What are smallRNAs?
•Small RNAs are a pool of 21 to 24 nt
RNAs that generally function in gene
silencing
•Small RNAs contribute to post-
transcriptional gene silencing by
affecting mRNA translation or stability
•Small RNAs contribute to
transcriptional gene silencing through
epigenetic modifications to chromatin
AAAAA
RNA Pol
Histone modification, DNA methylation
3.
• MicroRNAs
• siRNAs
•Ta-siRNAs (trans acting siRNAs)
• Nat-siRNAs (natural antisense transcript-derived small interfering RNAs)
• heterochromatic siRNAs (hc-siRNAs)
• Long small interfering RNAs (l-siRNAs)
Repertoire of Small RNAs
J. Couzin, "Small RNAs Make Big Splash", Science 298:2296-
7, 20 December2002.
2002's Breakthrough of the Year by SCIENCE
4.
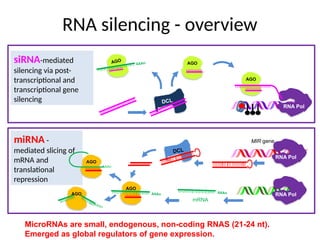
RNA silencing -overview
DCL
AGO
AGO
RNA Pol
AGO
siRNA-mediated
silencing via post-
transcriptional and
transcriptional gene
silencing
AAAn
DCL
MIR gene
RNA Pol
AGO
RNA Pol
miRNA -
mediated slicing of
mRNA and
translational
repression
mRNA
AGO
AGO
AAAn
AAAn
AAAn
AAAn
MicroRNAs are small, endogenous, non-coding RNAS (21-24 nt).
Emerged as global regulators of gene expression.
5.
• MicroRNAs (miRNAs)are genomically encoded non-coding RNAs that help
regulate gene expression, particularly during development. The phenomenon of
RNA interference, broadly defined, includes the endogenously induced gene
silencing effects of miRNAs as well as silencing triggered by foreign dsRNA.
• MicroRNAs (miRNAs) are an abundant class of ~20-24 nucleotide noncoding
RNAs with important roles in the regulation of gene expression, of protein coding
genes, in plants and animals.
• miRNAS are often found to be conserved evolutionarily across species
boundaries.
• miRNAs are known to silence genes posttranscriptionally by targeting cognate
mRNAs for degradation or translation repression.
• The total no. of miRNAs in each organism is unknown but is estimated to represent
~ 1% of the no. of coding genes
What are miRNAs
6.
microRNAs - miRNAS
•miRNAs are thought to have evolved from siRNAs, and are
produced and processed somewhat similarly
• Plants have a small number of highly conserved miRNAs, and
a large number of non-conserved miRNAs
• miRNAs are encoded by specific MIR genes but act on other
genes – they are trans-acting regulatory factors
• miRNAs in plants regulate developmental and physiological
events
7.
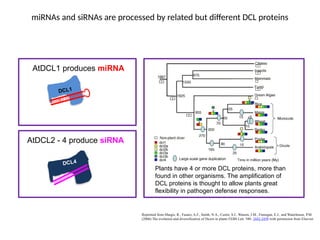
miRNAs and siRNAsare processed by related but different DCL proteins
Reprinted from Margis, R., Fusaro, A.F., Smith, N.A., Curtin, S.J., Watson, J.M., Finnegan, E.J., and Waterhouse, P.M.
(2006) The evolution and diversification of Dicers in plants FEBS Lett. 580: 2442-2450 with permission from Elsevier.
Plants have 4 or more DCL proteins, more than
found in other organisms. The amplification of
DCL proteins is thought to allow plants great
flexibility in pathogen defense responses.
AtDCL1 produces miRNA
AtDCL2 - 4 produce siRNA
DCL4
DCL1
8.
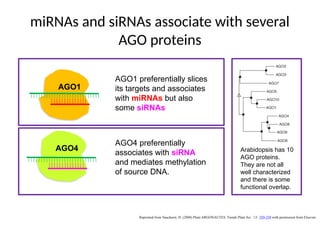
miRNAs and siRNAsassociate with several
AGO proteins
AGO1
AGO4
AGO1 preferentially slices
its targets and associates
with miRNAs but also
some siRNAs
AGO4 preferentially
associates with siRNA
and mediates methylation
of source DNA.
Arabidopsis has 10
AGO proteins.
They are not all
well characterized
and there is some
functional overlap.
Reprinted from Vaucheret, H. (2008) Plant ARGONAUTES. Trends Plant Sci. 13: 350-358 with permission from Elsevier.
9.
Differences between miRNAand siRNA
• miRNAs are processed from single RNA molecules that include an
imperfect stem loop secondary structure whereas siRNAs are
processed from long double stranded RNAs
• miRs are endogenously encoded small noncoding RNAs while
siRNAs are often of exogenous origin
• Many siRNAs target the gene from which they are derived or very
closely related genes. In contrast miRNAs regulate genes unrelated
to the loci encoding miRNAs.
• Many Arabidopsis siRNAs require RDRp for their biogenesis,
whereas miRNAs do not.
• Most Arab. miRNAs are processed by DCL1 whereas siRNA require
DCL3 or DCL4
10.
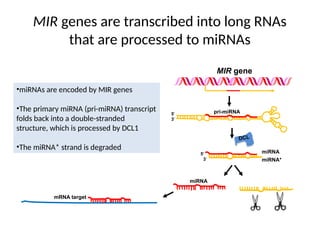
MIR genes aretranscribed into long RNAs
that are processed to miRNAs
•miRNAs are encoded by MIR genes
•The primary miRNA (pri-miRNA) transcript
folds back into a double-stranded
structure, which is processed by DCL1
•The miRNA* strand is degraded
DCL
3'
5' miRNA
miRNA*
3'
5' pri-miRNA
miRNA
MIR gene
mRNA target
11.
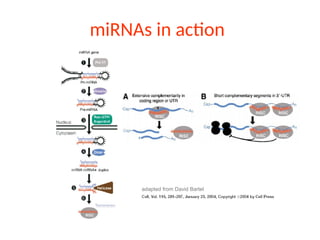
Diverse responses beingregulated by microRNAs
All plant miRNAs are highly complementary to target mRNAs, which the miRNA repress through direct
cleavage. Majority of known plant miRNAs targets encode transcription factors or other regulatory
proteins
12.
MicroRNAs
• short (20-25nt)RNA molecules
• post transcriptional gene regulation
• target mRNA for cleavage or
translational repression
• very abundant class of genes
Cell 1993; 75(5):843-854
TheC. elegans heterochronic gene lin-4 encodes small RNAs with
antisense complementarity to lin-14.
Lee RC, Feinbaum RL, Ambros V.
Nature 2000; 403(6772):901-906
The 21-nucleotide let-7 RNA regulates developmental timing in
Caenorhabditis elegans.
Reinhart BJ, Slack FJ, Basson M, Pasquinelli AE, Bettinger JC, Rougvie AE, Horvitz HR,
Ruvkun G.
The first miRNAs to be identified
15.
Genes Dev. 2002;16(13):1616-1626
MicroRNAs in plants.
Reinhart BJ, Weinstein EG, Rhoades MW, Bartel B, Bartel DP.
Whitehead Institute for Biomedical Research, and Department of Biology, Massachusetts Institute of Technology, Cambridge, Massachusetts, USA.
MicroRNAs (miRNAs) are an extensive class of ~22-nucleotide noncoding RNAs thought to
regulate gene expression in metazoans. We find that miRNAs are also present in plants,
indicating that this class of noncoding RNA arose early in eukaryotic evolution. In this paper
16 Arabidopsis miRNAs are described, many of which have differential expression patterns in
development. Eight are absolutely conserved in the rice genome. The plant miRNA loci
potentially encode stem-loop precursors similar to those processed by Dicer (a ribonuclease
III) in animals. Mutation of an Arabidopsis Dicer homolog, CARPEL FACTORY, prevents the
accumulation of miRNAs, showing that similar mechanisms direct miRNA processing in plants
and animals. The previously described roles of CARPEL FACTORY in the development of
Arabidopsis embryos, leaves, and floral meristems suggest that the miRNAs could play
regulatory roles in the development of plants as well as animals.
16.
Useful plant miRNAproperties
• (at least some) miRNAs are conserved
between species
• most precursor sequences are not conserved
• precursor structure is somewhat conserved
• mature miRNA always cut from 1 arm of RNA
helix in precursor
• plant precursors are more complex
17.
miRNA targets
• PlantmiRNAs show perfect or near-perfect
complementarity with their targets
• Computational detection of potential targets
is straightforward (Rhoades et al., 2002)
• Find these targets looking for the reverse
complement of the miRNA sequences in the
cDNAs allowing 2 to 4 mismatches at maximum
18.
Secondary structure parameters
•The miRNA should be part of a continuous helix
• The minimum number of paired residues in the
miRNA should be 15
• the maximum number of unpaired residues in both
the miRNA coding and complementary strand
should be 5.
• the maximum number of G:U pairs in the miRNA
should be 5
19.
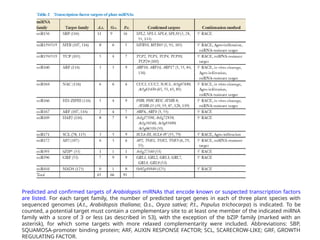
Predicted and confirmedtargets of Arabidopsis miRNAs that encode known or suspected transcription factors
are listed. For each target family, the number of predicted target genes in each of three plant species with
sequenced genomes (A.t., Arabidopsis thaliana; O.s., Oryza sativa; P.t., Populus trichocarpa) is indicated. To be
counted, a potential target must contain a complementary site to at least one member of the indicated miRNA
family with a score of 3 or less (as described in 53), with the exception of the bZIP family (marked with an
asterisk), for which some targets with more relaxed complementarity were included. Abbreviations: SBP,
SQUAMOSA-promoter binding protein; ARF, AUXIN RESPONSE FACTOR; SCL, SCARECROW-LIKE; GRF, GROWTH
REGULATING FACTOR.
A sampling ofphenotypes resulting from microRNA (miRNA) overexpression in Arabidopsis. Each panel depicts
wild type (left) compared to a corresponding specimen from a miRNA-overexpressing plant (right).
(a) miR156-overexpressing plants have increased leaf initiation and decreased apical dominance, resulting in
dramatically bushier plants.
(b) The stamens of miR164-overexpressing plants are frequently fused together rather than distinct (arrows
denote point of stamen separation).
(c) The outer floral organ whorls of miR172-overexpressing flowers are transformed into carpeloid tissue
rather than having four sepals and four petals. This phenotype is similar to that of loss-of-function mutants in
miR172 target AP2.
(d ) The jaw-D mutant phenotype, which includes severely affected leaf morphology, results from miR319
overexpression (103).
(e) Plants overexpressing miR166 have decreased stature and fertility (top) and fascinated, enlarged
inforescence stems (inset) with highly abnormal vasculature (bottom) (59).
( f ) miR159a-overexpressing plants have reduced male fertility and altered anther morphology. ( g) miR160-
overexpressing plants have disorganized root tips and fewer starch granules ( purple staining) (136).
miRNA overexpression
22.
Phenotypes of transgenicArabidopsis expressing miRNA-resistant targets. Each panel depicts a control plant,
either wild type or a plant expressing a miRNA-sensitive transgene (left), compared to a plant expressing a miRNA-
resistant transgene (right).
(a) Plants expressing miR159-resistant MYB33 have reduced stature and upwardly curled leaves (93).
(b) Plants expressing miR166-resistant PHB have radialized, reduced leaves with adaxial characteristics all around
the circumference of the leaf (87).
(c) Seedlings expressing a wild-type ARF17 transgene have two cotyledons (asterisks) and two emerging true leaves
(arrowheads), whereas miR160-resistant ARF17 seedlings have up to four cotyledons, with a leaf emerging between
each pair of cotyledons (84).
(d) Flowers expressing a wild-type CUC1 transgene have the expected four sepals (S, top) and four petals (bottom),
whereas flowers expressing a miR164-resistant CUC1 transgene often display two sepals (top) and six petals
(bottom).
(e) miR172-resistant AP2 transgenic flowers have variable numbers of foral organs; this flower has numerous
miRNA resistant target genes
23.
SUMMARY POINTS OFmicroRNAs
1.MicroRNAs are endogenously expressed, ~21-nucleotide RNAs that do not encode
proteins. A miRNA is initially expressed as a precursor RNA containing an imperfect stem-
loop, from which a miRNA/miRNA* duplex is excised by DCL1. The miRNA strand of this
duplex is subsequently incorporated into a silencing complex, where it guides targeting of
complementary RNAs.
2. Plant miRNA genes are generally not located within protein-coding genes but comprise
their own RNA polymerase II-dependent transcriptional units.
3. Plant miRNAs occur in gene families, each family contains 1–32 loci within a single
genome, each potentially encoding identical or nearly identical mature miRNAs. At least 20
miRNA families are broadly conserved in flowering plants.
24.
4. Plant miRNAsprimarily have been discovered through direct cloning and
sequencing of small cellular RNAs or through comparative genomics. Only a handful
of mutations at miRNA loci have been identified in genetic screens.
5. Nearly all plant miRNAs are highly complementary to target mRNAs, which the
miRNAs repress through directed RNA cleavage and perhaps other mechanisms.
The majority of known plant miRNA targets encode transcription factors or other
regulatory proteins, such as components of the ubiquitin and RNAi pathways.
6. Bioinformatic approaches have identified targets for nearly all plant miRNAs.
Several experimental methods have been used to confirm miRNA-target interactions
and explore the biological significance of miRNA-mediated regulation.
7. Plant miRNAs are high-level regulators of gene expression that affect numerous
aspects of plant biology, especially developmental patterning. Mutants impaired in
miRNA biogenesis exhibit severe, pleiotropic abnormalities, and plants that
overexpress particular miRNAs or express miRNA-resistant versions of particular
miRNA targets exhibit a wide array of unusual phenotypes.
25.
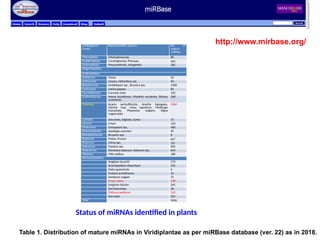
Status of miRNAsidentified in plants
http://www.mirbase.org/
Subkingdom/
Family
Representative species No. of
mature
miRNAs
Chlorophyta Chlamydomonas 86
Coniferophyta Cunninghamia, Pinussps. 669
Embryophyta Physcomitrella, Selaginella 362
Magnoliophyta
Eudictyledons
Araliaceae Panax 32
Asteraceae Cynara, Helianthus sps. 94
Brassicaceae Arabidopsis sps., Brassica sps. 1184
Caricaceae Carica papaya 81
Cucurbitaceae Cucumis melo 125
Euphorbiaceae Hevea brasiliensis, Manihot esculenta, Ricinus
communis
268
Fabaceae Acacia auriculiformis, Arachis hypogaea,
Glycine max, Lotus japonicus, Medicago
truncatula, Phaseolus vulgaris, Vigna
unguiculata
1960
Lamiales Avicennia, Digitalis, Salvia 72
Linacea Linum 124
Malvaceae Gossypium sps. 460
Ranunculaceae Aquilegia caerulea 45
Rhizophoraceae Bruuiera sps. 8
Rosaceae Malus, Prunus 667
Rutaceae Citrus sps. 261
Salicaceae Populus sps. 405
Solanaceae Nicotiana tabacum, Solanum sps. 654
Vitaceae Vitis vinifera 186
Monocotyledons
Aegilops tauschii 173
Brachypodium distachyon 525
Elaies guineensis 6
Festuca arundinacea 15
Hordeum vulgare 71
Oryza sativa 738
Sorghum bicolor 241
Saccharumsps. 36
Triticum aestivum 125
Zea mays 325
Total 9998
Table 1. Distribution of mature miRNAs in Viridiplantae as per miRBase database (ver. 22) as in 2018.
26.
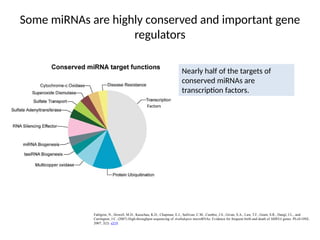
Some miRNAs arehighly conserved and important gene
regulators
Fahlgren, N., Howell, M.D., Kasschau, K.D., Chapman, E.J., Sullivan, C.M., Cumbie, J.S., Givan, S.A., Law, T.F., Grant, S.R., Dangl, J.L., and
Carrington, J.C. (2007) High-throughput sequencing of Arabidopsis microRNAs: Evidence for frequent birth and death of MIRNA genes. PLoS ONE.
2007; 2(2): e219.
Nearly half of the targets of
conserved miRNAs are
transcription factors.
Factors
27.
Targets of someconserved miRNAs
miRNA gene family Target gene family Function
156 SPL transcription factors Developmental timing
160 ARF transcription factors Auxin response,
development
165/6 HD-ZIPIII transcription
factors
Development, polarity
172 AP2 transcription factors Developmental timing, floral
organ identity
390 TAS3 (tasiRNA) which acts
on ARF transcription factors
Auxin response,
development
395 Sulfate transporter Sulfate uptake
399 Protein ubiquitination Phosphate uptake
Adapted from Willmann, M.R., and Poethig, R.S. (2007) Conservation and evolution of miRNA regulatory programs in plant development. Curr. Opin. Plant Biol. 10: 503–511..
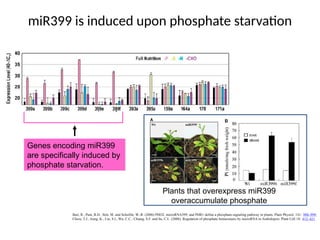
miR399 is inducedupon phosphate starvation
Bari, R., Pant, B.D., Stitt, M. and Scheible, W.-R. (2006) PHO2, microRNA399, and PHR1 define a phosphate-signaling pathway in plants. Plant Physiol. 141: 988–999;
Chiou, T.J., Aung, K., Lin, S.I., Wu, C.C., Chiang, S.F. and Su, C.L. (2006) Regulation of phosphate homeostasis by microRNA in Arabidopsis. Plant Cell 18: 412–421.
Genes encoding miR399
are specifically induced by
phosphate starvation.
Plants that overexpress miR399
overaccumulate phosphate
30.
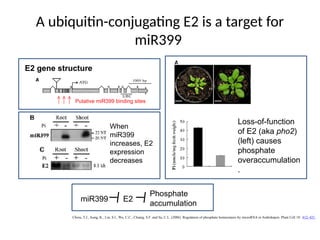
A ubiquitin-conjugating E2is a target for
miR399
E2 gene structure
Putative miR399 binding sites
When
miR399
increases, E2
expression
decreases
Loss-of-function
of E2 (aka pho2)
(left) causes
phosphate
overaccumulation
.
miR399 E2
Phosphate
accumulation
Chiou, T.J., Aung, K., Lin, S.I., Wu, C.C., Chiang, S.F. and Su, C.L. (2006) Regulation of phosphate homeostasis by microRNA in Arabidopsis. Plant Cell 18: 412–421.
31.
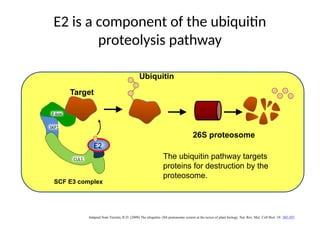
E2 is acomponent of the ubiquitin
proteolysis pathway
Adapted from Vierstra, R.D. (2009) The ubiquitin–26S proteasome system at the nexus of plant biology. Nat. Rev. Mol. Cell Biol. 10: 385-397.
CUL1
SKP1
F-box
SCF E3 complex
26S proteosome
Target
Ubiquitin
E2
The ubiquitin pathway targets
proteins for destruction by the
proteosome.
32.
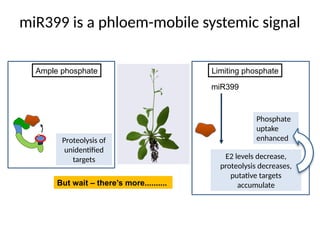
miR399 is aphloem-mobile systemic signal
Phosphate
uptake
enhanced
miR399
Proteolysis of
unidentified
targets
Ample phosphate Limiting phosphate
E2 levels decrease,
proteolysis decreases,
putative targets
accumulate
But wait – there’s more..........
33.
miRNAs and nutrientuptake- summary
•The rate of nutrient uptake in the roots is regulated by signals from the shoot
•PHO2 encodes a ubiquitin-conjugating E2 that presumably contributes to
directed proteolysis
•PHO2 levels are regulated by miR399 which accumulates upon phosphate
starvation
•Starvation-induced miR399 moves from shoot to root, regulating phosphate
uptake through PHO2
34.
Fig. 1. AmiRNA responds to an environmental abiotic stress in a stress-, species-, and miRNA-dependent manner.
Various miRNAs were aberrantly expressed under different abiotic stress treatments in a multiple plant species.
Red colour indicates up-regulated (u). Green colour indicates down-regulated (d). Yellow colour indicates that both
up-regulation and down-regulation were observed among different tissues or at different developmental stages.
The data were based on current literature of Arabidopsis, rice, wheat, barley, switchgrass, and cotton.
36.
The enhanced CSD1and CSD2 transcripts contribute to enhanced
detoxification of ROS, which can accumulate at high levels during stress.
miRNA and oxidative stress
• MicroRNA398 plays an important dual but opposite role during normal growth conditions and
abiotic stress.
• Both plant development and stress resistance pathways constitute a complex network
of multiple pathways.
Cu/Zn Superoxide
Dismutase 1
(CSD1)
37.
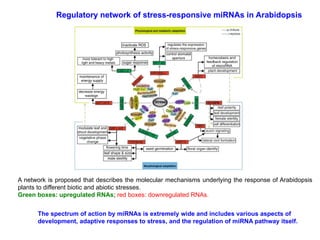
A network isproposed that describes the molecular mechanisms underlying the response of Arabidopsis
plants to different biotic and abiotic stresses.
Green boxes: upregulated RNAs; red boxes: downregulated RNAs.
Regulatory network of stress-responsive miRNAs in Arabidopsis
The spectrum of action by miRNAs is extremely wide and includes various aspects of
development, adaptive responses to stress, and the regulation of miRNA pathway itself.
38.
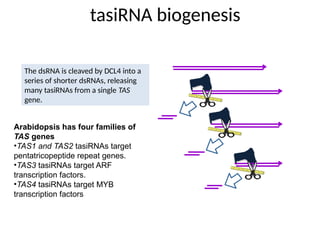
tasiRNA biogenesis
DICER
The dsRNAis cleaved by DCL4 into a
series of shorter dsRNAs, releasing
many tasiRNAs from a single TAS
gene.
Arabidopsis has four families of
TAS genes
•TAS1 and TAS2 tasiRNAs target
pentatricopeptide repeat genes.
•TAS3 tasiRNAs target ARF
transcription factors.
•TAS4 tasiRNAs target MYB
transcription factors
39.
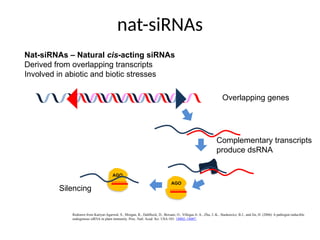
nat-siRNAs
Redrawn from Katiyar-Agarwal,S., Morgan, R., Dahlbeck, D., Borsani, O., Villegas Jr. A., Zhu, J.-K., Staskawicz, B.J., and Jin, H. (2006) A pathogen-inducible
endogenous siRNA in plant immunity. Proc. Natl. Acad. Sci. USA 103: 18002–18007.
Nat-siRNAs – Natural cis-acting siRNAs
Derived from overlapping transcripts
Involved in abiotic and biotic stresses
AGO
AGO
Overlapping genes
Complementary transcripts
produce dsRNA
Silencing
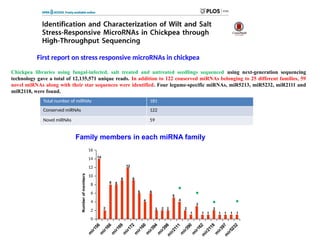
First report onstress responsive microRNAs in chickpea
Total number of miRNAs 181
Conserved miRNAs 122
Novel miRNAs 59
m
i
r
1
5
6
m
i
r
1
6
6
m
i
r
1
6
9
m
i
r
1
7
2
m
i
r
1
6
0
m
i
r
3
9
4
m
i
r
3
9
8
m
i
r
2
1
1
1
m
i
r
3
9
0
m
i
r
1
6
2
m
i
r
2
1
1
8
m
i
r
3
9
7
m
i
r
5
2
3
2
0
2
4
6
8
10
12
14
16
Number
of
members
14
2
8 8
9
12
9
6
4
6
2 2 2
5
4
2
1
3
1 1 1 1 1 1
2
Family members in each miRNA family
Chickpea libraries using fungal-infected, salt treated and untreated seedlings sequenced using next-generation sequencing
technology gave a total of 12,135,571 unique reads. In addition to 122 conserved miRNAs belonging to 25 different families, 59
novel miRNAs along with their star sequences were identified. Four legume-specific miRNAs, miR5213, miR5232, miR2111 and
miR2118, were found.
42.
miR ID SequenceLength MFE* Target gene
car-miRNA008 ACGAGACAGAUGGACACGACGG 22 -28.5
RING/U-box superfamily protein,
chalcone synthase (CHS)
car-miRNA011 AUGGUUGAGAGGGUGACUUGA 21 -72.3
SERPIN family protein, amelogenin,
RNA binding (RRM/RBD/RNP
motifs),LEA, anion channel protein
family
car-miRNA015 CGAGACAGAUGGACACGACGG 21 -28.5
Complex 1 protein (LYR family),
ribosomal L23/L15e family protein
car-miRNA020 CUGUAGCAUCACUAUAGCCGC 21 - 45
TPR-like superfamily protein, ARM
superfamily protein, FAD/NAD(P)-
binding oxidoreductase,
car-miRNA051 UGGGACAAUCGAUUUGGACAUC 22 -22.4
SMG7, HAD superfamily protein,
unique electron transfer flavoprotein
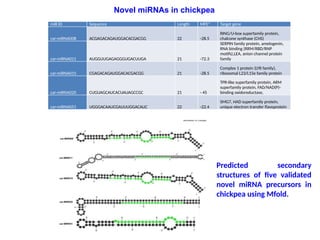
Novel miRNAs in chickpea
Predicted secondary
structures of five validated
novel miRNA precursors in
chickpea using Mfold.
43.
Mapping of targetmRNA cleavage site of
miR166 by modified RACE.
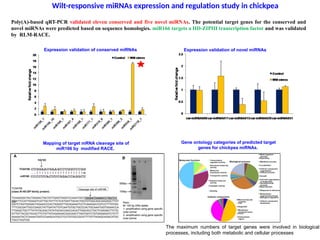
Wilt-responsive miRNAs expression and regulation study in chickpea
Expression validation of conserved miRNAs Expression validation of novel miRNAs
Poly(A)-based qRT-PCR validated eleven conserved and five novel miRNAs. The potential target genes for the conserved and
novel miRNAs were predicted based on sequence homologies. miR166 targets a HD-ZIPIII transcription factor and was validated
by RLM-RACE.
Gene ontology categories of predicted target
genes for chickpea miRNAs.
The maximum numbers of target genes were involved in biological
processes, including both metabolic and cellular processes
44.
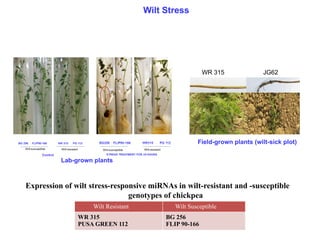
Wilt Stress
Expression ofwilt stress-responsive miRNAs in wilt-resistant and -susceptible
genotypes of chickpea
Wilt Resistant Wilt Susceptible
WR 315
PUSA GREEN 112
BG 256
FLIP 90-166
WR 315 JG62
Field-grown plants (wilt-sick plot)
Lab-grown plants
45.
BG 256 FLIP90-166 WR 315 PUSA
GREEN 112
0
0.5
1
1.5
2
2.5
3
3.5
CONTROL
STRESS
BG 256 FLIP 90-166 WR 315 PUSA GREEN
112
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
CONTROL
STRESS
Expression profiling of miR2118
Expression profiling of target gene of miR2118
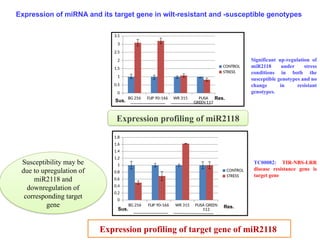
TC00082: TIR-NBS-LRR
disease resistance gene is
target gene
Significant up-regulation of
miR2118 under stress
conditions in both the
susceptible genotypes and no
change in resistant
genotypes.
Sus. Res.
Sus. Res.
Expression of miRNA and its target gene in wilt-resistant and -susceptible genotypes
Susceptibility may be
due to upregulation of
miR2118 and
downregulation of
corresponding target
gene
46.
BG 256 FLIP90 166 WR315 PUSA GREEN
112
0
0.2
0.4
0.6
0.8
1
1.2
CONTROL STRESS
Relative
fold
change
Susceptible Resistant
Resistance may be due
to upregulation of
miR530 and
downregulation of its
target gene
BG256 FLIP90-166 WR 315 PUSAGREEN
112
0
0.5
1
1.5
2
2.5
3
3.5
CONTROL STRESS
Relative
fold
change
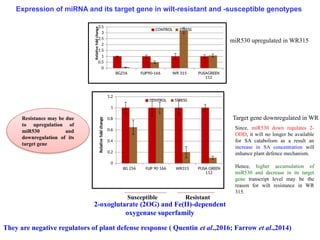
They are negative regulators of plant defense response ( Quentin et al.,2016; Farrow et al.,2014)
2-oxoglutarate (2OG) and Fe(II)-dependent
oxygenase superfamily
miR530 upregulated in WR315
Target gene downregulated in WR
Since, miR530 down regulates 2-
ODD, it will no longer be available
for SA catabolism as a result an
increase in SA concentration will
enhance plant defence mechanism.
Hence, higher accumulation of
miR530 and decrease in its target
gene transcript level may be the
reason for wilt resistance in WR
315.
Expression of miRNA and its target gene in wilt-resistant and -susceptible genotypes
47.
N39 N107 N108N6-87
-0.8
-0.6
-0.4
-0.2
0
0.2
0.4
0.6
0.8
1
1.2
Kabuli Flower Desi Flower
Relative
expression
-1
0
1
2
3
4
5
6
7
8 Kabuli Flower
Desi Flower
Relative
expression
Target prediction of miRNAs was done by using psRNATarget (//http://plantgrn.noble.org/psRNATarget/?
function=1).
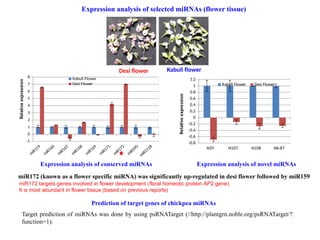
miR172 (known as a flower specific miRNA) was significantly up-regulated in desi flower followed by miR159
Desi flower Kabuli flower
Expression analysis of selected miRNAs (flower tissue)
Expression analysis of conserved miRNAs Expression analysis of novel miRNAs
Prediction of target genes of chickpea miRNAs
miR172 targets genes involved in flower development (floral homeotic protein AP2 gene).
It is most abundant in flower tissue (based on previous reports)
48.
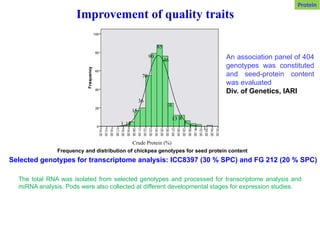
An association panelof 404
genotypes was constituted
and seed-protein content
was evaluated
Div. of Genetics, IARI
Frequency and distribution of chickpea genotypes for seed protein content
Selected genotypes for transcriptome analysis: ICC8397 (30 % SPC) and FG 212 (20 % SPC)
The total RNA was isolated from selected genotypes and processed for transcriptome analysis and
miRNA analysis. Pods were also collected at different developmental stages for expression studies.
1 14
15
36
70
90
85
46
16
13 8
5
1 1 1
Crude Protein (%)
Improvement of quality traits
Protein
49.
Genome-wide discovery andcharacterization of microRNAs and their target genes
associated with seed protein content
Genotypes used: ICC8397 (30 % SPC) and FG 212 (20 % SPC)
• Small RNA libraries were constructed for each sample in replicates
• 30-34 Mn reads were generated for each sample
Figure: Length distribution of miRNAs in all the chickpea libraries
The majority of small RNAs (approximately 50%) were 24 nt in
length, which is similar to other plant species.
Protein
50.
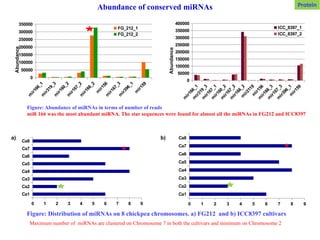
Figure: Abundance ofmiRNAs in terms of number of reads
miR 166 was the most abundant miRNA. The star sequences were found for almost all the miRNAs in FG212 and ICC8397
Abundance of conserved miRNAs
m
ir166_1
m
ir319_3
m
ir167_1
m
ir160_2
m
ir167_2
m
ir166_2
m
ir2118
m
ir156
m
ir166_3
m
ir167_3
m
ir396_1
m
ir159
0
50000
100000
150000
200000
250000
300000
350000
400000
ICC_8397_1
ICC_8397_2
Abundance
Ca1
Ca2
Ca3
Ca4
Ca5
Ca6
Ca7
Ca8
0 1 2 3 4 5 6 7 8 9
Ca1
Ca2
Ca3
Ca4
Ca5
Ca6
Ca7
Ca8
0 1 2 3 4 5 6 7 8 9
a) b)
Figure: Distribution of miRNAs on 8 chickpea chromosomes. a) FG212 and b) ICC8397 cultivars
Maximum number of miRNAs are clustered on Chromosome 7 in both the cultivars and minimum on Chromosome 2
0
50000
100000
150000
200000
250000
300000
350000
FG_212_1
FG_212_2
Abundance
Protein
51.
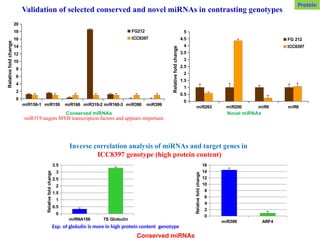
Validation of selectedconserved and novel miRNAs in contrasting genotypes
miR263 miR200 miR6 miR8
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
5
FG 212
ICC8397
Novel miRNAs
Relative
fold
change
miR156-1 miR159 miR166 miR319-2 miR160-3 miR398 miR399
0
2
4
6
8
10
12
14
16
18
20
FG212
ICC8397
Conserved miRNAs
Relative
fold
change
miRNA156 7S Globulin
0
0.5
1
1.5
2
2.5
3
3.5
Relative
fold
change
miR399 ARF4
0
2
4
6
8
10
12
14
16
Relative
fold
change
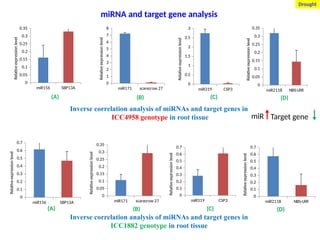
Inverse correlation analysis of miRNAs and target genes in
ICC8397 genotype (high protein content)
Conserved miRNAs
miR319 targets MYB transcription factors and appears important.
Protein
Exp. of globulin is more in high protein content genotype
52.
ICC1882
ICC4958
C S CS
A
(A) Root morphology of control and stressed plants of ICC 4958 and
ICC 1882 genotype (B) phenotypic variations of Pusa362 and SBD377
Pusa 362
C S
SBD 377
C S
Pusa 362
C S
SBD 377
C S
Pusa 362
C S
SBD 377
C S
B
Reduced
biomass
Genome-wide discovery and characterization of drought-
responsive microRNAs and their target genes
Drought tolerant Drought sensitive
ICC 4958 ICC 1882
PUSA 362 SBD 377
Stage: 45 day old plants subjected to stress of 42 -47 days
Stage 45 day old plants
RWC 55 % for stressed samples
(71 % for control samples)
Drought
53.
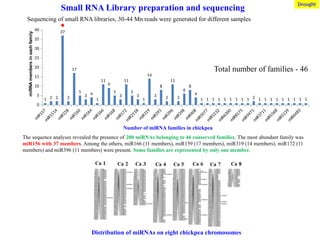
Small RNA Librarypreparation and sequencing
Sequencing of small RNA libraries, 30-44 Mn reads were generated for different samples
m
i
R
1
5
7
m
i
R
1
5
1
4
m
i
R
1
5
8
m
i
R
1
6
0
m
i
R
1
6
4
m
i
R
1
6
6
m
i
R
1
6
8
m
i
R
1
7
2
m
i
R
2
1
1
8
m
i
R
3
1
9
m
i
R
3
9
3
m
i
R
3
9
6
m
i
R
3
9
8
m
i
R
4
0
8
m
i
R
5
0
7
7
m
i
R
5
2
3
2
m
i
R
6
3
0
0
m
i
R
8
1
7
5
m
i
R
9
4
7
1
m
i
R
3
7
1
1
m
i
R
5
3
6
8
m
i
R
5
1
3
9
m
i
R
6
4
8
3
0
5
10
15
20
25
30
35
40
1 2 2
37
2
17
5
3 4
1
11
9
5
3
11
5
3
1
14
3
8
2
11
2
6
8
4
1 1 1 1 1 1 1 1 1 2 1 1 1 1 1 1 1 1 1
miRNA family
miRNA
members
in
each
family
Total number of families - 46
The sequence analyses revealed the presence of 200 miRNAs belonging to 46 conserved families. The most abundant family was
miR156 with 37 members. Among the others, miR166 (11 members), miR159 (17 members), miR319 (14 members), miR172 (11
members) and miR396 (11 members) were present. Some families are represented by only one member.
Number of miRNA families in chickpea
Distribution of miRNAs on eight chickpea chromosomes
Drought
54.
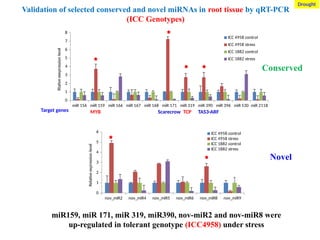
Validation of selectedconserved and novel miRNAs in root tissue by qRT-PCR
(ICC Genotypes)
miR 156 miR 159 miR 166 miR 167 miR 168 miR 171 miR 319 miR 390 miR 396 miR 530 miR 2118
0
1
2
3
4
5
6
7
8
ICC 4958 control
ICC 4958 stress
ICC 1882 control
ICC 1882 stress
Rlative
eexpression
level
nov_miR2 nov_miR4 nov_miR5 nov_miR6 nov_miR8 nov_miR9
0
1
2
3
4
5
6 ICC 4958 control
ICC 4958 stress
ICC 1882 control
ICC 1882 stress
Relative
expression
level
Conserved
Novel
miR159, miR 171, miR 319, miR390, nov-miR2 and nov-miR8 were
up-regulated in tolerant genotype (ICC4958) under stress
MYB Scarecrow TCP TAS3-ARF
Target genes
Drought
55.
miR 156 miR159 miR 166 miR 167 miR 168 miR 171 miR 319 miR 390 miR 396 miR 530 miR
2118
0
1
2
3
4
5
6
7
8
9 Pusa 362 control
Pusa 362 stress
SBD 377 control
SBD 377 stress
Relative
expression
level
nov_miR2 nov_miR4 nov_miR5 nov_miR6 nov_miR8 nov_miR9
0
2
4
6
8
10
12
14 Pusa 362 control
Pusa 362 stress
SBD 377 control
SBD 377 stress
Relative
expression
level
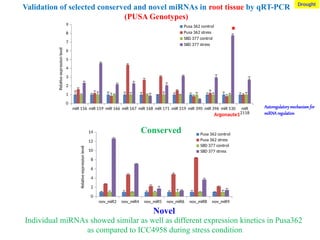
Individual miRNAs showed similar as well as different expression kinetics in Pusa362
as compared to ICC4958 during stress condition
Validation of selected conserved and novel miRNAs in root tissue by qRT-PCR
(PUSA Genotypes)
Conserved
Novel
Argonaute1
Drought
Autoregulatory mechanism for
miRNA regulation
56.
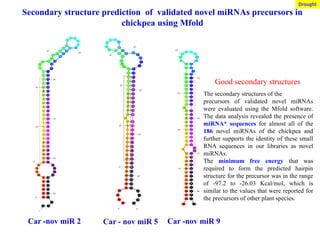
Car -nov miR2 Car - nov miR 5 Car -nov miR 9
Secondary structure prediction of validated novel miRNAs precursors in
chickpea using Mfold
Good secondary structures
The secondary structures of the
precursors of validated novel miRNAs
were evaluated using the Mfold software.
The data analysis revealed the presence of
miRNA* sequences for almost all of the
186 novel miRNAs of the chickpea and
further supports the identity of these small
RNA sequences in our libraries as novel
miRNAs.
The minimum free energy that was
required to form the predicted hairpin
structure for the precursor was in the range
of -97.2 to -26.03 Kcal/mol, which is
similar to the values that were reported for
the precursors of other plant species.
Drought
57.
miR168 WD repeatand HMG-box DNA-binding protein 1 (LOC101508348), hsp70-Hsp90 organizing protein 3
(LOC101494428), zinc finger protein ZAT4 (LOC101511854)
miR167 auxin response factor 6 (LOC101500671), zinc-finger homeodomain protein 11 (LOC101495604), F-box/LRR-
repeat protein At1g67190 (LOC101489858)
miR159 transcription factor GAMYB-like (LOC101495598), cytochrome P450 89A2-like (LOC101501122), AP-1 complex
subunit gamma (LOC101495931)
miR156 squamosa promoter-binding-like protein 9 (LOC101502532),
AUXIN SIGNALING F-BOX 2 (LOC101505746), MADS-box transcription factor 1-like (LOC101503539)
miR166 homeobox-leucine zipper protein ATHB-15 (LOC101493124),
transcription factor bHLH49 (LOC101491087), GRF1-interacting factor 3 (LOC101508094)
miR319 transcription factor TCP4-like (LOC101504111), E3 ubiquitin-protein ligase MBR2 (LOC101507426), GAMYB
(LOC101499054)
miR396 growth-regulating factor 4 (LOC101500728), ABSCISIC ACID-INSENSITIVE protein 2 (LOC101490892),
phytochrome A (LOC101496082)
miR390 PCTP-like protein (LOC101494413)
miR530 midasin-like (LOC101507441)
miR171 scarecrow-like protein 6 (LOC101512296), scarecrow-like protein 27 (LOC101512296)
miR2118 disease resistance protein (TIR-NBS-LRR class), auxin response factor 17 (LOC101514889)
Target gene prediction of validated conserved miRNAs by miRanda and psRNA target software
Several Transcription factors were identified as target genes of different miRNAs
Drought
miR171
1: Outer primer
2:Inner primer
M: 100 bp ladder
M 1 2
335 bp
miR166
1: Outer primer
2: Inner primer
M: 100 bp
M
1 2
1 2
M
243 bp
HD-bZIP gene
SCL gene
The target genes of miR166 and miR171 were experimentally validated by modified 5’RLM-RACE
The arrow indicates the cleavage site, and the numbers above the arrow denote the frequencies of the sequenced clones.
Mapping of target mRNA cleavage site
Drought
#2 The red symbol represents the small RNA. The green symbol with the AAAAA indicates an mRNA. The drawing on the bottom represents double stranded DNA being transcribed by RNA Polymerase to produce a transcript. Association of a small RNA with this transcript helps to target the enzymes that covalently modify chromatin (histone modifying enzymes and DNA methyltransferases) to confer silencing.
#4 MicroRNAs are encoded by MIR genes, fold into hairpin structures that are recognized and cleaved by DCL (Dicer-like) proteins.
#7 Note that mammals make do with one dicer, and insects and fungi with two. Like most components of the siRNA pathway, dicer-like genes are amplified in plants.
#29 The top figure shows expression levels of several miRNAs in response to different forms of nutrient starvation. The MIR399 genes are specifically induced during P starvation. Note that the MIR395 gene is induced upon S starvation. In the lower figure, the yellowness of the leaves in the mir399 overaccumulating plants is caused by overaccumulation of P to toxic levels. The measured amount of P in the leaves of the wild-type and mir399-overaccumulating plants is shown to the right; P accumulates in the shoots but not the roots.
#30 The top panel shows the structure of the PHO2 (aka E2) gene, indicating the putative miR399 binding sites. The lower panel shows that miR399 is induced by P starvation, whereas PHO2 expression is complementary to miR399 expression. The panel to the right shows that loss-of-function of PHO2 causes P overaccumulation (the plant on the left is a pho2 loss-of-function mutant and showing signs of P-toxicity) as well as P overaccumulation, indicated by the bar graph. PHO2 is a negative regulator of P accumulation; loss-of-function of PHO2 causes P overaccumulation.
#31 The target(s) affected by PHO2 accumulation are not known, but presumably these targets are targetted for proteolytic degradation by the PHO2/ E2 ligase in conjunction with the SCF E3 complex and the 26S proteosome.
#32 This slide summarizes the role of miR399 in signaling P starvation and promoting P uptake.
#54 TEOSINTE BRANCHED1, CYCLOIDEA, and PROLIFERATING CELL NUCLEAR ANTIGEN BINDING FACTOR (TCP)
scarecrow-like proteins (SCL6/22/27)
In plants, miR390 directs the production of tasiRNAs from TRANS-ACTING SIRNA3 (TAS3) transcripts to regulate AUXIN RESPONSIVE FACTOR (ARF) genes, critical for auxin signaling; these tasiRNAs are known as tasiARFs
#55 miR159 and 319 different
miR166 and miR 530 same