

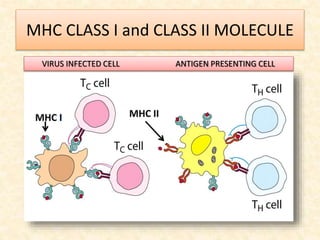












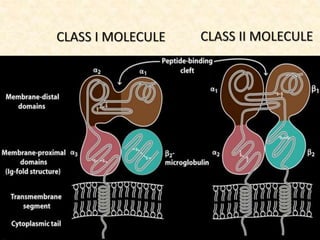
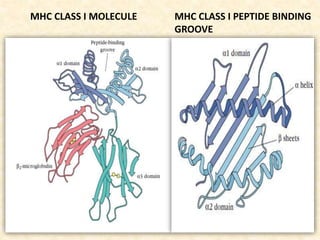
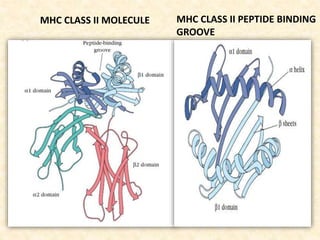




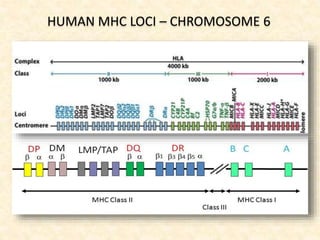
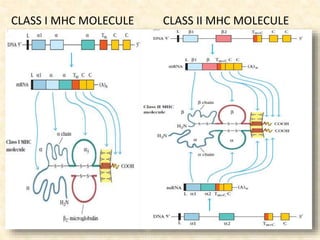
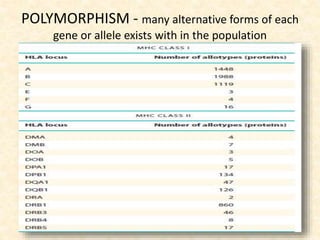
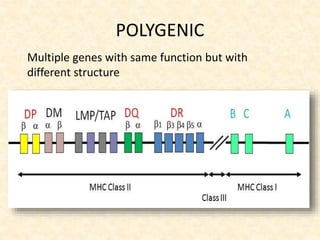

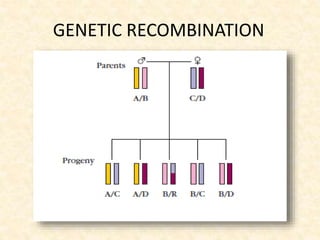
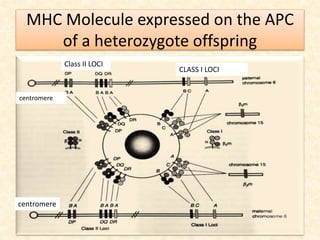
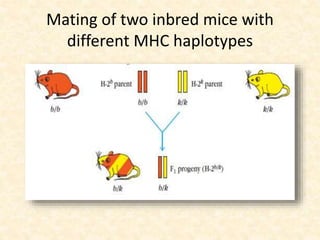
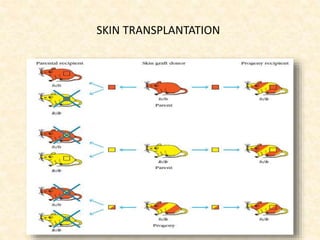











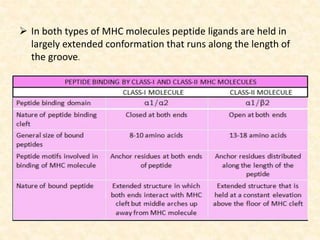
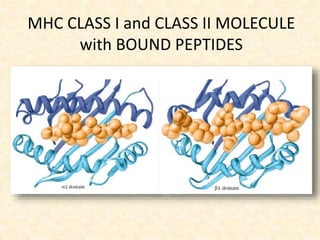
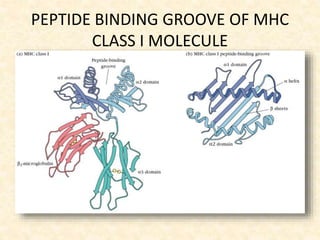
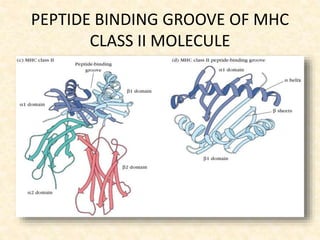


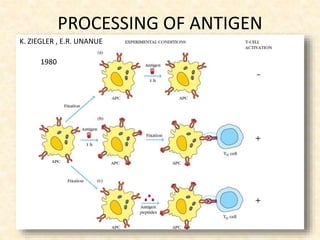
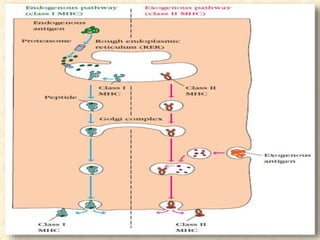

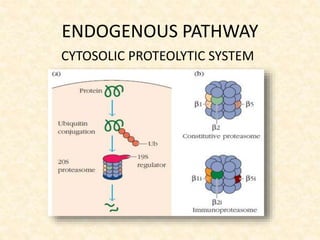

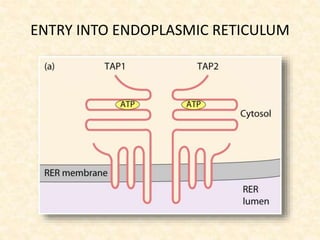
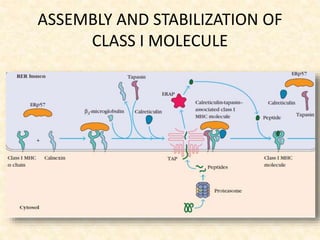
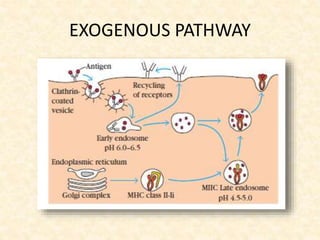

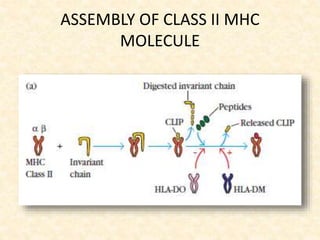
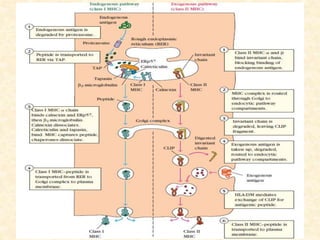




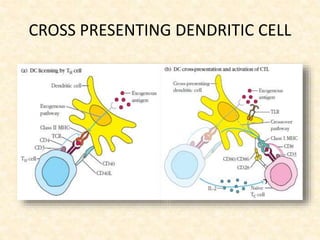








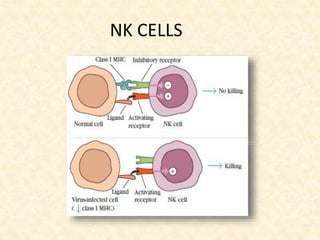



This document provides information on MHC class I and class II molecules, including their structure, function, and role in antigen presentation. It discusses that MHC class I molecules are expressed on all nucleated cells and present intracellular antigens to CD8+ T cells. MHC class II molecules are expressed primarily on antigen presenting cells and present extracellular antigens to CD4+ T cells. The peptide binding grooves of MHC class I and II molecules differ in their structure and the size of peptides they can bind.











































































![谷歌留痕技术 [ 𝙩𝙤𝙥 𝟮𝟯𝟯. 𝙘 𝙤𝙢 ]](https://cdn.slidesharecdn.com/ss_thumbnails/top233-260130174328-3833018c-thumbnail.jpg?width=640&height=640&fit=bounds)
