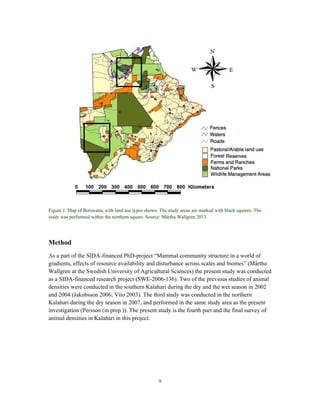
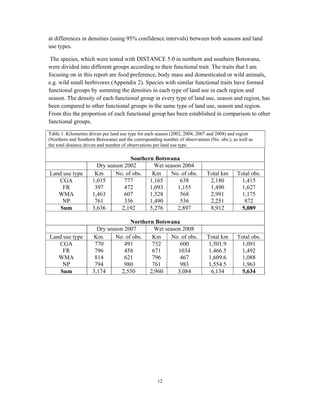
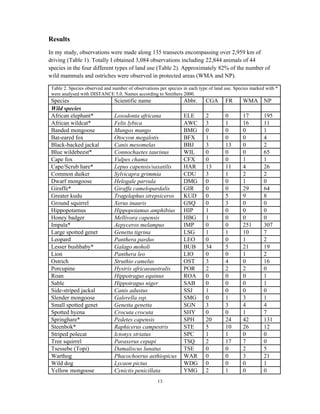
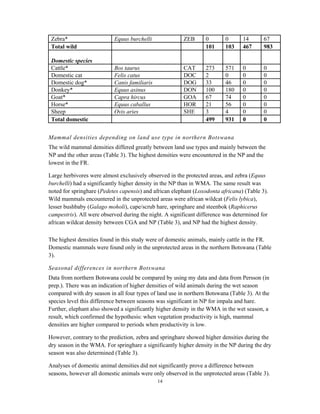
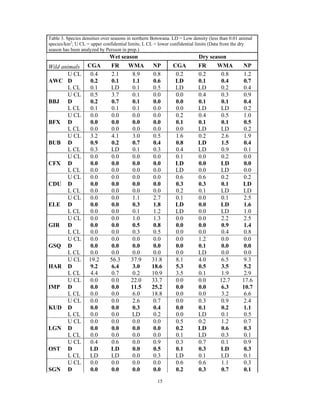
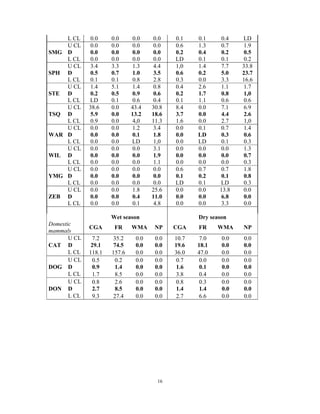
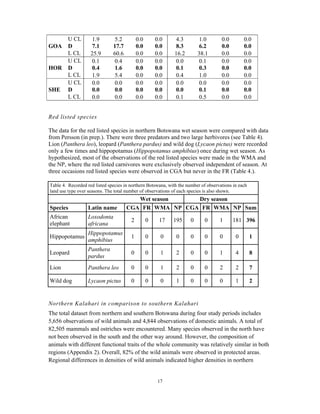
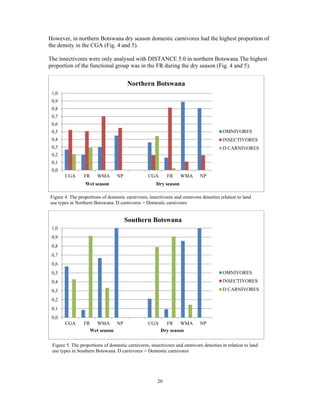
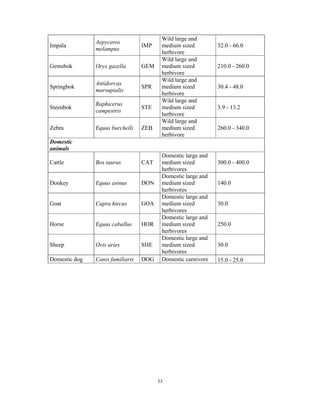
This document summarizes a study on mammal densities in the Kalahari region of Botswana and how they are impacted by seasons and land use. The study compares mammal densities across four different land use areas - Communal Grazing Areas, Fenced Ranches, Wildlife Management Areas, and National Parks. Previous studies have shown that human activities like livestock grazing can negatively impact wildlife by reducing resources and fragmenting the landscape. The goal of this study was to investigate how spatial and temporal variations in mammal densities relate to different types of human land use. Large and medium herbivores and carnivores were found to be most affected by human activities, while smaller mammals were least affected. Seasonal and regional environmental differences had less impact than the