Download to read offline





![6
PHYSICAL REVIEW RESEARCH 2, 012042(R) (2020)
Rapid Communications
Synaptic balance due to homeostatically self-organized quasicritical dynamics
Mauricio Girardi-Schappo ,1,*,†
Ludmila Brochini,2
Ariadne A. Costa,3
Tawan T. A. Carvalho ,4
and Osame Kinouchi 1,*,‡
1
Universidade de São Paulo, FFCLRP, Departamento de Física, Ribeirão Preto, SP, 14040-901, Brazil
2
Universidade de São Paulo, Instituto de Matemática e Estatística, São Paulo, SP, 05508-090, Brazil
3
Universidade Federal de Goiás - Regional Jataí, Unidade Acadêmica Especial de Ciências Exatas, Jataí, GO, 75801-615, Brazil
4
Universidade Federal de Pernambuco, Departamento de Física, Recife, PE, 50670-901, Brazil
(Received 30 July 2019; accepted 23 January 2020; published 20 February 2020)
Recent experiments suggested that a homeostatic regulation of synaptic balance leads the visual system to
recover and maintain a regime of power-law avalanches. Here we study an excitatory/inhibitory (E/I) mean-
field neuronal network that has a critical point with power-law avalanches and synaptic balance. When short-
term depression in inhibitory synapses and firing threshold adaptation are added, the system hovers around
the critical point. This homeostatically self-organized quasicritical (SOqC) dynamics generates E/I synaptic
current cancellation in fast timescales, causing fluctuation-driven asynchronous-irregular (AI) firing. We present
the full phase diagram of the model without adaptation varying external input versus synaptic coupling. This
system has a rich dynamical repertoire of spiking patterns: synchronous regular (SR), asynchronous regular
(AR), synchronous irregular (SI), slow oscillations (SO), and AI. It also presents dynamic balance of synaptic
currents, since inhibitory currents try and compensate excitatory currents over time, resulting in both of them
scaling linearly with external input. Our model thus unifies two different perspectives on cortical spontaneous
activity: both critical avalanches and fluctuation-driven AI firing arise from SOqC homeostatic adaptation and
are indeed two sides of the same coin.
DOI: 10.1103/PhysRevResearch.2.012042
Experimental and theoretical evidence suggests that spon-
taneous cortical activity happens in the form of asynchronous
irregular firing patterns (AI). This could be generated by
the balance of excitatory/inhibitory (E/I) synaptic currents
independent homeostatic mechanisms to generate the SOqC
dynamics: plasticity in the inhibitory synapses [18] and adap-
tive firing thresholds [19].
As for the second point, we will show that our homeo-](https://image.slidesharecdn.com/seminariogranada-200606110911/75/A-double-homeostatic-mechanism-to-get-self-organized-quasi-criticality-6-2048.jpg)
![The model
• N neurons (all-to-all graph)
• N
E
excitatory neurons, p = N
E
/N
• N
I
inhibitory neurons, q = N
I
/N p+q = 1
• a = E (excitatory) or I (inhibitory)
• Xi
a
= 1 (i-th neuron spikes)
• Xi
a
= 0 (i-th neuron remains silent)
• Vi
a
[t] = membrane potential at (discrete) time t
• Wij
ab
= synapses
• Ii = input
• 𝛉 = firing threshold
• 𝛍 = leakage parameter
7](https://image.slidesharecdn.com/seminariogranada-200606110911/75/A-double-homeostatic-mechanism-to-get-self-organized-quasi-criticality-7-2048.jpg)

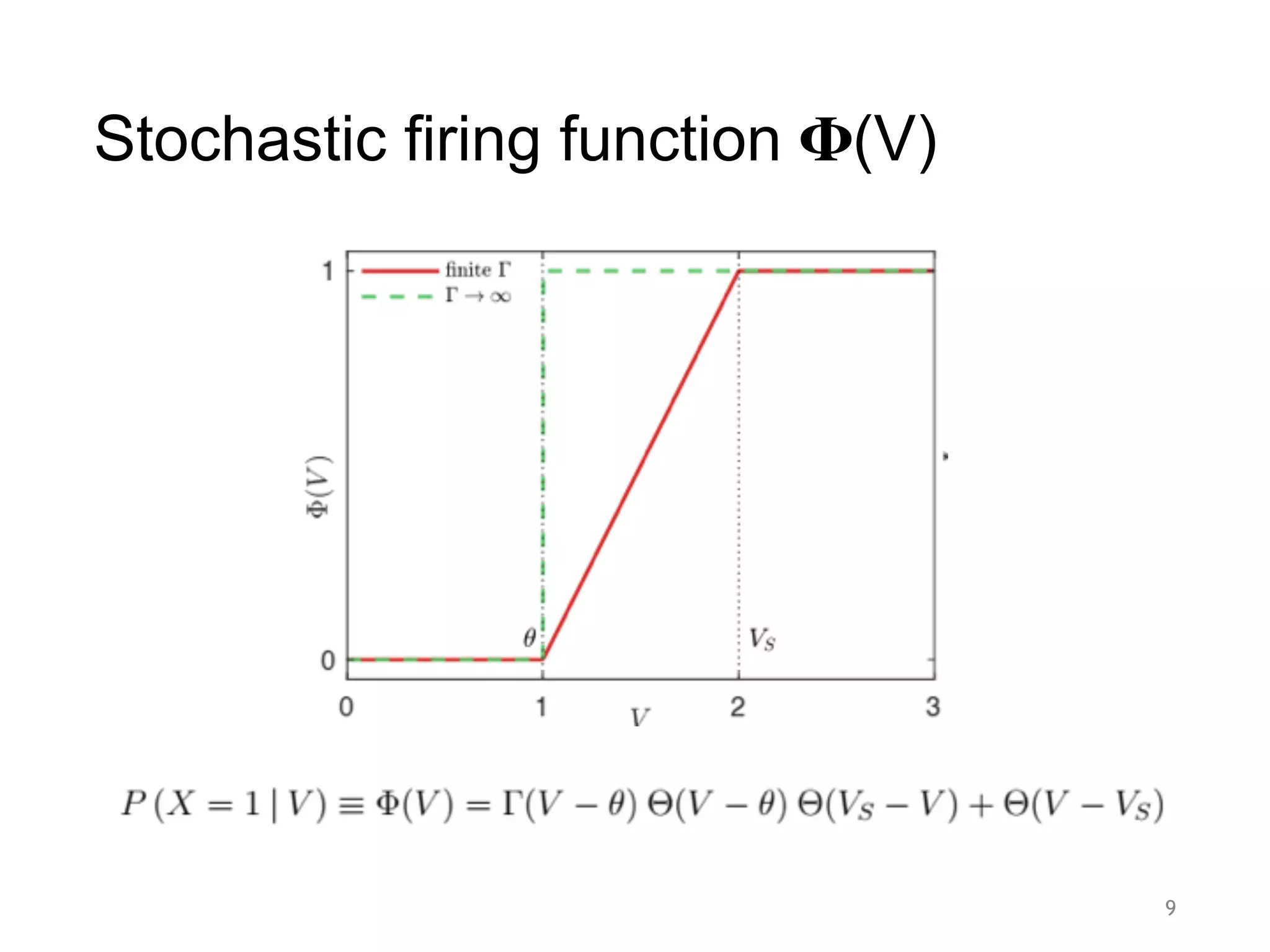
![Order and control parameters
10
𝛒a[t] = < Xa
j>[t] density of firing sites
Wab = < Wij
ab > average synaptic weight](https://image.slidesharecdn.com/seminariogranada-200606110911/75/A-double-homeostatic-mechanism-to-get-self-organized-quasi-criticality-10-2048.jpg)
![The quiescent (Q) or absorbing (𝛒0) phase
11
𝛒E = 𝛒I = 𝛒0 = 0
V[t+1] = 𝛍V[t] + I
Stationary: (1 - 𝛍) V* = I
Maximum V* for 𝛒0 = 0 is Vmax = 𝛉
Thus, Imax = (1 - 𝛍) 𝛉
Define field h = I - Imax = I - (1 - 𝛍) 𝛉
Thus transition to the absorbing state occurs at h = 0](https://image.slidesharecdn.com/seminariogranada-200606110911/75/A-double-homeostatic-mechanism-to-get-self-organized-quasi-criticality-11-2048.jpg)








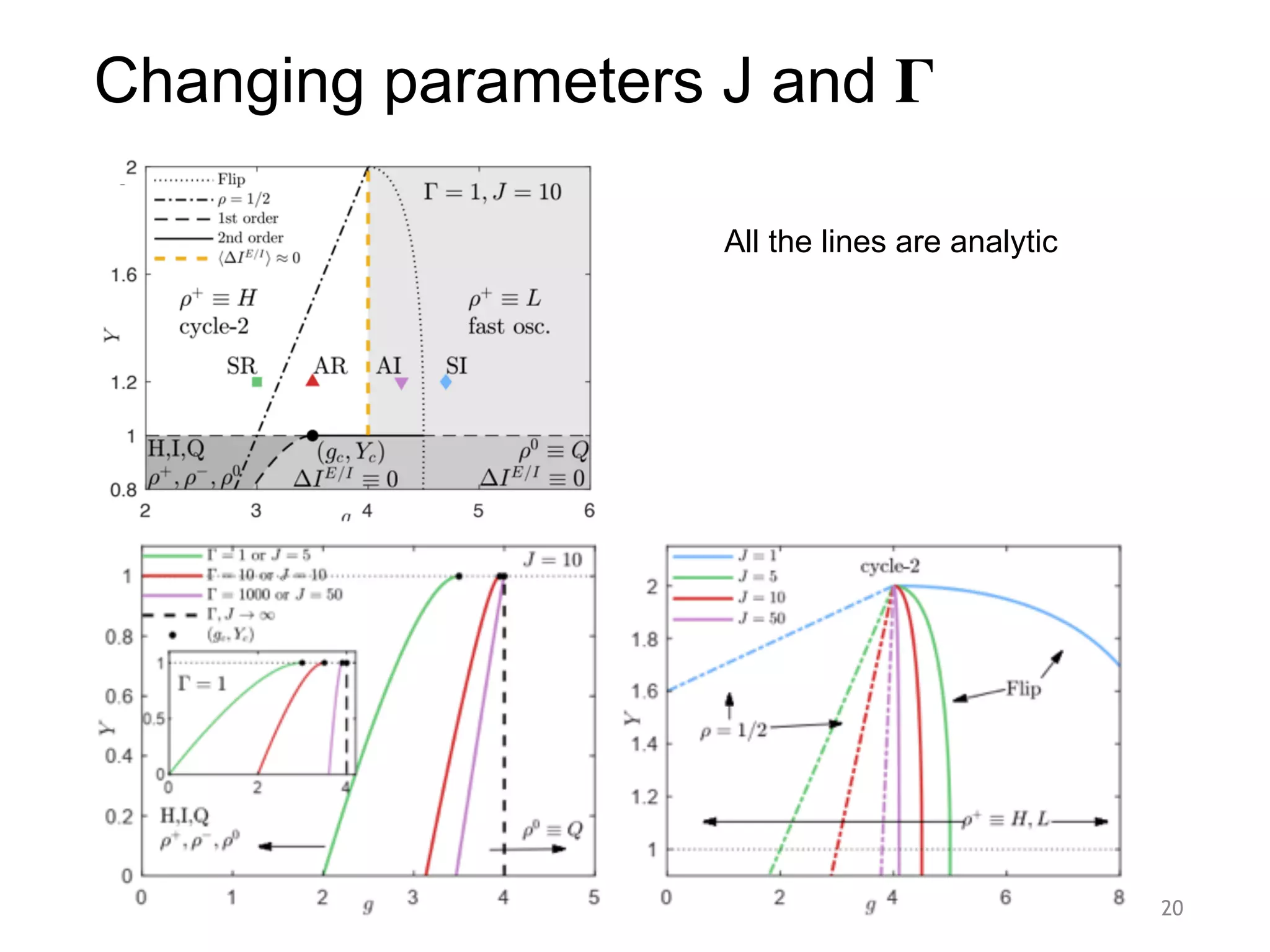


![.
23
2 3 4 5 6
0.8
1
1.2
AR
500 520 560 580
0
0.5
SR
0
0.5
AI
500 520 560 580
0
0.5
SI
0
0.5
SO
0
0.05
0 200 400 600 800 1000 1200 1400
0
0.01
0 2 4 6 8
-10 100
102
10-6
10-3
(c)
FIG. 2. Avalanches and firing patterns. (a) Phase diagram for µ = 0, and J = 10; a critical line starts at gc = 3.5, see Eq. (13), for Y = 1.
The critical point, the subcritical region with g > gFold; Y 1, and the supercritical region g = 4; Y > 1 have balanced synaptic currents, such
that the net current is IE/I
= IE
+ II
≈ 0. At Y = 1.2, from left to right: SR/cycle-2 (g = 3), AR/High (g = 3.5), AI/Low (g = 4.3), and
SI/fast oscillations (g = 4.7). SR and AR are separated by a bifurcation tho cycle-2 due to the refractory period; SI and AI are separated by
a flip bifurcation. (b) Distribution of avalanche sizes (main plot, τ = 1.5) and duration (bottom inset, τt = 2) at the critical point. Top inset:
size and duration scaling law s ∼ T a
has a crossover with a = 2.5 for small avalanches (a finite-size effect) and a = 2 for the rest of the
data. (c) Network simulation results (N = 106
neurons), ρ[t], for the points in (a). From the top left to the bottom right panel: critical point
absorbing-state avalanches (peaks); SR, AR, SO (slow waves for Y Yc = 1 − µ, µ = 0.9, Y = 0.101), SI, and AI. The background shows
the raster plot of 1,000 randomly selected neurons.
The variable ρ ≈ ρ1 = IE
/(pJ) is shown in the inset of
Fig. 1(b). These currents saturate for large enough J. This
linear scaling highlights the dynamic balance of synaptic
inputs, as inhibition tracks excitation over time [25,35].
Phase diagram. The soft threshold neurons’ phase diagram
is shown in Fig. 2(a). The curves are bifurcations of the stable
avalanches also respect the scaling law 1/(σνz) = (τt −
1)/(τ − 1) [inset in Fig. 2(b)], as expected for the DP uni-
versality class [30,36,37], and observed in experiments [38].
The simulated network activity in all the six dynami-
cal regimes is shown in Fig. 2(c). The critical point (gc =
3.5,Y = 1) displays avalanches sparked by a vanishing
PHYSICAL REVIEW RESEARCH 2, 012042(R) (2020)
6
R
R
AI
500 520 560 580
0
0.5
SI
0
0.5
SO
0
0.05
0
0 2 4 6 8
-10
-5
0
100
102
10-6
10-3
100
100
102
100
105
(b)
= 0, and J = 10; a critical line starts at gc = 3.5, see Eq. (13), for Y = 1.
Figure 8. Simulation of a network of 10,000 pyramidal cells and 2,500 interneurons, with connection probability 0.1 and JE = 0.1 mV. For
each of the four examples are indicated the temporal evolution of the global activity of the system (instantaneous firing frequency computed in
bins of 0.1 ms), together with the firing times (rasters) of 50 randomly chosen neurons. The instantaneous global activity is compared in each
SO
SISR
AI](https://image.slidesharecdn.com/seminariogranada-200606110911/75/A-double-homeostatic-mechanism-to-get-self-organized-quasi-criticality-23-2048.jpg)


![Homeostatic set point
26
In the W,h coordinates:
W* = [Wc+A/(𝛕WuW)/(1+1/(𝛕WuW)]
h* = 1/(c 𝛕 𝛉 u 𝛉)2
In the g, Y coordinates:
g* = gc / (1+1/(𝛕WuW))
+1/q (p-A/J) / (1+𝛕WuW)
Y* = Yc [1-1/( I c𝛕 𝛉 u 𝛉)2]
For large separation of time scales
(𝛕W , 𝛕 𝛉 > 100 ms):
W*, h*, g*, Y* → Wc , hc , gc , Yc](https://image.slidesharecdn.com/seminariogranada-200606110911/75/A-double-homeostatic-mechanism-to-get-self-organized-quasi-criticality-26-2048.jpg)


![Alternative homeostatic mechanisms
(only 𝛕 parameter)
• Wij[t+1] = Wij[t] + 1/𝛕 Wij[t] - Wij[t] Xj[t]
• Wij[t+1] = Wij[t] + 1/𝛕 - Wij[t] Xj[t]
• 𝚪i[t+1] = 𝚪i[t] + 1/𝛕 𝚪i - 𝚪i[t] Xi[t]
• 𝚪i[t+1] = 𝚪i[t] + 1/𝛕 - 𝚪i[t] Xi[t]
• 𝛉i[t+1] = 𝛉i[t] - 1/𝛕 + 𝛉i[t] Xi[t]
29](https://image.slidesharecdn.com/seminariogranada-200606110911/75/A-double-homeostatic-mechanism-to-get-self-organized-quasi-criticality-29-2048.jpg)




This document discusses several papers related to self-organized criticality in neural networks. It begins by summarizing Turing's 1950 speculation that subcritical, critical, and slightly supercritical branching processes could describe human and animal minds. It then discusses later work in 1995 and 2003 providing experimental support for the idea that neural networks operate near a critical point, enhancing information processing. The document proposes a new mechanism for achieving criticality - dynamic neuronal gains related to firing rate adaptation - instead of mechanisms related to synaptic dynamics studied previously. It concludes by discussing a 2020 paper finding that inhibitory synaptic depression and firing threshold adaptation can lead a neural network model to hover near criticality in a self-organized quasicritical state, maintaining synaptic balance.


























































