BMIC 1201
DNA
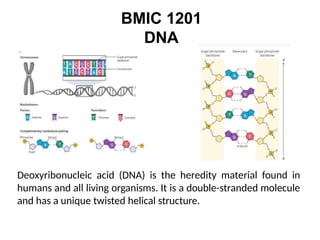
Deoxyribonucleic acid(DNA) is the heredity material found in
humans and all living organisms. It is a double-stranded molecule
and has a unique twisted helical structure.
2.
Genetic Elements
150 yearsago Gregor Johann Mendel---the
inheritance
patterns
Mendel to conclude that hereditary information is transmitted
in the form of distinct units.—gene/factor
genes consist of DNA sequences that encode
functional products—usually proteins, but in some
important cases RNAs.
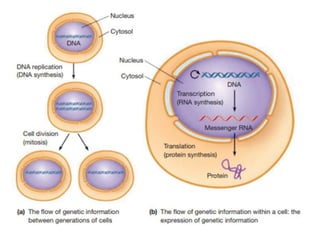
A cell’s DNA molecules undergo replication, generating two
DNA copies that are distributed to the daughter cells when the
cell divides.
The aspect of information flow --the DNA is expressed through
the processes of transcription (RNA synthesis) and translation
(protein synthesis).
4.
A Swissphysician Johann Friedrich Miescher
reported the discovery of the substance now known as
DNA in 1869, just a few years before the cell biologist
Walther Flemming first observed chromosomes as he
studied dividing cells under the microscope.
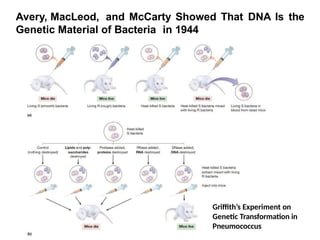
British physician and microbiologist Frederick Griffith, who
was studying a pathogenic strain of a bacterium, then
called “pneumococcus,” that causes a fatal pneumonia in
animals.
This bacterium (now called Streptococcus pneumoniae )
exists in two forms, called the S strain and the R strain.
When grown on a solid agar medium, the S strain
produces colonies that are smooth and shiny because of
the mucous polysaccharide coat each cell secretes,
whereas the R strain lacks the ability to manufacture a
mucous coat and therefore produces colonies exhibiting a
rough boundary
5.
Avery, MacLeod, andMcCarty Showed That DNA Is the
Genetic Material of Bacteria in 1944
Griffith’s Experiment on
Genetic Transformation in
Pneumococcus
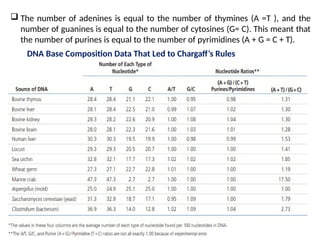
Chargaff’s Rules RevealThat A = T and G =
C
Erwin Chargaff was interested in the base composition of DNA.
Between 1944 and 1952, Chargaff used chromatographic
methods to separate and quantify the relative amounts of the
four bases—adenine (A), guanine (G), cytosine (C), and thymine
(T)—found in DNA.
DNA isolated from different cells of a given species has the same
percentage of each of the four bases.
The percentage does not vary with individual, tissue, age,
nutritional state, or environment.
Chargaff did find that DNA base composition varies from species
to species.
The relative amounts of the bases A and T versus G and C in the
DNAs of various organisms.
DNA preparations from closely related species have similar base
compositions, whereas those from very different species tend to
exhibit quite different base compositions.
8.
The numberof adenines is equal to the number of thymines (A =T ), and the
number of guanines is equal to the number of cytosines (G= C). This meant that
the number of purines is equal to the number of pyrimidines (A + G = C + T).
DNA Base Composition Data That Led to Chargaff’s Rules
9.
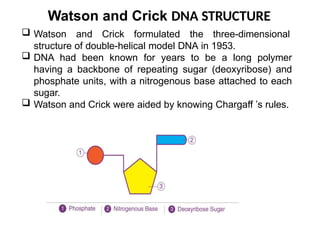
Watson and CrickDNA STRUCTURE
Watson and Crick formulated the three-dimensional
structure of double-helical model DNA in 1953.
DNA had been known for years to be a long polymer
having a backbone of repeating sugar (deoxyribose) and
phosphate units, with a nitrogenous base attached to each
sugar.
Watson and Crick were aided by knowing Chargaff ’s rules.
10.
The particularforms in which the bases A, G, C, and T exist at
physiological pH permit specific hydrogen bonds to form
between pairs of them. The crucial experimental evidence,
however, came from an X-ray diffraction picture of DNA
produced by Rosalind Franklin.
Franklin’s painstaking analysis of the diffraction pattern
revealed that DNA was a long, thin, helical molecule with one
type of structural feature repeated every 0.34 nm and another
repeated every 3.4 nm.
Watson and Crick approached the puzzle by building wire
models of possible structures. Based on the information
provided by Franklin’s picture, Watson and Crick eventually
produced a DNA model consisting of two intertwined strands—
a double helix.
11.
(a) The originalX-ray diffraction data generated by Rosalind Franklin and used by
Watson and Crick to construct their model.
(b)One of Watson and Crick’s original models, made with brass plates and wire.
The base has a hand-drawn scale indicating 10 nm.
Modeling DNA Structure
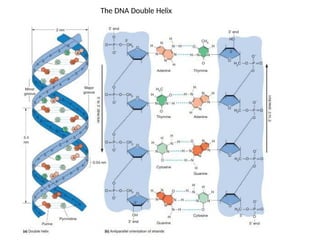
The Watson–Crick Model
In the Watson–Crick double Helix the sugar-phosphate backbones
of the two strands are on the outside of the helix, and the bases
face inward toward the center of the helix, forming the “steps” of
the “circular staircase” that the structure resembles.
The helix is right-handed, meaning that it curves “upward” to
the right.
It contains ten nucleotide pairs per turn and advances 0.34
nm per nucleotide pair. Consequently, each complete turn of the
helix adds 3.4 nm to the length of the molecule. The diameter
of the helix is 2 nm.
This distance turns out to be too small for two purines and too
great for two pyrimidines, but it accommodates a purine and a
pyrimidine well, consistent with Chargaff ’s rules.
The two strands are held together by hydrogen bonding between
the bases in opposite strands.
14.
The Watson–Crick Model
The hydrogen bonds holding together the two strands of the
double helix fit only when they form between the base adenine (A)
in one chain and thymine (T) in the other or between the base
guanine (G) in one chain and cytosine (C) in the other.
The base sequence of one chain determines the base sequence
of the opposing chain; the two chains of the DNA double helix are
therefore said to be complementary to each other. The standard
pairings referred to as Watson–Crick base pairings.
The Watson–Crick model suggested a mechanism by which cells
can faithfully replicate their genetic information: the two strands of
the DNA double helix could simply separate from each other
before cell division so that each strand could function as a
template, dictating the synthesis of a new complementary DNA
strand using Watson–Crick base-pairing rules.
The base A in the template strand would specify insertion of the
base T in the newly forming strand, the base G would specify
insertion of the base C, the base T would specify insertion of the
base A, and the base C would specify insertion of the base G.
15.
Structure and Compositionof DNA
DNA is made of two helical chains that intertwine with each
other to form a double helix. The most widely accepted structure
of DNA is right-handed helix DNA also known as the B-form of
DNA, which is 1.9 nm in diameter.
These
helical
chains run anti-parallel to
chain runs from 5’ to
3’
each other,
and the
one
other
polynucleotide
polynucleotide chain runs from 3’ to 5’. These chains are
connected to each other via nitrogen bases through
hydrogen
bonding.
Hydrogen bonding contributes to the specificity of base pairing.
Adenine preferentially pairs with Thymine through 2 hydrogen
bonds. Similarly, Cytosine preferentially pairs with Guanine
through 3 hydrogen bonds.
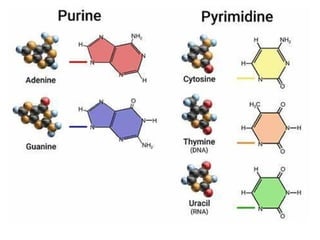
The base pairing happens when Pyrimidines pair with Purines
ring structure of
because Pyrimidines refers to the single
Thymine and Cytosine and Purines refers to double-ring
structures, Adenine and Guanine.
17.
Structure and Compositionof DNA
The base pairs A = T and G ≡ C are known as
complementary base pairs. Hence, the amount of Adenine
is equal to the amount of Thymine, and the amount of
Guanine is equal to the amount of Cytosine.
The geometry of the DNA is influenced by the distance between
the backbones and the angle at which the nitrogenous bases
are attached to the backbone.
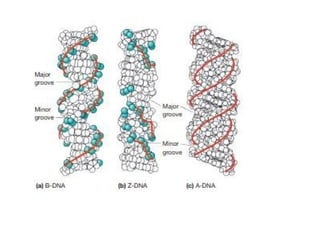
The major groove occurs when the backbones are far apart from
each other and the minor groove occurs when they are close.
The regularity of the helical structure forms two repeating and
alternating spaces: Major and Minor grooves.
18.
Structure and Compositionof DNA
These groves act on base-pair recognition and binding sites for
protein, the major groove contains base pair specific information
while the minor groove is largely base-pair nonspecific, caused
by protein interactions in the grooves
The double-helical structure of DNA is highly regular, each turn
of the helix measures approximately 10 base pairs. In addition to
hydrogen bonding in between the bases, the staging of bases
also stabilizes the structure, there are pi-pi interactions between
staged aromatic rings of the bases.
The distance between each turn is 3.4 nm.
The major groove is 2.2 nm wide and the minor groove is 1.1 nm
wide.
19.
Properties of DNA
Thetwo strands are twisted around each other so that there is a
major groove and a minor groove. Regulatory proteins often bind to
the major groove and recognize specific base sequences without
unfolding the DNA double helix.
DNA is made up of two helical strands that are coiled around the
same axis. If coiled from right it is known as right-handed helices
DNA and if coiled from left then it is known as left-handed helices.
However, the right-handed helices DNA is the most stable and thus
the structure of it is to be referred to as the standard.
The two chains of helices run antiparallel to each other. Thus, one
strand runs 5’ to 3’ and another strand runs from 3’ to 5’. The
opposite orientation of the two strands has important implications
for both DNA replication and transcription.
Both the strands denature on heating and can renature or
say hybridize on cooling. However, the temperature on which
these strands are separately permanently is referred to as
melting temperature and varies according to the specific sequence
of DNA.
20.
Properties of DNA
Forinstance, the region of higher concentration of C-G has a
higher melting temperature cause these bases are bonded
with three hydrogen bonds, which require more energy to
break than the region of higher concentration A-T which are
bonded only with two hydrogen bonds.
These nitrogenous bases store genetic information and thus
encode for amino acids which give rise to proteins.
The sequence of nucleotides found along the length of a piece
of DNA is a defining feature of DNA. In addition, DNA (and
RNA) can be characterized by size. Because each nucleotide
contains a nitrogenous base, and nucleotides in DNA pair
across the double helix, the length of DNA is measured in
base pairs (bp). Larger stretches of DNA are measured in
multiples of a single base pair. For example, a kilobase (kb) is
1000 bp, and a megabase (Mb) is 1,000,000 bp.
21.
Major and MinorGrooves of the DNA
As a result of the double helical nature of DNA, the molecule
has two asymmetric grooves. One groove is smaller than the
other.
This asymmetry is a result of the geometrical configuration of
the bonds between the phosphate, sugar, and base groups that
forces the base groups to attach at 120 degree angles instead
of 180 degree.
The larger groove is called the major groove, occurs when the
backbones are far apart; while the smaller one is called
the minor groove, occurs when they are close together.
Since the major and minor grooves expose the edges of the
bases, the grooves can be used to tell the base sequence of a
specific DNA molecule.
The possibility for such recognition is critical, since proteins must
be able to recognize specific DNA sequences on which to bind in
order for the proper functions of the body and cell to be carried out.
22.
Functions of DNA
DNA stores complete genetic information that requires to specify
an organism. Genes are the small segments of DNA, consisting
mostly of 250 – 2 million base pairs
It can replicate to give rise to two daughter cells and transfer
one copy to the daughter cells during cell division. Thus,
maintaining the genetic materials from generation to generation.
DNA is responsible for transcription for DNA to RNA.
It is the source of information that is needed in order to
synthesize cellular proteins, and other macromolecules required
by an organism.
It is responsible for identifying and determining the individuality
of the given organism.
It can also be taken as a targeted element during the diagnosis
of a particular disease.
Mutations: The changes which occur in the DNA sequences
Gene Therapy
23.
2. B-form
• Thestandard structure of DNA that is commonly known, was described
by Watson and Crick and is a right-handed double helix.
•The double-helical chains run antiparallel to each other, one running
from 5’ to 3’ and another running from 3’ to 5’ and are joined together via
complementary nitrogenous base pairing.
•Based upon Chargaff’s rules, bases coherent with another, only when
one purine of one strand pairs with one pyrimidine of another strand. A with
T and G with C
• The pair formed is a keto base pair, with an amino base, a purine with a
pyrimidine.
•The two strands of the DNA molecule are plectonemic coil meaning that
these two strands are coiled around the same axis and are intertwined with
each other.
• The consequence of this plectonemic coil is that these two strands can’t
be separated without the DNA rotating.
•The distance between the base pairs is 0.34 nm. One turn of the helix
contains 10 base pairs with a length of 3.4 nm.
• This form of DNA is 1.9 nm in diameter, which means the width of the
helix is 1.9 nm.
• The wide and shallow major groove of 2.2 nm, making it easily
assessable to proteins, and narrow and minor groove of 1.1 nm.
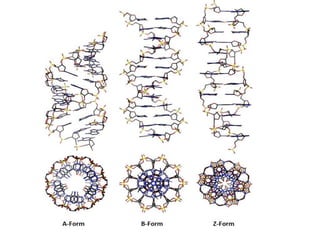
Types of DNA on the basis of forms
24.
Types of DNAon the basis of forms
1. A-form
•The major difference between the A and B forms of DNA is the
conformation of the deoxyribose sugar ring. For B form, it is in the C2
endoconformation, while in A form it is in the C3 endoconformation.
• Another important difference between A and B-form is
the
the
arrangement or placement of nitrogenous base pairs within
duplex.
•In B-form, the base pairs are almost in the center over the helical
axis, whereas in A-form, the base pairs are diverted away from the
central axis towards the major groove.
•The distance between two base pairs is 0.29 nm. One turn of the
helix contains 11 base pairs with a length of 2.8 nm
•Shorter than B-form of DNA. However, the helical width is 2.3 nm
which is more than B-form.
• Narrow and deep major groove and wide and shallow minor
groove.
•This form of DNA is favored by low hydration and by repeating
units of purines or pyrimidines.
25.
3. Z-form
•It isa left-handed helix and is a very different structure when
compared with the A and B-form.
•This form of DNA can form when the DNA is in alternating purines-
pyrimidines sequences.
• The backbone is not a smooth helix but an irregular zig-zag, which is
resulted from alternating sequences of purines and pyrimidines.
•The B form DNA can take the Z form when proteins are bound to DNA in
one helical conformation and force the DNA to adopt a different
conformation.
• This adoption happens at the G nucleotide, the sugar in this form is of
C3 endoconformation and the guanine base is in the synconformation.
• The result of which places the guanine back over the sugar ring, which
is unusual than the B and A form.
• It is long and thin than the B and A forms.
• The helical width is 1.8 nm, being the smallest among the three forms.
•The distance between the base pairs is 0.37 nm. One turn of the helix
contains 12 base pairs with a length of 4.56 nm.
• The major groove is flat and the minor groove is narrow and deep.
28.
Types of DNAon the basis of location
1. Nuclear DNA
•As the name suggests, these DNAs are located inside the nucleus organized in the
chromosome.
• These chromosomes are 43 pairs in humans and are linear with open ends and
contain 3 billion nucleotides.
•Nuclear DNA houses genes that are transcribed into mRNA and ultimately
translated to proteins, that are necessary for the functioning and maintaining the
integrity of the cell.
• It is inherited from both parents, so this is diploid and considered unique to each
individual except for identical twins.
• It is usually present in two copy numbers per cell
2. Mitochondrial DNA
• It is located inside the mitochondria.
• It is small and circular in structure
• It is inherited only from the mother, so is a haploid.
• It is present in a much higher copy number. i.e., 100-10,000 per cell.
• It has only 16,500 base pairs and encodes proteins that are
specificfor
mitochondria. These proteins are vital for producing energy.
•Mitochondrial DNA encoded proteins also play a pivotal role during intracellular
29.
Other Types ofDNA
1. D-DNA: A rare variation having eight base pairs per helical turn and no guanine in its
structure.
2. E- DNA: Extended or unusual DNA.
3. Spacer DNA: the nucleotide sequences that occur between genes; in eukaryotes, these
sequences are frequently lengthy and comprise several repeating sequences; in particular,
the DNA that occurs between genes encoding ribosomal RNA.
4. Complementary or copy DNA (cDNA): synthetic DNA transcribed from a specific RNA via the
reverse transcriptase enzyme process.
5. Recombinant DNA: Recombinant DNA is a DNA molecule made of linked sequences that do
not ordinarily occur in the same molecule, such as a bacterial plasmid into which a length of
viral DNA has been inserted.
6. Single copy DNA (scDNA): The bulk of gene sequences encoding polypeptides in eukaryotes
are single copy DNA (scDNA).
7. Repetitive DNA: Repetitive DNA consists of nucleotide sequences that occur several times
within a genome; they are characteristic of eukaryotes and do not typically encode
polypeptides. Clustered or dispersed sequences may be moderately (10 to 104 copies per
genome) or extensively (>106 copies per genome) repetitive. Some structural genes for
ribosomal RNA and histones are encoded by moderately repetitive DNA sequences; the
majority of highly repetitive sequences are satellite DNA.
30.
Bent DNA
Ingeneral, DNA sequences containing the adenine base are stiff and straight.
When A-tracts are replaced by other bases or when the helix collapses into the minor groove of A-tract,
DNA has a curved shape.
DNA structural bending has also been attributed to photochemical damage or mispairing of nucleotides.
Certain anticancer medications (e.g. cisplatin) induce bent structure in DNA. This altered structure can
absorb proteins that harm DNA.
Triple-stranded DNA
The creation of triple-stranded DNA may result from extra hydrogen bonds between the nucleotides.
Thus, a thymine can make two Hoogsteen hydrogen bonds with the adenine in an A-T pair to generate T-A-T.
Similarly, a protonated cytosine can establish two hydrogen bonds with guanine of G–C pairings,
resulting in the formation of C–G–C.
The triple helix is less stable than the double helix. This is because the triple helix’s three
negatively charged backbone strands result in a greater electrostatic repulsion.
An outline of Hoogsteen triple helical structure of DNA.
Four-stranded DNA
Extremely guanine-rich polynucleotides can create a unique tetrameric structure known as G-
quartets.
Hoogsteen hydrogen bonds hold these planar structures together.
G-tetraplexes, antiparallel four-stranded DNA structures, have also been found.
The telomeres at the ends of eukaryotic chromosomes, which are rich in guanine, create G-
tetraplexes.
In recent years, anti-cancer chemotherapies have focused on telomeres.
Other Types of DNA
31.
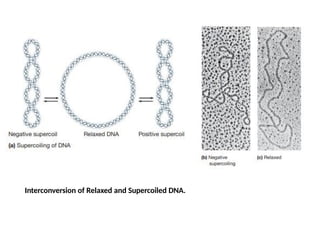
Relaxed and SupercoiledForms DNA
The DNA double helix can be twisted upon itself to a more compact
form called supercoiled DNA.
A DNA molecule can go back and forth between the supercoiled state
and the nonsupercoiled, or relaxed.
A relaxed DNA molecule can be converted to a positive supercoil by
twisting in the same direction as the double helix is wound and into a
negative supercoil by twisting in the opposite direction.
Circular DNA molecules found in nature, including those of bacteria,
viruses, and eukaryotic organelles, are invariably negatively
supercoiled.
Supercoiling also occurs in linear DNA molecules when regions of the
molecule are anchored to some cellular structure (for example, the
nuclear matrix) and so cannot freely rotate.
At any given time, significant portions of the linear DNA in the nucleus
of eukaryotic cells may be supercoiled; when DNA is packaged into
chromosomes at the time of cell division, extensive supercoiling helps
make the DNA more compact.
Relaxed and SupercoiledForms DNA
By Influencing both the spatial organization and the energy state
of DNA, supercoiling affects the ability of a DNA molecule to
interact with other molecules.
Positive supercoiling involves tighter winding of the double helix
and therefore reduces opportunities for interaction.
In contrast, negative supercoiling is associated with unwinding of
the double helix, which gives its strands increased access to
proteins involved in DNA replication or transcription. This
explains why negative supercoiling is favored in a cell.
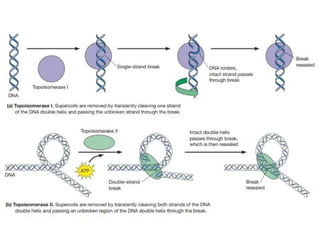
The interconversion between relaxed and supercoiled forms of
DNA is catalyzed by enzymes known as topoisomerases, which
are classified as either type I or type II.
34.
Relaxed and SupercoiledForms DNA
Both types catalyze the relaxation of supercoiled DNA; type I
enzymes do so by introducing transient single-strand breaks in
DNA, whereas type II enzymes introduce transient double-strand
breaks.
The temporary breaks affect DNA supercoiling.
Type I topoisomerases induce DNA relaxation by cutting one
strand of the double helix, thereby allowing the DNA to rotate and
the uncut strand to be passed through the break before the
broken strand is resealed.
In contrast, type II topoisomerases induce relaxation by cutting
both DNA strands and then passing a segment of uncut double
helix through the break before resealing.
Type I and type II topoisomerases are able to remove both
positive and negative supercoils from DNA.
36.
Relaxed and SupercoiledForms DNA
In addition, bacteria have a type II topoisomerase called DNA
gyrase, which can induce as well as relax supercoiling.
DNA gyrase is one of several enzymes involved in DNA
replication.
It can relax the positive supercoiling that results from partial
unwinding of a double helix, or it can actively introduce negative
supercoils that promote strand separation, thereby facilitating
access of other proteins involved in DNA replication. Like other
type II topoisomerases, DNA gyrase requires ATP to generate
supercoiling but not to relax an already supercoiled molecule.
37.
The Importance ofDNA Supercoiling
DNA supercoiling is important for DNA packaging within
all cells. Because the length of DNA can be thousands of
times that of a cell, packaging this genetic material into
the cell or nucleus (in eukaryotes ) is a difficult feat.
Supercoiling of DNA reduces the space and allows for
much more DNA to be packaged. In prokaryotes,
plectonemic supercoils are predominant, because of the
circular chromosome and relatively small amount of
genetic material.
In eukaryotes, DNA supercoiling exists on many levels of
both plectonemic and solenoidal supercoils, with the
solenoidal supercoiling proving the most effective in
compacting the DNA.
Solenoidal supercoiling is achieved with histones to form
a 10 nm fiber. This fiber is further coiled into a 30 nm
fiber, and further coiled upon itself numerous times more.
38.
The Importance ofDNA Supercoiling
DNA packaging is greatly increased during nuclear division
events such as mitosis or meiosis, where DNA must be
compacted and segregated to daughter cells.
Condensins and cohesins are structural maintenance of
chromosome (SMC) proteins that aid in the condensation of
sister chromatids and the linkage of the centromere in sister
chromatids. These SMC proteins induce positive supercoils.
Supercoiling is also required for DNA and RNA synthesis.
Because DNA must be unwound for DNA and RNA
polymerase action, supercoils will result.
The region ahead
of
unwound; this stress
the polymerase complex will be
is compensated with positive
supercoils ahead of the complex. Behind the complex, DNA
is rewound and there will be compensatory negative
supercoils.
39.
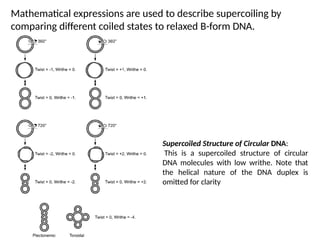
Mathematical expressions areused to describe supercoiling by
comparing different coiled states to relaxed B-form DNA.
Supercoiled Structure of Circular DNA:
This is a supercoiled structure of circular
DNA molecules with low writhe. Note that
the helical nature of the DNA duplex is
omitted for clarity
40.
As a generalrule, the DNA of most organisms is negatively
supercoiled. In a “relaxed” double-helical segment of B-DNA,
the two strands twist around the helical axis once every 10.4 to
10.5 base pairs of sequence. Adding or subtracting twists, as
some enzymes can do, imposes strain. If a DNA segment
under twist strain were closed into a circle by joining its two
ends and then allowed to move freely, the circular DNA would
contort into a new shape, such as a simple figure-eight. Such a
contortion is a supercoil.
The simple figure eight is the simplest supercoil, and is the shape a
circular DNA assumes to accommodate one too many or one too
few helical twists. The two lobes of the figure eight will appear
rotated either clockwise or counterclockwise with respect to one
another, depending on whether the helix is over or underwound.
For each additional helical twist being accommodated, the lobes
will show one more rotation about their axis.
41.
Supercoiled DNA formstwo structures; a plectoneme or a toroid, or a
combination of both. A negatively supercoiled DNA molecule will
produce either a one-start left-handed helix, the toroid, or a two-start
right-handed helix with terminal loops, the plectoneme. Plectonemes
are typically more common in nature, and this is the shape most
bacterial plasmids will take. For larger molecules, it is common for
hybrid structures to form – a loop on a toroid can extend into a
plectoneme. If all the loops on a toroid extend, it becomes a branch
point in the plectonemic structure.