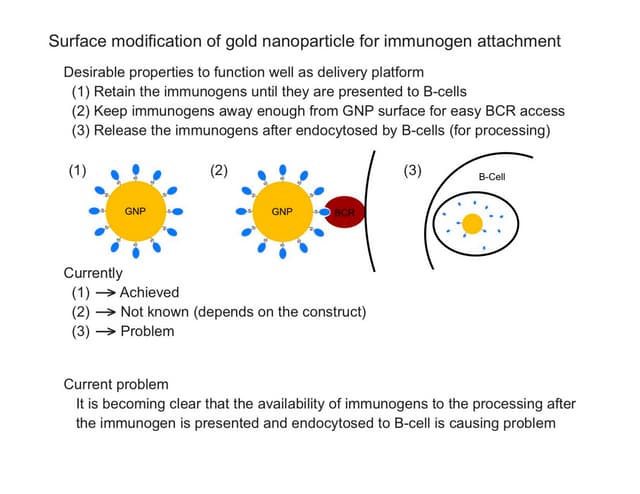
Monovalent cations like sodium ions can induce the formation of native tertiary structure in the Tetrahymena thermophila ribozyme in the absence of divalent ions like magnesium. Using hydroxyl radical footprinting and analytical ultracentrifugation, the researchers monitored the formation of individual tertiary contacts and global compaction of the ribozyme as a function of sodium ion concentration. They found that most native tertiary contacts form at high sodium ion concentrations, though a few contacts near where divalent ions bind require magnesium. Both native and non-native tertiary contacts can form at low sodium ion concentrations. The results suggest that monovalent ions can push the ribozyme into the native folded state by forming native
![letters
nature structural biology • advance online publication 1
Monovalent cations
mediate formation of
native tertiary structure of
the Tetrahymena
thermophila ribozyme
Keiji Takamoto1,2, Qin He1,2, Stephanie Morris3,
Mark R. Chance1–3 and Michael Brenowitz2,3
1Department of Physiology and Biophysics, 2Center for Synchrotron
Biosciences and 3Department of Biochemistry, Albert Einstein College of
Medicine, 1300 Morris Park Avenue, Bronx, New York 10461, USA.
Published online 18 November 2002; doi:10.1038/nsb871
The formation of individual tertiary contacts of the
Tetrahymena L-21 Sca I ribozyme has been monitored by
hydroxyl radical footprinting and its global conformation
by analytical ultracentrifugation as a function of mono-
valent ion concentration in the absence of divalent ions.
Advanced methods of data analysis, which allow the hydro-
xyl radical reactivity of every nucleotide to be quantified,
permit monitoring of each and every structural element of
the RNA. Monovalent ion-mediated global compaction of
the ribozyme is accompanied by the formation of native
tertiary contacts; most native tertiary contacts are evident
except several that are located near where divalent ions are
observed in crystallographic structures. Non-native tertiary
contacts are also observed at low but not high concentra-
tions of monovalent ions. In light of recent studies that have
shown that the presence of monovalent ions greatly acceler-
ates the Mg2+-dependent folding of the Tetrahymena
ribozyme, the present studies suggest that Na+ concentra-
tion changes not only the starting position of the RNA on its
folding funnel but also pushes it deep into the well by form-
ing native tertiary contacts and, thus, favoring fast and cor-
rect folding pathways.
Although Mg2+ has been long known to be required for the
folding and catalytic activity of the Tetrahymena thermophila
group I ribozyme1, monovalent cations can compact the
ribozyme2–5 and accelerate and dictate the predominant path-
ways it uses for folding6–12. Here we explore the ability of a
monovalent cation (Na+), in the absence of magnesium, to mod-
ulate the structure of the Tetrahymena Sca I ribozyme. Hydroxyl
radical ‘footprinting’ and analytical ultracentrifugation are used
to demonstrate the formation of native and non-native, and dis-
ruption of non-native, tertiary contacts by Na+ alone. The impli-
cations of these results for the acceleration of rates of folding9,11
and the partitioning of RNA among multiple pathways10 are dis-
cussed.
Single nucleotide structural analysis
Hydroxyl radical (OH•) footprinting titrations of the Sca I
ribozyme were conducted as a function of [NaCl] in the absence
of Mg2+. To obtain a comprehensive and objective OH• reactivity
map with single nucleotide resolution, a ‘single-band’ analysis
protocol13,14 was developed and applied to the autoradiograms
obtained for the Tetrahymena ribozyme (Fig. 1a–c). Hydroxyl
radical footprinting reports the solvent accessibility of individ-
ual nucleotides. Experiments conducted with RNA separately
labeled at the 3′ and 5′ termini, together with increasing dura-
tions of electrophoretic separation, allow almost all the
nucleotides of the ribozyme to be quantitatively analyzed
(Fig. 1d). The use of the single-band quantitative analysis and
display techniques may account for the observation of
Na+-dependent effects on formation of specific tertiary contacts
that have previously gone undetected15.
The results of this analysis are summarized in a false-color
map (Fig. 2) whose color mapping is referenced to the lowest
[Na+] studied (CE buffer), which was the starting point for bio-
chemical analysis of the Tetrahymena ribozyme16–18. The bar
below the Na+ titration (Fig. 2) summarizes the OH• reactivity
profile of the ribozyme folded in the presence of Mg2+, which is
0 100 200 300 400
0
100
200
300
400
500
600
700
800
Lane 1
Lane 2
Pixels
Intensity
22 409
54-91
64-134
79-152
85-210
141-205
171-246 313-395
289-368
258-317
266-315
202-276
3'5'
L-21 Sca I Ribozyme
3' end labeled
5' end labeled
Lane 1
Lane 2
Fig. 1 Summary of the data analysis procedures. a, A ‘line profile’
encompassing the middle ∼50% of a lane, defined in the 16-bit digital
image of an autoradiogram of a gel using ImageQuant (Molecular
Dynamics). Lane 2 is a region of the Tetrahymena ribozyme in CE buffer
containing 10 mM Na+. Lane 1 is ribozyme in CE buffer containing 10 mM
Na+ and 10 mM MgCl2. b, The density profile calculated using
ImageQuant and transferred to Origin 6.1 (OriginLab) equipped with
their Peak Fitting Module. c, Deconvolution of the data in (b) carried out
by fitting the peaks to a series of Lorentzian curves, taking care to ensure
that the global minimum was reached in each analysis. The numerical
peak as a function of nucleotide position were used to create the false
color image in Fig. 2. d, Schematic representation of the overlapping
regions of either 5′- or 3′-labeled ribozyme sequence that were individu-
ally analyzed using a sequential series of electrophoresis durations.
a
b
c
d
©2002NaturePublishingGrouphttp://www.nature.com/naturestructuralbiology](https://image.slidesharecdn.com/d7d7362e-fd72-404e-9d8d-53d9d8784b87-141201180628-conversion-gate02/75/Monovalent-Cations-Mediate-Formation-of-Native-Tertiary-Structure-of-Tetrahymena-Thermophila-Ribozyme-1-2048.jpg)
![letters
2 nature structural biology • advance online publication
also referenced to the OH• reactivity measured in CE buffer. The
ribozyme is annealed and folded in the presence of 10 mM Mg2+
under conditions that fully yield the native, catalytically active
conformation (data not shown). We have classified and mapped
the OH• protections and enhancements on a secondary/tertiary
structural ribozyme representation19 (Fig. 3).
Induction of native tertiary contacts
Increasing concentrations of Na+ induce formation of native ter-
tiary contacts in the Tetrahymena ribozyme in the absence of
Mg2+ (blue, Fig. 2)20–24. Regions of enhanced OH• reactivity
induced by Na+ are identical to those evident in the Mg2+-folded
native conformation (red, Fig. 2). The close similarity of the OH•
protection patterns in 1.5 M NaCl and 10 mM MgCl2 suggests
that monovalent cations mediate formation of many elements of
native structure. Examples of tertiary contacts that are formed in
1.5 M NaCl include the nucleotides of the tetraloop–receptor
interaction within P4–P6, the peripheral helix contact P13, the
proposed P9–P5 interaction, the OH• protections around P3 and
native catalytic core protections. Formation of P13 in buffer
containing only NaCl has also been observed by dimethyl sulfate
(DMS) footprinting10. Nucleotides 265–269 are not protected in
10 mM Mg2+ but are at high [Na+]. Thus, monovalent cations
can also induce formation of non-native tertiary structure. DMS
modification studies have shown the formation of the alt-P3
alternative secondary structures at <100 mM Na+, with P3 pre-
sent at >100 mM Na+ (ref. 10). At [Na+] >300 mM, we infer that
P3 secondary structure is present on the basis of the similarities
of the OH• reactivity pattern surrounding P3 with that observed
at 10 mM Mg2+.
Divalent ion-specific tertiary structure
Despite the overall similarity of Na+- and Mg2+-folded RNA, not
all of the native tertiary interactions are formed in the presence
of 1.5 M Na+. Protection is absent for nucleotides 302–306 (J8/7)
and 99–100 (P3) within the catalytic core and nucleotides
167–173 that constitute one-half of the P14 tertiary interaction.
(Because our data does not report on the other side of the P14
contact in P2, the absence of P14 is inferred.) Protection is
absent within P5c and is barely evident within the A-bulge at
1.5 M Na+ (Figs 2, 3). Thus, Na+ mimics some, but not all, of the
structural roles played by Mg2+.
Bound Mg2+ ions are observed in the crystal structure of the
isolated P4–P6 domain within P5c and the A-bulge20,21,25,26. Urea
denaturation-hydroxyl radical footprinting analysis of the iso-
lated Mg2+-folded P4–P6 domain shows that the P5c and
A-bulge protections have elevated stability compared with those
in the rest of the domain12. Phosphothioate substitution27 and
bound Fe2+-mediated OH• cleavage studies28 also suggest specifi-
cally bound divalent cations in this region. Finally, a Mg2+-
dependent secondary structure change has been observed for
isolated P5abc by NMR26,29. Together, these results are consistent
with divalent ion-specific stabilization of structure in this
region. The absence of native contacts in the ribozyme core at
high monovalent ion concentrations in the absence of divalent
cation is consistent with the absence of detectable catalytic activ-
ity under these solution conditions (data not shown).
Induction of native tertiary ‘exposures’
Nucleotide-specific enhancements of OH• reactivity are also
observed relative to low-salt reference condition (red, Fig. 2).
Fig. 2 A false-color map of the changes in OH• reactivity for the Tetrahymena ribozyme observed as a function of Na+ concentration. The OH• reac-
tivity is referenced to the lowest salt concentration analyzed, CE buffer (green). Decreased OH• reactivity (‘protection’) is blue and increased reactiv-
ity (‘enhancement’) is red, as shown in the insert to the figure. The bar beneath the Na+ titration shows the ribozyme annealed in CE buffer with
10 mM MgCl2 added during annealing of the RNA, which is represented using the same color coding. Data were collected in CE buffer (∼10 mM Na+)
and 40, 80, 160, 400, 600, 750 and 1,500 mM Na+ by the addition of NaCl. The data underlying the graph were interpolated and smoothed to
improve its visualization.
©2002NaturePublishingGrouphttp://www.nature.com/naturestructuralbiology](https://image.slidesharecdn.com/d7d7362e-fd72-404e-9d8d-53d9d8784b87-141201180628-conversion-gate02/75/Monovalent-Cations-Mediate-Formation-of-Native-Tertiary-Structure-of-Tetrahymena-Thermophila-Ribozyme-2-2048.jpg)
![letters
nature structural biology • advance online publication 3
These increases in OH• reactivity are typically observed in loops
on the outside of the native, Mg2+-folded structure and, thus, are
highly solvent accessible24 (Fig. 3). If the ‘unfolded’ RNA at low
salt is viewed as an ensemble of structures, it is plausible to view
these regions as fluctuating between ‘inside’ and ‘outside’ posi-
tions. In the presence of cations, they become progressively
locked outside as the ensemble of partially folded structures con-
verges towards the unique native conformation.
Na+-linking folding isotherms
The false-color mapping (Fig. 2) clearly shows the Na+ concen-
tration dependence of native tertiary structure formation. Each
of the tertiary contacts can also be represented by a folding
isotherm relating the fractional saturation of the protection to
[Na+] (Fig. 4). The Na+-dependent folding isotherms differ dra-
matically from the published Mg2+-folding isotherms whose
equilibrium constants are tightly clustered and moderately
(P4–P6) or highly (the catalytic core) concerted15,16. To capture
both the concentration dependence and concerted nature of the
Na+-folding isotherms in a single measure, the ion concentration
at which they achieve 20% saturation is used to classify the data
and color coding (Fig. 3) to visualize the onset of the transitions.
The behavior of the Na+-dependent folding isotherms
obtained by OH• footprinting shows changes that can be
grouped within four [Na+] ranges (Figs 2–4). (i) At 30–40 mM
Na+, protection to 20% saturation of a proposed contact
between P9 and P5 is evident as is the tetraloop–receptor within
P4–P6 (green, Figs 3, 4a). Enhanced reactivity of nucleotides
381–382 within P9.2 is also observed. (ii) The isotherms for the
P9.1a–P2.1 contact of the peripheral helices (P13) and core ter-
tiary contacts reach 20% saturation at 150–300 mM Na+ (cyan,
Figs 3, 4b). (iii) A large number of isotherms for protections
within P4–P6 and the catalytic core reach 20% saturation by
750 mM Na+ (blue, Fig. 3). (iv) Additional native protections
(including the A-bulge) show the onset of protection at 1.5 M
Na+ but do not fully saturate in our experiments (purple, Figs 3,
4c). In aggregate, a high proportion of native tertiary contacts
showing robust protections are observed in 1.5 M Na+ in the
absence of divalent cations.
Some Na+-linked folding isotherms display characteristics not
seen in Mg2+-induced folding. For example, OH• protection
appears at intermediate concentrations of Na+ only to vanish as
the monovalent ion concentration increases15. Such transient
protections pertaining to [Na+] are observed in P2.1
(nucleotides 61–62, 64–65 and 85–86; Figs 2, 4d). The latter two
protections are not observed in the presence of Mg2+ and, thus,
are presumed to be non-native.
Na+-linked folding isotherms are biphasic at several other
sites, including the spatially close protections at the J8/7 side of
P8 (nucleotides 298–300) and J2.1/3 (nucleotides 93–96) and
nucleotides 202–203 in a P5–P9 contact. The initial phase of
these biphasic transitions may represent non-native contacts
formed in the electrostatic collapse of these regions, followed by
rearrangement to the native conformation at higher concentra-
tions of Na+. Thus, titrating the ribozyme with Na+ does not
result in a monotonic march towards a ‘native-like’ ensemble of
structures. Rather, each salt concentration is a unique mix of
native and non-native tertiary interactions.
Fig. 3 Structural mapping and classification of the OH• reactivity changes. The color-coding of this figure is based upon the concentration of Na+ at
which either the protection or enhancement reaches 20% of the maximum change observed as a function of the Na+- against the Mg2+-folded value.
(The 20% saturation value was chosen to capture the contribution of both the midpoint and slope of the isotherms in a single value.) Sites repre-
sented by two colors indicate the presence of biphasic transitions in the isotherms (for example, nucleotides 93–96). Open rectangles with two col-
ors denote transient transitions that appear at the lower concentration and disappear at higher concentrations of Na+ (for example, nucleotides
65–67). The regions surrounded by black with colored letters denote nucleotides characterized by greater protection in Na+ than in Mg2+ (for exam-
ple, nucleotides 265–269). The arrows show several long-range interactions present in the native folded ribozyme.
©2002NaturePublishingGrouphttp://www.nature.com/naturestructuralbiology](https://image.slidesharecdn.com/d7d7362e-fd72-404e-9d8d-53d9d8784b87-141201180628-conversion-gate02/75/Monovalent-Cations-Mediate-Formation-of-Native-Tertiary-Structure-of-Tetrahymena-Thermophila-Ribozyme-3-2048.jpg)
![letters
4 nature structural biology • advance online publication
Several conclusions can be drawn from these data. First, the
monovalent cation-induced electrostatic collapse of the
ribozyme3,7,11 includes formation of both native and non-native
tertiary contacts. Second, this process does not proceed in a
strictly monotonic fashion. Structural rearrangements from
non-native to native tertiary contacts are observed. Third, non-
native tertiary contacts present at low concentrations of Na+ may
be a source of the kinetic traps that retard Mg2+-induced folding
under this solution condition16. Because these kinetic traps may
be released in the ensemble of molecules that comprise the ‘ini-
tial state’, folding of the RNA may be accelerated in their
absence11,12,30.
Global and local tertiary correlations
Small angle X-ray scattering (SAXS)3,10, analytical ultracentrifu-
gation11 and gel electrophoresis7 studies have demonstrated
compaction of the Tetrahymena ribozyme with increasing con-
centrations of monovalent cations. An analytical ultracentrifu-
gation analysis of the L-21 ribozyme was carried out over the
range of Na+ concentrations probed by OH• footprinting to cor-
relate the formation of local tertiary contacts with changes in the
global conformation.
Sedimentation (S20,w) and diffusion coefficients (D20,w) are
determined from a sedimentation velocity experiment. From
these quantities, the Stokes radius (RH) and the axial ratio (a / b)
were calculated for the ribozyme as a function of [Na+] or
[Mg2+]. With divalent cations, compaction of the ribozyme
occurs coincidently with a decrease in the axial ratio, predomi-
nantly as a single concerted step with a midpoint of ∼0.2 mM
MgCl2. A second transition of minimal amplitude is observed
with a midpoint of 7.5–8.9 mM MgCl2 (open circles, Fig. 5).
Similarly, two transitions are evident in the Na+ titrations. The
midpoint of the predominant transition is at 39 mM, with the
second transition at 587 mM (solid squares, Fig. 5). The
Na+-mediated transitions are much less concerted than the
Mg2+-mediated transition. Together, the changes in RH and a / b
show that the ribozyme does not uniformly condense upon the
addition of Na+. The ultracentrifugation results are in overall
agreement with small angle X-ray scattering studies conducted
over a comparable range of [Na+]3,10,31 (R. Das, I. Millett,
R. Russell, S. Doniach and D. Herschlag, pers. comm.).
A model of monovalent ion-induced folding
The global conformational changes together with the direct
reports of local solvent accessibility provided by OH• footprint-
ing allow a model for the Na+-induced compaction of the
Tetrahymena ribozyme to be developed. The appearance of the
P5–P9 and tetraloop–receptor protections at low concentrations
of Na+ (green, Fig. 3) suggests that the initial compact conforma-
tion is stabilized by the formation of specific, native tertiary con-
tacts. The presence of the non-native P13 protection as well as
the appearance of the transitions that are transient with [Na+] at
the intersection of P2.1–P3 and P8–J8/7 suggests that specific
non-native interactions also stabilize this compact conforma-
tion. The formation of the P5–P9 tertiary contact dramatically
limits both the size and length of the ensemble of RNA molecules
and, thus, can rationalize the initial compaction observed by
ultracentrifugation and SAXS.
Further increases in [Na+] from 300 to 750 mM results in pro-
gressive organization of P4–P6, the catalytic core and the periph-
eral helices. Formation of these native tertiary contacts correlates
with the second global compaction transition observed by ultra-
centrifugation (Fig. 5). Additional increases in the [Na+] to
1.5 M seem to ‘tighten-up’ the ribozyme structure. Overall, these
results suggest that Na+ drives formation of an ensemble of com-
pact and spatially constrained structures stabilized by both
native and non-native contacts that converges to more native-
like structures with increasing [Na+]. This absence of several
local tertiary contacts between ribozyme folded with Na+ com-
pared with Mg2+ is reflected in the difference in global confor-
mation evident at high [Na+] and [Mg2+] (Fig. 5).
Implications for the kinetics of folding
The time-dependence of both tertiary contact formation and
compaction of the Tetrahymena ribozyme upon Mg2+-initiated
folding have been measured at low [Na+]; tertiary structure
forms at least five times more slowly than global compaction
under this experimental condition17,31. Thus, the search for
Fig. 4 Examples of the Na+-dependent folding isotherms derived from
the data summarized in the false-color map of Fig. 2. Each isotherm rep-
resents the averaged protection for the indicated nucleotides as a func-
tion of Na+ concentration (Fig. 2). a, For the P4–P6 tetraloop, Na+
20% =
103 mM and nH = 1.2; b, core region, Na+
20% = 131 mM and nH = 0.9;
c, A-Bulge, Na+
20% = 829 mM and nH = 4.0; and d, the transient transition
at P2.1, Na+
20% = 22 mM and 333 mM, and nH = 2.0 and 3.8.
a
b
c
d
©2002NaturePublishingGrouphttp://www.nature.com/naturestructuralbiology](https://image.slidesharecdn.com/d7d7362e-fd72-404e-9d8d-53d9d8784b87-141201180628-conversion-gate02/75/Monovalent-Cations-Mediate-Formation-of-Native-Tertiary-Structure-of-Tetrahymena-Thermophila-Ribozyme-4-2048.jpg)
![letters
nature structural biology • advance online publication 5
contact partners occurs within a constrained conformational
space.
Monovalent cations not only condense RNA4 but also alter the
predominant pathway of its folding7,10. Compaction per se can be
argued not to facilitate fast and error-free folding but, rather, the
elimination of non-native contacts and/or the formation of
native contacts in the ensemble of RNA molecules is responsible.
The present data argue for contributions from both mechanisms
and suggest that the time lag of stable tertiary structure forma-
tion behind global compaction will diminish as the monovalent
cation concentration increases.
Substantial acceleration of the early steps of folding for both
the isolated P4–P6 domain9,11,30 and the L-21 ribozyme
(T. Uchida, Q.H., C.Y.R., M.R.C. and M.B., submitted) is
induced by increases in the Na+ concentration from ∼10 to
200 mM. The formation of native tertiary contacts and the
destabilization of non-native tertiary contacts occur over this
[Na+] (Fig. 2). A prediction of these data is that a progressively
smaller subset of Mg2+ folding transitions will be observed as
[Na+] is increased.
For the catalytic core, numerous short- and long-range inter-
actions present at high Na+ concentrations will restrain the
ensemble to a small number of native-like populations. The P3
rather than alt-P3 secondary structure is dominant above
300 mM NaCl10, eliminating a well characterized kinetic trap32,33.
These structures and contacts are predicted to favor direct fold-
ing. Although the above-described interactions should facilitate
folding of the catalytic core, the structure surrounding J8/7 is
non-native in the absence of Mg2+ and P1 is apparently not
docked to J9/7. Thus, the contribution of kinetic traps to Mg2+-
dependent folding of the catalytic core cannot be discounted even
when a large fraction of the native tertiary structure is present.
In summary, the ensemble of structures present at high con-
centrations of monovalent cation are predicted to be committed
to fast and correct folding because of the high probability of for-
mation of native tertiary contacts. Thus, increases in monova-
lent ion concentration induce structural changes that cause
folding to commence, not only from different locations along
the edge of folding funnel, but also from deep within it.
However, Na+ cannot effectively substitute for all Mg2+ ions;
Mg2+ is required for the formation of key catalytic core interac-
tions that allow catalysis to occur.
Methods
Equilibrium titrations. The L-21 Sca I ribozyme from
Tetrahymena thermophila was transcribed by T7 polymerase and 5′
or 3′ end labeled with 32P as described34–36. The labeled RNA in CE
buffer was heated to 95 °C for 2 min and annealed by slow cooling
to room temperature. Aliquots of the RNA were equilibrated at
42 °C in CE buffer to which the indicated concentration of NaCl
was added. Fe-EDTA-mediated Fenton chemistry was used to gen-
erate OH• (refs 18,37–39). The RNA samples were footprinted at
42 ºC for 2 min and processed for electrophoresis as described16.
Following electrophoresis, the gel was imaged using a phosphor
storage plate that was scanned by a Storm 800 PhosphorImager
(Molecular Dynamics).
Analytical ultracentrifugation. Sedimentation velocity experi-
ments were performed at 20 °C using a Beckman Optima XL-I ana-
lytical ultracentrifuge, An-60 Ti rotor and Al-Epon/Charcoal
centerpieces at 30,000g. The Tetrahymena ribozyme at 71.4 nM
(8.96 µg ml–1) was folded under the same conditions as described
above with indicated concentrations of either NaCl or MgCl2. The
sedimentation boundaries were directly fit as dc / R using SVED-
BERG40 to determine the sedimentation and diffusion coefficients, S
and D, respectively. The observed values were normalized to stan-
dard conditions at 20 °C in water. The Stokes radius, RH, and axial
ratio, a/b, were calculated using Sednterp41,42 assuming υ = 0.53 cm3
gm–1 and hydration = 0.59.
Footprinting data analysis. The relative densities of the bands
representing each reaction product were determined by single
band analysis following a published protocol43. The gel images are
converted to lane profiles using ImageQuant (Molecular Dynamics)
that are fit to a series of Lorentzian curves using the Peak Fitting
Module within Origin v6.1 (OriginLab). Iterations are run until the
fidelity of fitted peak profile reaches at least 99.9% of original pro-
file. The fitted peak areas are assembled as a two-dimensional
matrix. Each column (lane) is ‘standardized’ by dividing the peak
area values by single or averaged ‘standard’ bands16,43–45. (Standard
bands on a gel electrophoretogram do not vary as a function of the
experimental parameter being probed in a series of footprinting
reactions and correct for variation in the amount of sample loaded
onto each lane of a gel.)
The preceding published protocols have been automated by
analysis software that determines and uses the most appropriate
reference bands. The principle underlying the automated system is
that the standard deviation of the peak area of valid standard
bands is minimal compared with bands whose peak area varies with
the experimental parameter being assayed. The algorithm sequen-
tially calculates the standard deviation for a set of bands and then
ranks them by a ‘score’ that is equal to the ‘global standard devia-
tion of matrix’ multiplied by the ‘mean value of standard deviations
of matrix row element’. The standardized matrix is then ‘normal-
ized’45 to the reference lane with the lowest salt concentration by
Fig. 5 Values of the Stokes radius and axial ratio (RH and a / b, respective-
ly) derived from sedimentation velocity experiments conducted in CE
buffer to which the indicated [NaCl] or [MgCl2] was added. The transi-
tions were modeled as the sum of two Hill equilibria. For the MgCl2 titra-
tion (open circles), the midpoints of these transitions are 0.2 and 7.5 mM
for RH and 0.2 and 8.9 mM for a / b. For the NaCl titration (solid squares),
the midpoints of these transitions are 39 mM and 587 mM for RH and
28 and 582 mM for a / b.
©2002NaturePublishingGrouphttp://www.nature.com/naturestructuralbiology](https://image.slidesharecdn.com/d7d7362e-fd72-404e-9d8d-53d9d8784b87-141201180628-conversion-gate02/75/Monovalent-Cations-Mediate-Formation-of-Native-Tertiary-Structure-of-Tetrahymena-Thermophila-Ribozyme-5-2048.jpg)
![letters
6 nature structural biology • advance online publication
division. The details of these procedures and the software devel-
oped to streamline these procedures will be described elsewhere.
Sets of individual analyses were averaged and combined to yield
the two-dimensional map of the entire ribozyme using KyPlot v2.0β
(K. Yoshioka, Japan).
The Na+-dependent folding isotherms were generated using a
summation over protection values from the single band analysis
(Fig. 2). The integrated density, ρi, as a function of [Na+] was scaled
to the fractional saturation of the protection (ρi – ρlower) / (ρupper –
ρlower), where ρlower and ρupper are the lower and upper limits of the
transition curves determined from the CE buffer (‘unfolded’) and
Mg2+-CE buffer (native) lanes, respectively16,44,45. The isotherms were
fit by single or multiple-transition Hill equations46.
Acknowledgments
We thank D. Herschlag and R. Russell for pre-publication discussion of their
results. This work was supported by grants from the National Institute of General
Medical Sciences and the Biomedical Technology Program of the Division of
Research Resources.
Competing interests statement
The authors declare that they have no competing financial interests.
Correspondence should be addressed to M.B. email: brenowit@aecom.yu.edu
Received 21 May, 2002; accepted 16 October, 2002.
1. Piccirilli, J.A., Vyle, J.S., Caruthers, M.H. & Cech, T.R. Nature 361, 85–88 (1993).
2. Downs, W.D. & Cech, T.R. RNA 2, 718–732 (1996).
3. Russell, R., Millett, I.S., Doniach, S. & Herschlag, D. Nature Struct. Biol. 7, 367–370
(2000).
4. Buchmueller, K.L., Webb, A.E., Richardson, D.A. & Weeks, K.M. Nature Struct.
Biol. 7, 362–366 (2000).
5. Fang, X. et al. Biochemistry 39, 11107–11113 (2000).
6. Heilman-Miller, S.L., Pan, J., Thirumalai, D. & Woodson, S.A. J. Mol. Biol. 309,
57–68 (2001).
7. Heilman-Miller, S.L., Thirumalai, D. & Woodson, S.A. J. Mol. Biol. 306, 1157–1166
(2001).
8. Russell, R. & Herschlag, D. J. Mol. Biol. 291, 1155–1167 (1999).
9. Silverman, S.K., Deras, M.L., Woodson, S.A., Scaringe, S.A. & Cech, T.R.
Biochemistry 39, 12465–12475 (2000).
10. Russell, R. et al. Proc. Natl. Acad. Sci. USA 99, 155–160 (2002).
11. Deras, M.L., Brenowitz, M., Ralston, C.Y., Chance, M.R. & Woodson, S.A.
Biochemistry 39, 10975–10985 (2000).
12. Uchida, T., He, Q., Ralston, C.Y., Brenowitz, M. & Chance, M.R. Biochemistry 41,
5799–5806 (2002).
13. Strahs, D. & Brenowitz, M. J. Mol. Biol. 244, 494–510 (1994).
14. Shadle, S.E. et al. Nucleic Acids Res. 25, 850–860 (1997).
15. Celander, D.W. & Cech, T.R. Science 251, 401–407 (1991).
16. Sclavi, B., Woodson, S., Sullivan, M., Chance, M.R. & Brenowitz, M. J. Mol. Biol.
266, 144–159 (1997).
17. Sclavi, B., Sullivan, M., Chance, M.R., Brenowitz, M. & Woodson, S.A. Science 279,
1940–1943 (1998).
18. Ralston, C.Y., He, Q., Brenowitz, M. & Chance, M.R. Nature Struct. Biol. 7,
371–374 (2000).
19. Cech, T.R., Damberger, S.H. & Gutell, R.R. Nature Struct. Biol. 1, 273–280 (1994).
20. Cate, J.H., Hanna, R.L. & Doudna, J.A. Nature Struct. Biol. 4, 553–558 (1997).
21. Cate, J.H. et al. Science 273, 1678–1685 (1996).
22. Cate, J.H. et al. Science 273, 1696–1699 (1996).
23. Golden, B.L., Gooding, A.R., Podell, E.R. & Cech, T.R. Science 282, 259–264 (1998).
24. Lehnert, V., Jaeger, L., Michel, F. & Westhof, E. Chem. Biol. 3, 993–1009 (1996).
25. Cate, J.H. & Doudna, J.A. Structure 4, 1221–1229 (1996).
26. Zheng, M., Wu, M. & Tinoco, I. Jr Proc. Natl. Acad. Sci. USA 98, 3695–3700 (2001).
27. Christian, E.L. & Yarus, M. Biochemistry 32, 4475–4480 (1993).
28. Berens, C., Streicher, B., Schroeder, R. & Hillen, W. Chem. Biol. 5, 163–175 (1998).
29. Wu, M. & Tinoco, I. Jr Proc. Natl. Acad. Sci. USA 95, 11555–11560 (1998).
30. Silverman, S.K. & Cech, T.R. RNA 7, 161–166 (2001).
31. Russell, R. et al. Proc. Natl. Acad. Sci. USA 99, 4266–4271 (2002).
32. Pan, J. & Woodson, S.A. J. Mol. Biol. 280, 597–609 (1998).
33. Pan, J., Deras, M.L. & Woodson, S.A. J. Mol. Biol. 296, 133–144 (2000).
34. Zaug, A.J., Grosshans, C.A. & Cech, T.R. Biochemistry 27, 8924–8931 (1988).
35. Lingner, J. & Keller, W. Nucleic Acids Res. 21, 2917–2920 (1993).
36. Latham, J.A., Zaug, A.J. & Cech, T.R. Methods Enzymol. 181, 558–569 (1990).
37. Tullius, T.D., Dombroski, B.A., Churchill, M.E. & Kam, L. Methods Enzymol. 155,
537–558 (1987).
38. Wang, X.D. & Padgett, R.A. Proc. Natl. Acad. Sci. USA 86, 7795–7799 (1989).
39. Emerick, V.L. & Woodson, S.A. Proc. Natl. Acad. Sci. USA 91, 9675–9679 (1994).
40. Philo, J.S. Biophys. J. 72, 435–444 (1997).
41. Hinz, H.-J. Thermodynamic Data for Biochemistry and Biotechnology (Springer-
Verlag, New York; 1986).
42. Glasel, J.A. & Deutscher, M.P. Introduction to Biophysical Methods for Protein
and Nucleic Acid Research (Academic Press, San Diego; 1995).
43. Pastor, N., Weinstein, H., Jamison, E. & Brenowitz, M. J. Mol. Biol. 304, 55–68
(2000).
44. Brenowitz, M., Senear, D.F., Shea, M.A. & Ackers, G.K. Methods Enzymol. 130,
132–181 (1986).
45. Sclavi, B., Woodson, S., Sullivan, M., Chance, M. & Brenowitz, M. Methods
Enzymol. 295, 379–402 (1998).
46. Hill, A.V. J. Physiol. (London) 40, iv–vii (1910).
©2002NaturePublishingGrouphttp://www.nature.com/naturestructuralbiology](https://image.slidesharecdn.com/d7d7362e-fd72-404e-9d8d-53d9d8784b87-141201180628-conversion-gate02/75/Monovalent-Cations-Mediate-Formation-of-Native-Tertiary-Structure-of-Tetrahymena-Thermophila-Ribozyme-6-2048.jpg)







![Spectral studies of 5-({4-amino-2-[(Z)-(2-hydroxybenzylidene) amino] pyrimidi...](https://cdn.slidesharecdn.com/ss_thumbnails/e0662835-150425014612-conversion-gate02-thumbnail.jpg?width=640&height=640&fit=bounds)