP processing
and trafficking pathways in the formation of amyloid beta-
protein. Ann N Y Acad Sci 777, 57-64.
[10] Xia W, Zhang J, Ostaszewski BL, Kimberly WT, Seubert
P, Koo EH, Shen J, Selkoe DJ (1998) Presenilin 1 regulates
the processing of beta-amyloid precursor protein C-terminal
fragments and the generation of amyloid beta-protein in
endoplasmic reticulum and Golgi. Biochemistry 37, 16465-
16471.
[11] Vassar R, Bennett BD, Babu-Khan S, Kahn S, Mendiaz EA,
Denis P, Teplow DB, Ross S, Amarante P, Loeloff R, Luo
Y, Fisher S, Fuller J, Edenson S, Lile J, Jarosinski MA,
Biere AL, Curran E, Burgess T, Louis JC, Collins F, Tre-
anor J, Rogers G, Citron M (1999) Beta-secretase cleavage
of Alzheimer’s amyloid precursor protein by the transmem-
brane aspartic protease BACE. Science 286, 735-741.
[12] Bredesen DE (2009) Neurodegeneration in Alzheimer’s dis-
ease: Caspases and synaptic element interdependence. Mol
Neurodegener 4, 27.
[13] Bertrand E, Brouillet E, Caille I, Bouillot C, Cole GM,
Prochiantz A, Allinquant B (2001) A short cytoplasmic
domain of the amyloid precursor protein induces apoptosis
in vitro and in vivo. Mol Cell Neurosci 18, 503-511.
[14] Lourenco FC, Galvan V, Fombonne J, Corset V, Llambi F,
Muller U, Bredesen DE, Mehlen P (2009) Netrin-1 interacts
with amyloid precursor protein and regulates amyloid-beta
production. Cell Death Differ 16, 655-663.
[15] Lu DC, Rabizadeh S, Chandra S, Shayya RF, Ellerby LM,
Ye X, Salvesen GS, Koo EH, Bredesen DE (2000) A second
cytotoxic proteolytic peptide derived from amyloid beta-
protein precursor. Nat Med 6, 397-404.
[16] Park SA, Shaked GM, Bredesen DE, Koo EH (2009) Mech-
anism of cytotoxicity mediated by the C31 fragment of the
amyloid precursor protein. Biochem Biophys Res Commun
388, 450-455.
[17] Barger SWMM (1996) Induction of neuroprotective kappa-
B-dependent transcription by secreted forms of the
Alzheimer’s (-amyloid precursor. Mol Brain Res 40, 116-
126.](https://image.slidesharecdn.com/9d4f4bed-5158-4741-be4b-0954b89c28d3-161106220247/85/Spilman-Corset-Netrin-paper-18-320.jpg)
![P.R. Spilman et al. / Netrin-1 Disrupts Aβ Amplification 241
[18] Shaked GM, Kummer MP, Lu DC, Galvan V, Bredesen DE,
Koo EH (2006) Abeta induces cell death by direct inter-
action with its cognate extracellular domain on APP (APP
597-624). FASEB J 20, 1254-1256.
[19] Libeu CP, Poksay KS, John V, Bredesen DE (2011) Struc-
tural and functional alterations in amyloid-beta precursor
protein induced by amyloid-beta peptides. J Alzheimers Dis
25, 547-566.
[20] Lu DC, Shaked GM, Masliah E, Bredesen DE, Koo EH
(2003) Amyloid beta protein toxicity mediated by the for-
mation of amyloid-beta protein precursor complexes. Ann
Neurol 54, 781-789.
[21] Rama N, Goldschneider D, Corset V, Lambert J, Pays
L, Mehlen P (2012) Amyloid precursor protein regulates
netrin-1-mediated commissural axon outgrowth. J Biol
Chem 287, 30014-30023.
[22] Steinacker P, Kuhle FL, Petzold J, Tumani A, Ludolph H,
Otto AC, Brettschneider MJ (2011) Soluble beta-amyloid
precursor protein is related to disease progression in amy-
otrophic lateral sclerosis. PLoS One 6, e23600.
[23] Keino-Masu K, Masu M, Hinck L, Leonardo ED, Chan SS,
Culotti JG, Tessier-Lavigne M (1996) Deleted in colorectal
cancer (DCC) encodes a netrin receptor. Cell 87, 175-185.
[24] Eichler JHR (1997) Synthesis of cyclic disulfide peptides:
Comparison of oxidation methods. Protein Pept Lett 4, 157-
164.
[25] Mucke L, Yu ME, Mallory GQ, Rockenstein M, Tatsuno
EM, Hu G, Kholodenko K, Johnson-Wood D, McConlogue
KL (2000) High-level neuronal expression of abeta 1-42
in wild-type human amyloid protein precursor transgenic
mice: Synaptotoxicity without plaque formation. J Neurosci
20, 4050-4058.
[26] Sinha S, Anderson JP, Barbour R, Basi GS, Caccavello R,
Davis D, Doan M, Dovey HF, Frigon N, Hong J, Jacobson-
Croak K, Jewett N, Keim P, Knops J, Lieberburg I, Power M,
TanH,TatsunoG,TungJ,SchenkD,SeubertP,Suomensaari
SM, Wang S, Walker D, Zhao J, McConlogue L, John V
(1999)Purificationandcloningofamyloidprecursorprotein
beta-secretase from human brain. Nature 402, 537-540.
[27] Utomo AR, Nikitin AY, Lee WH (1999) Temporal, spatial,
and cell type-specific control of Cre-mediated DNA recom-
bination in transgenic mice. Nat Biotechnol 17, 1091-1096.
[28] Hsia AY, Masliah E, McConlogue L, Yu GQ, Tatsuno G,
Hu K, Kholodenko D, Malenka RC, Nicoll RA, Mucke L
(1999) Plaque-independent disruption of neural circuits in
Alzheimer’s disease mouse models. Proc Natl Acad Sci U
S A 96, 3228-3233.
[29] Bevins RA, Besheer J (2006) Object recognition in rats and
mice: A one-trial non-matching-to-sample learning task to
study ‘recognition memory’. Nat Protoc 1, 1306-1311.
[30] Matsushita T, Elliger S, Elliger C, Podsakoff G, Villarreal L,
Kurtzman GJ, Iwaki Y, Colosi P (1998) Adeno-associated
virus vectors can be efficiently produced without helper
virus. Gene Ther 5, 938-945.
[31] Wright JF, Qu G, Tang C, Sommer JM (2003) Recombinant
adeno-associated virus: Formulation challenges and strate-
gies for a gene therapy vector. Curr Opin Drug Discov Devel
6, 174-178.
[32] Barua NU, Woolley M, Bienemann AS, Johnson D, Wyatt
MJ, Irving C, Lewis O, Castrique E, Gill SS (2013)
Convection-enhanced delivery of AAV2 in white matter–a
novelmethodforgenedeliverytocerebralcortex.JNeurosci
Methods 220, 1-8.
[33] Rosenbluth KH, Luz M, Mohr E, Mittermeyer S, Bringas
J, Bankiewicz KS (2011) Design of an in-dwelling cannula
for convection-enhanced delivery. J Neurosci Methods 196,
118-123.
[34] Lonser RR, Sarntinoranont M, Morrison PF, Oldfield EH
(2015) Convection-enhanced delivery to the central nervous
system. J Neurosurg 122, 697-706.
[35] Clavaguera F, Bolmont T, Crowther RA, Abramowski D,
Frank S, Probst A, Fraser G, Stalder AK, Beibel M,
Staufenbiel M, Jucker M, Goedert M, Tolnay M (2009)
Transmission and spreading of tauopathy in transgenic
mouse brain. Nat Cell Biol 11, 909-913.
[36] Frost B, Jacks RL, Diamond MI (2009) Propagation of tau
misfolding from the outside to the inside of a cell. J Biol
Chem 284, 12845-12852.
[37] Guo JL, Lee VM (2011) Seeding of normal Tau by patholog-
ical Tau conformers drives pathogenesis of Alzheimer-like
tangles. J Biol Chem 286, 15317-15331.
[38] Mougenot AL, Nicot S, Bencsik A, Morignat E, Verchere J,
Lakhdar L, Legastelois S, Baron T (2012) Prion-like accel-
eration of a synucleinopathy in a transgenic mouse model.
Neurobiol Aging 33, 2225-2228.
[39] Volpicelli-Daley LA, Luk KC, Patel TP, Tanik SA,
Riddle DM, Stieber A, Meaney DF, Trojanowski JQ, Lee
VM (2011) Exogenous alpha-synuclein fibrils induce Lewy
body pathology leading to synaptic dysfunction and neuron
death. Neuron 72, 57-71.
[40] Narkiewicz J, Giachin G, Legname G (2014) in vitro aggre-
gation assays for the characterization of alpha-synuclein
prion-like properties. Prion 8, 19-32.
[41] Munch C, O’Brien J, Bertolotti A (2011) Prion-like
propagation of mutant superoxide dismutase-1 misfold-
ing in neuronal cells. Proc Natl Acad Sci U S A 108,
3548-3553.
[42] Grad LI, Cashman NR (2014) Prion-like activity of Cu/Zn
superoxide dismutase: Implications for amyotrophic lateral
sclerosis. Prion 8, 33-41.
[43] Liberski PP (2014) Prion, prionoids and infectious amyloid.
Parkinsonism Relat Disord 20(Suppl 1), S80-S84.
[44] Frost B, Diamond MI (2010) Prion-like mechanisms in neu-
rodegenerative diseases. Nat Rev Neurosci 11, 155-159.
[45] Hamaguchi T, Eisele YS, Varvel NH, Lamb BT, Walker LC,
Jucker M (2012) The presence of Abeta seeds, and not age
per se, is critical to the initiation of Abeta deposition in the
brain. Acta Neuropathol 123, 31-37.
[46] Domert J, Rao SB, Agholme L, Brorsson AC, Marcusson
J, Hallbeck M, Nath S (2014) Spreading of amyloid-beta
peptides via neuritic cell-to-cell transfer is dependent on
insufficient cellular clearance. Neurobiol Dis 65, 82-92.
[47] Harris JA, Devidze N, Verret L, Ho K, Halabisky B,
Thwin MT, Kim D, Hamto P, Lo I, Yu GQ, Palop JJ,
Masliah E, Mucke L (2010) Transsynaptic progression
of amyloid-beta-induced neuronal dysfunction within the
entorhinal-hippocampal network. Neuron 68, 428-441.
[48] LangerF,EiseleYS,FritschiSK,StaufenbielM,WalkerLC,
Jucker M (2011) Soluble Abeta seeds are potent inducers
of cerebral beta-amyloid deposition. J Neurosci 31, 14488-
14495.
[49] Silveira JR, Raymond GJ, Hughson AG, Race RE, Sim VL,
Hayes SF, Caughey B (2005) The most infectious prion
protein particles. Nature 437, 257-261.
[50] Tian Y, Crump CJ, Li YM (2010) Dual role of alpha-
secretase cleavage in the regulation of gamma-secretase
activity for amyloid production. J Biol Chem 285, 32549-
32556.
[51] Peters-Libeu C, Campagna J, Mitsumori M, Poksay KS,
Spilman P, Sabogal A, Bredesen DE, John V (2015)](https://image.slidesharecdn.com/9d4f4bed-5158-4741-be4b-0954b89c28d3-161106220247/85/Spilman-Corset-Netrin-paper-19-320.jpg)
![242 P.R. Spilman et al. / Netrin-1 Disrupts Aβ Amplification
sAbetaPPalpha is a potent endogenous inhibitor of BACE1.
J Alzheimers Dis 47, 545-555.
[52] Obregon D, Hou H, Deng J, Giunta B, Tian J, Darlington
D, Shahaduzzaman M, Zhu Y, Mori T, Mattson MP, Tan
J (2012) Soluble amyloid precursor protein-alpha modu-
lates beta-secretase activity and amyloid-beta generation.
Nat Commun 3, 777.
[53] He X, Cooley K, Chung CH, Dashti N, Tang J (2007)
Apolipoprotein receptor 2 and X11 alpha/beta mediate
apolipoprotein E-induced endocytosis of amyloid-beta pre-
cursor protein and beta-secretase, leading to amyloid-beta
production. J Neurosci 27, 4052-4060.
[54] de la Monte SM, Tong M (2014) Brain metabolic dysfunc-
tion at the core of Alzheimer’s disease. Biochem Pharmacol
88, 548-559.
[55] Sivanandam TM, Thakur MK (2012) Traumatic brain
injury: A risk factor for Alzheimer’s disease. Neurosci
Biobehav Rev 36, 1376-1381.
[56] Popa-Wagner A, Buga AM, Popescu B, Muresanu D (2015)
Vascular cognitive impairment, dementia, aging and energy
demand. A vicious cycle. J Neural Transm 122(Suppl 1),
S47-S54.
[57] Wetzel-Smith MK, Hunkapiller J, Bhangale TR, Srinivasan
K, Maloney JA, Atwal JK, Sa SM, Yaylaoglu MB, Foreman
O, Ortmann W, Rathore N, Hansen DV, Tessier-Lavigne M,
Mayeux R, Pericak-Vance M, Haines J, Farrer LA, Schel-
lenberg GD, Goate A, Behrens TW, Cruchaga C, Watts RJ,
Graham RR (2014) A rare mutation in UNC5C predisposes
tolate-onsetAlzheimer’sdiseaseandincreasesneuronalcell
death. Nat Med 20, 1452-1457.
[58] Nikolaev A, McLaughlin T, O’Leary DD, Tessier-Lavigne
M (2009) APP binds DR6 to trigger axon pruning and neu-
ron death via distinct caspases. Nature 457, 981-989.
[59] Liao SJ, Gong Q, Chen XR, Ye LX, Ding Q, Zeng JS, Yu
J (2013) Netrin-1 rescues neuron loss by attenuating sec-
ondary apoptosis in ipsilateral thalamic nucleus following
focal cerebral infarction in hypertensive rats. Neuroscience
231, 225-232.
[60] Fan Y, Shen F, Chen Y, Hao Q, Liu W, Su H, Young WL,
Yang GY (2008) Overexpression of netrin-1 induces neo-
vascularizationintheadultmousebrain.JCerebBloodFlow
Metab 28, 1543-1551.
[61] Goldman JS, Ashour MA, Magdesian MH, Tritsch NX,
Harris SN, Christofi N, Chemali R, Stern YE, Thompson-
Steckel G, Gris P, Glasgow SD, Grutter P, Bouchard JF,
Ruthazer ES, Stellwagen D, Kennedy TE (2013) Netrin-
1 promotes excitatory synaptogenesis between cortical
neurons by initiating synapse assembly. J Neurosci 33,
17278-17289.
[62] Kim S, Martin KC (2015) Neuron-wide RNA transport
combines with netrin-mediated local translation to spatially
regulate the synaptic proteome. Elife 4, e04158.
[63] Marklund N, Farrokhnia N, Hanell A, Vanmechelen E,
Enblad P, Zetterberg H, Blennow K, Hillered L (2014) Mon-
itoring of beta-amyloid dynamics after human traumatic
brain injury. J Neurotrauma 31, 42-55.
[64] Wen J, Qian S, Yang Q, Deng L, Mo Y, Yu Y (2014)
Overexpression of netrin-1 increases the expression of tight
junction-associated proteins, claudin-5, occludin, and ZO-
1, following traumatic brain injury in rats. Exp Ther Med 8,
881-886.
[65] Mellott TJ, Pender SM, Burke RM, Langley EA, Blusz-
tajn JK (2014) IGF2 ameliorates amyloidosis, increases
cholinergic marker expression and raises BMP9 and neu-
rotrophin levels in the hippocampus of the APPswePS1dE9
Alzheimer’s disease model mice. PLoS One 9, e94287.
[66] Cheng X, Shen Y, Li R (2014) Targeting TNF: A therapeutic
strategy for Alzheimer’s disease. Drug Discov Today 19,
1822-1827.
[67] Nagahara AH, Merrill DA, Coppola G, Tsukada S,
Schroeder BE, Shaked GM, Wang L, Blesch A, Kim A,
Conner JM, Rockenstein E, Chao MV, Koo EH, Geschwind
D, Masliah E, Chiba AA, Tuszynski MH (2009) Neuropro-
tective effects of brain-derived neurotrophic factor in rodent
and primate models of Alzheimer’s disease. Nat Med 15,
331-337.
[68] Tuszynski MH, Yang JH, Barba D, U HS, Bakay RA, Pay
MM, Masliah E, Conner JM, Kobalka P, Roy S, Nagahara
AH (2015) Nerve growth factor gene therapy: Activation of
neuronal responses in Alzheimer disease. JAMA Neurol 72,
1139-1147.](https://image.slidesharecdn.com/9d4f4bed-5158-4741-be4b-0954b89c28d3-161106220247/85/Spilman-Corset-Netrin-paper-20-320.jpg)

































































