3. RESTING MEMBRANE POTENTIAL
Cells may use the membrane poten1al as a source of energy
• Secondary ac1ve transport
• ATP synthesis on chloroplast membranes
Or may modify the res1ng membrane poten1al to generate signals (excitable cells)
For a res(ng Em of -80 mV and a thickness of 60 A ̊ ,
the voltage gradient sustained across the
membrane is 133.000 V/cm. Thus, the cell
membrane tolerates an enormous voltage gradient.
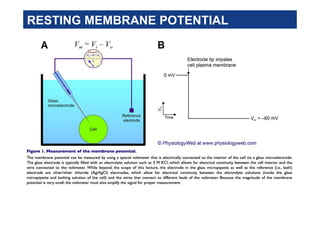
4. Voltmeter
Microelectrode inside cell
Plasma membrane Ground electrode outside cell
Neuron
Axon
Generated by:
Differences in ionic makeup of ICF and ECF
Differential permeability of the plasma membrane
Potential difference across the membrane of a
resting cell
Approximately –70 mV in neurons (cytoplasmic side of
membrane is negatively charged relative to outside)
RESTING MEMBRANE POTENTIAL
ICF: intracellular fluid
ECF: extracellular fluid
7. Figure 3. Two solu/on compartments separated by a
membrane that contains potassium (K+) channels.
(A) If two solu*on compartments are separated by a
membrane that contains selec*ve ion channels for
only one ion species (in this case, K+ channels), the
ion (K+) can diffusion down its concentra*on gradient
from one compartment to the other (i to o). The
counterion (Cl−) cannot follow because no selec*ve
channels exist for this ion. Thus, the net movement of
K+ from i to o leads to the addi*on of net posi*ve
charge to compartment o and leaves behind net
nega*ve charge in compartment i. This leads to
charge separa*on and, hence, a voltage poten*al
difference across the membrane.
RESTING MEMBRANE POTENTIAL
8. (A) If two solu*on compartments are separated by
a membrane that contains selec*ve ion channels
for only one ion species (in this case, K+ channels),
the ion (K+) can diffusion down its concentra*on
gradient from one compartment to the other (i to
o). The counterion (Cl−) cannot follow because no
selec*ve channels exist for this ion. Thus, the net
movement of K+ from i to o leads to the addi*on
of net posi*ve charge to compartment o and
leaves behind net nega*ve charge in compartment
i. This leads to charge separa*on and, hence, a
voltage poten*al difference across the membrane.
(B) As K+ diffuses from i to o, an electrical
gradient (voltage poten*al difference) is
established across the plasma membrane
that grows in size un*l it exactly balances the
K+ chemical gradient (i.e., concentra*on
gradient). When the chemical and electrical
gradients are equal in size, the ion is said to
be in electrochemical equilibrium, and the
membrane poten*al established is the
equilibrium poten*al (Veq.) for the ion.
(C) In the system described above, opening
of K+ channels, leads to the establishment
of a membrane poten*al, which grows in
size un*l it reaches the K+ equilibrium
poten*al (VK). The magnitude of VK
depends on the K+ concentra*on gradient
across the membrane. In this scenario,
based on the intracellular (150 mM) and
extracellular (5 mM) concentra*ons of K+,
Vm = VK = −90 mV.
Figure 3. Two solu/on compartments separated by a membrane that contains potassium (K+) channels.
RESTING MEMBRANE POTENTIAL
Equilibrium poten/al
9. NERNST EQUATION (Walther Nernst, 1887)
The Nernst equa-on calculates the equilibrium poten-al (also referred to as the Nernst poten-al) for an
ion based on the charge on the ion (i.e., its valence) and its concentra*on gradient across the membrane
• VEq. is the equilibrium poten-al (Nernst poten-al) for a given ion. If only one ionic species is present
in the system, and/or channels for only one ionic species are present (and open), then VEq. will also
be the membrane poten-al (Vm).
• R is the universal gas constant and is equal to 8.314 J.K-1.mol-1 (Joules per Kelvin per mole).
• T is the temperature in Kelvin (K = °C + 273.15).
• z is the valence of the ionic species. For example, z is +1 for Na+, +1 for K+, +2 for Ca2+, -1 for Cl-, etc.
• F is the Faraday's constant and is equal to 96485 C.mol-1 (Coulombs per mole).
• [X]out is the concentra-on of the ionic species X in the extracellular fluid.
• [X]in is the concentra-on of the ionic species X in the intracellular fluid.
RESTING MEMBRANE POTENTIAL
11. For the Nernst poten*al to be established,
selec*ve channels must be present and open.
Only three factors are important in
determining the magnitude and direc*on of
the Nernst poten*al:
(1) the concentra-on gradients
(2) the valence of the ionic species
(3) temperature.
If there is no concentra*on gradient across
the plasma membrane, the ra*o of the
extracellular to intracellular concentra*on
terms becomes 1. The natural logarithm of 1
is zero. Hence, there will be no poten*al
difference across this membrane, even if the
channels for the ion were fully open.
Therefore, a concentra*on difference is
essen*al in establishing a Nernst poten*al.
The valence of the ion also influences the
magnitude and direc*on of the Nernst
poten*al for the ion.
Temperature (T) also plays a small role in
determining the magnitude of the Nernst
poten*al.
RESTING MEMBRANE POTENTIAL
14. GOLDMAN-HODGKIN-KATZ EQUATION
In living cells, the res-ng membrane poten-al (Vm) is seldom governed by only one ion such as K+, Na+,
Cl-, etc. If this were the case, the membrane poten-al could be predicted by the equilibrium poten-al
(VEq.) for that ion, and could be easily calculated by using the Nernst equa-on. Instead, the membrane
poten-al is generally established as a result of the rela-ve contribu-ons of several ions. In many cells, K+,
Na+, and Cl- are the main contributors to the membrane poten-al.
• VEq. is the equilibrium poten0al (Nernst poten0al) for a given ion. If only one ionic species is present in the system, and/or channels for
only one ionic species are present (and open), then VEq. will also be the membrane poten0al (Vm).
If two or more ions contribute to the membrane poten0al, the Nernst poten0al no longer yields the Vm. In this case, use the Goldman-
Hodgkin-Katz (GHK) equa0on to calculate the Vm.
• R is the universal gas constant and is equal to 8.314 J.K-1.mol-1 (Joules per Kelvin per mole).
• T is the temperature in Kelvin (K = °C + 273.15).
• z is the valence of the ionic species. For example, z is +1 for Na+, +1 for K+, +2 for Ca2+, -1 for Cl-, etc. Note that z is unitless.
• F is the Faraday's constant and is equal to 96485 C.mol-1 (Coulombs per mole).
• [X]out is the concentra0on of the ionic species X in the extracellular fluid.
• [X]in is the concentra0on of the ionic species X in the intracellular fluid.
RESTING MEMBRANE POTENTIAL
17. Voltmeter
Microelectrode inside cell
Plasma membrane Ground electrode outside cell
Neuron
Axon
Generated by:
Differences in ionic makeup of ICF and ECF
Differential permeability of the plasma membrane
Potential difference across the membrane of a
resting cell
Approximately –70 mV in neurons (cytoplasmic side of
membrane is negatively charged relative to outside)
RESTING MEMBRANE POTENTIAL
ICF: intracellular fluid
ECF: extracellular fluid
18. The Na+/K+-ATPase helps maintain res5ng poten5al, avail
transport, and regulate cellular volume.
It also func5ons as signal transducer/integrator to regulate MAPK
pathway, ROS, as well as intracellular calcium.
In most animal cells, the Na+/K+-ATPase is responsible for about
1/5 of the cell's energy expenditure.
For neurons, the Na+/K+-ATPase can be responsible for up to
2/3 of the cell's energy expenditure.
Only under ideal conditions!!!
30-32 mol ATP more realistic
19. IN A REAL CELL, THE HIGH EXTRACELLULAR Na+ CONCENTRATION
NEUTRALIZES OSMOTIC FORCES...
RESTING MEMBRANE POTENTIAL
THE Na+/K+-ATPase MAINTAINS THE ASYMETRIC DISTRIBUTION OF Na+ AND K+...
...AND CONTRIBUTES TO THE RESTING MEMBRANE POTENTIAL (ELECTROGENIC)
Na+
K+ 3
2
20. • Differences in ionic makeup (Na/K+ ATPase)
• ICF has lower concentration of Na+ and Cl– than ECF
• ICF has higher concentration of K+ and negatively
charged proteins (A–) than ECF
Na+
K+
Na+ K+
RESTING MEMBRANE POTENTIAL
21. Na+
K+
Na+ K+
• Differences in ionic makeup (Na/K+ ATPase)
• ICF has lower concentration of Na+ and Cl– than ECF
• ICF has higher concentration of K+ and negatively
charged proteins (A–) than ECF
RESTING MEMBRANE POTENTIAL
22. Voltmeter
Microelectrode inside cell
Plasma membrane Ground electrode outside cell
Neuron
Axon
Generated by:
Differences in ionic makeup of ICF and ECF
Differential permeability of the plasma membrane
Potential difference across the membrane of a
resting cell
Approximately –70 mV in neurons (cytoplasmic side of
membrane is negatively charged relative to outside)
RESTING MEMBRANE POTENTIAL
23. • Differential permeability of membrane
• Impermeable to A–
• Slightly permeable to Na+ (leakage channels)
• 75 times more permeable to K+ (more leakage channels)
Na+
K+ K+
Na+
K+
Na+ K+
RESTING MEMBRANE POTENTIAL
24. • Negative interior of the cell is due to much greater diffusion of K+ out of
the cell than Na+ diffusion into the cell
• Sodium-potassium pump stabilizes the resting membrane potential by
maintaining the concentration gradients for Na+ and K+
Na+
K+ K+
RESTING MEMBRANE POTENTIAL
26. Depolarizing stimulus
Time (ms)
Inside
positive
Inside
negative
Resting
potential
Depolarization
(a) Depolarization: The membrane potential
moves toward 0 mV, the inside becoming
less negative (more positive).
Depolarization
A reduction in membrane potential (toward zero)
Inside of the membrane becomes less negative
than the resting potential
Increases the probability of producing a nerve
impulse
RESTING MEMBRANE POTENTIAL
27. Hyperpolarizing stimulus
Time (ms)
Resting
potential
Hyper-
polarization
(b) Hyperpolarization: The membrane
potential increases, the inside becoming
more negative.
Hyperpolarization
An increase in membrane potential (away from zero)
Inside of the membrane becomes more negative
than the resting potential
Reduces the probability of producing a nerve
impulse
RESTING MEMBRANE POTENTIAL
28. Voltmeter
Microelectrode inside cell
Plasma membrane Ground electrode outside cell
Neuron
Axon
Generated by:
Differences in ionic makeup of ICF and ECF
Differential permeability of the plasma membrane
Potential difference across the membrane of a
resting cell
Approximately –70 mV in neurons (cytoplasmic side of
membrane is negatively charged relative to outside)
RESTING MEMBRANE POTENTIAL
29. Two main types of channels:
Leakage channels (non-gated)—always open!!
Gated channels (three types):
Ligand-gated channels
open with binding of a specific neurotransmitter
Voltage-gated channels
open and close in response to changes in membrane potential
Mechanically gated channels
open and close in response to physical deformation of receptors
(a) Chemically (ligand) gated ion channels open when the
appropriate neurotransmitter binds to the receptor,
allowing (in this case) simultaneous movement of
Na+ and K+.
(b) Voltage-gated ion channels open and close in response
to changes in membrane voltage.
RESTING MEMBRANE POTENTIAL