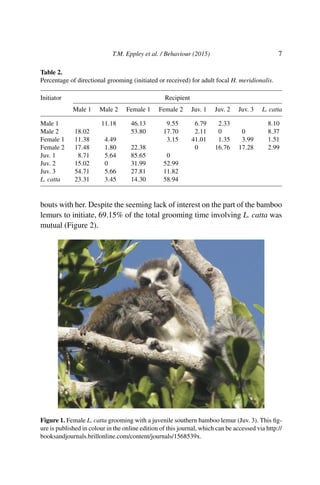
This document describes an unusual case of association between a female ring-tailed lemur (Lemur catta) and a group of southern bamboo lemurs (Hapalemur meridionalis) in southeast Madagascar. The female L. catta was observed feeding, travelling, and interacting affiliatively with the H. meridionalis group over a prolonged period. Both species exhibited behaviors like vocal coordination, dietary overlap, and the L. catta was observed grooming and caring for the H. meridionalis infant. This flexible behavior allowed the successful integration of the female ring-tailed lemur into the bamboo lemur social group.



















































![The red ruffed lemur[1]](https://cdn.slidesharecdn.com/ss_thumbnails/theredruffedlemur1-110204140855-phpapp02-thumbnail.jpg?width=640&height=640&fit=bounds)












