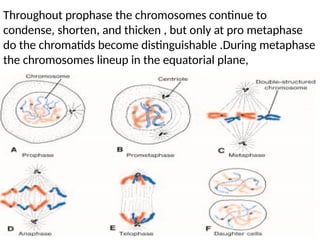
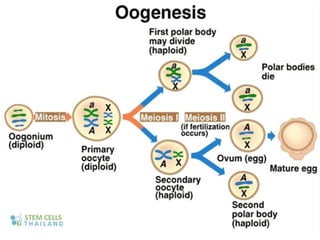
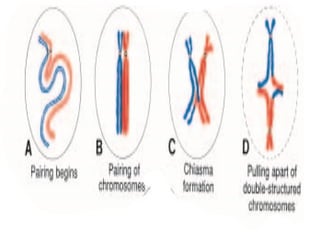
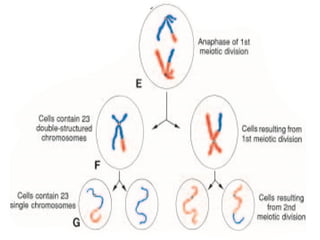
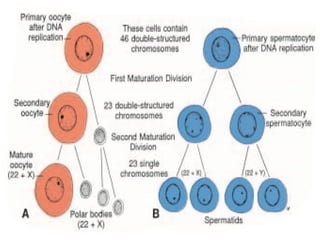
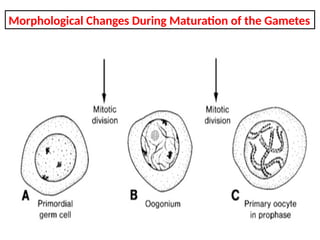
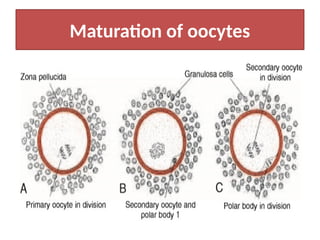
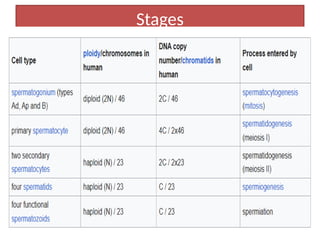
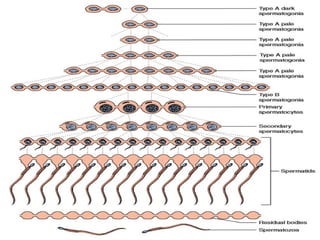
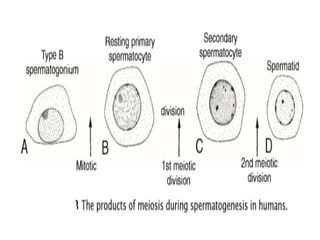
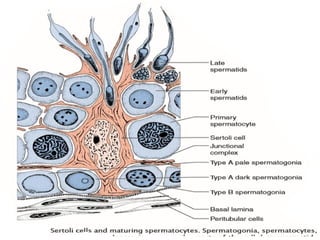
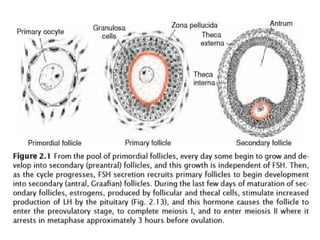
The document outlines the developmental anatomy of primordial germ cells and their role in gametogenesis, including the processes of mitosis and meiosis as they develop into male and female gametes. It details the chromosome theory of inheritance and the hormonal regulation of the ovarian and spermatogenic cycles, emphasizing the changes that occur during the maturation of oocytes and spermatozoa. Fertilization is described as a multi-phase process involving capacitation and the acrosome reaction, ultimately leading to the fusion of male and female gametes.
































































































![CTEV [ clubfoot] DR ARUN LAL ,DR MOHAMED ASHRAF travancore medical college k...](https://cdn.slidesharecdn.com/ss_thumbnails/ctevclubfootdrarunlaldrmohamedashraftravancoremedicalcollegekollamkeralaindia-260208063247-18fc466c-thumbnail.jpg?width=640&height=640&fit=bounds)


![ONFH[AVN HIP] -TRIPLE REGIME -A NOVAL SURGICAL CONCEPT .pptx](https://cdn.slidesharecdn.com/ss_thumbnails/onfhavnhip2026koaconcalicutdrgokuldevdrmashraf-260210064517-213ec005-thumbnail.jpg?width=640&height=640&fit=bounds)







