Downloaded 82 times










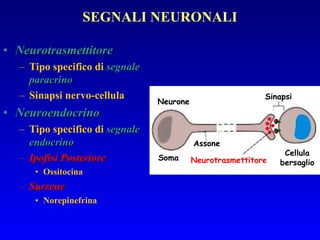





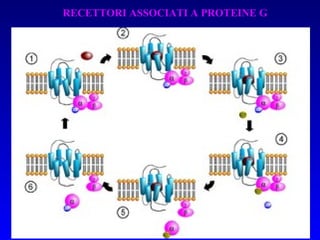
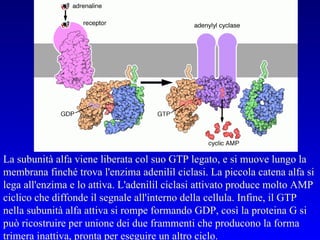
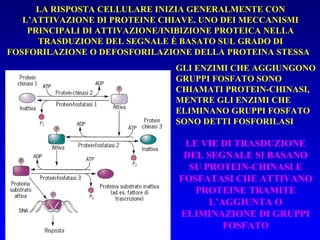




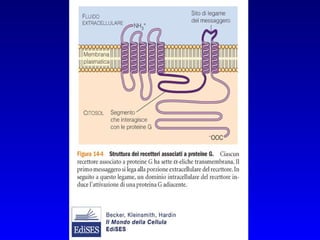
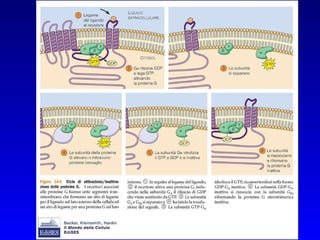

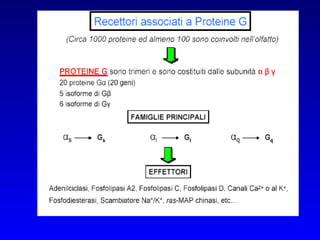


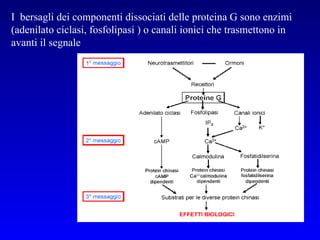
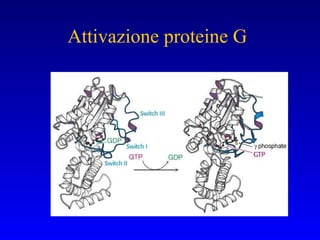
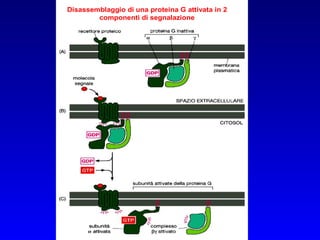



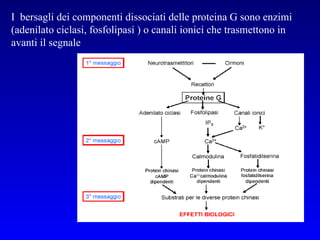
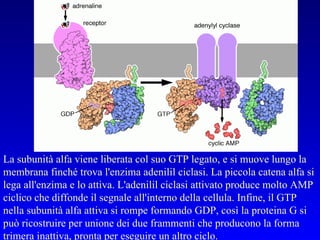
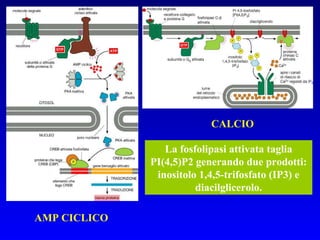
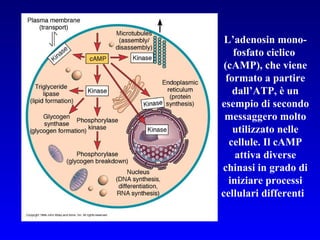
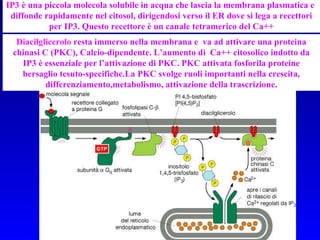
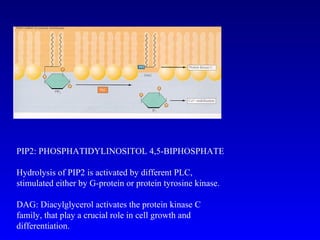



![PLC DAG PKC Calcio – il messaggero universale L’IP 3 si lega al recettore per l’IP 3 sul reticolo endoplasmatico e apre un canale del Ca 2+ (che fa parte del recettore) L’attivazione della PLC porta alla formazione di IP 3 solubile in acqua Il Ca 2+ rilasciato dal RE si lega alle Calmoduline permettendole di interagire con altre proteine e attivarle La Calmodulina può attivare pompe del Ca 2+ del reticolo endoplasmatico abbassando la [Ca 2+ ] citosolico La Calmodulina può attivare pompe del Ca 2+ sulla membrana plasmatica abbassando la [Ca 2+ ] citosolico La Calmodulina attiva una vasta gamma di proteine p.es chinasi calmodulina-dipendenti kinasi](https://image.slidesharecdn.com/comunicazionecellulare-110603133748-phpapp02/85/Comunicazione-cellulare-66-320.jpg)



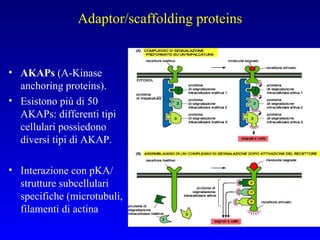
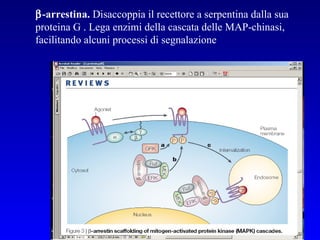



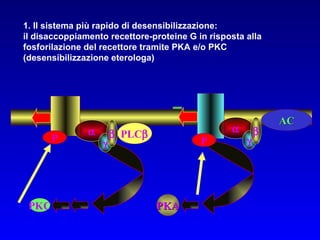
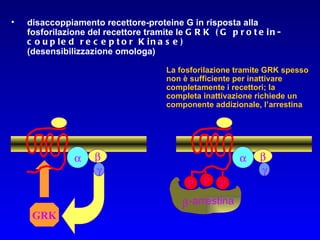


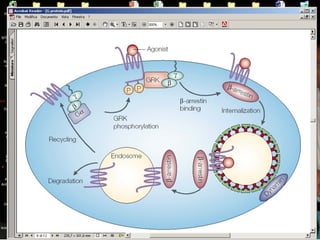
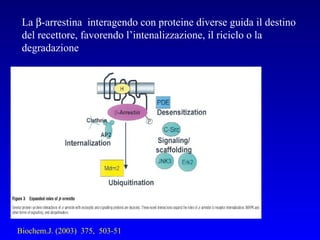



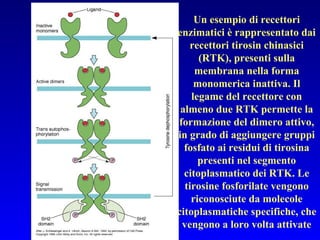
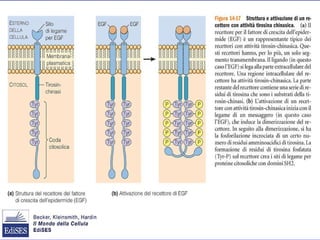
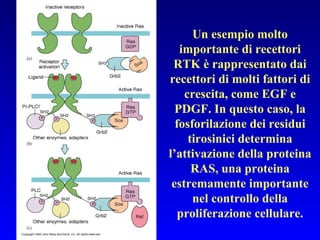
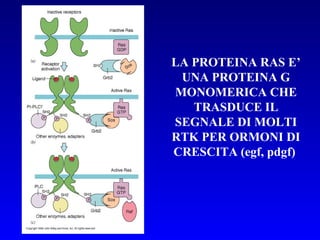
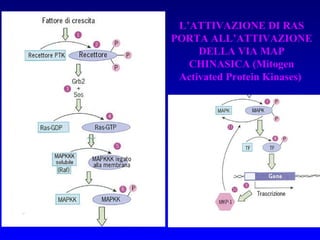
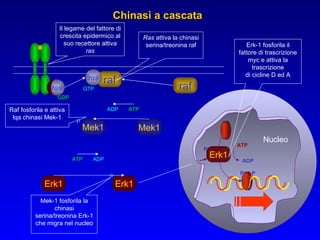
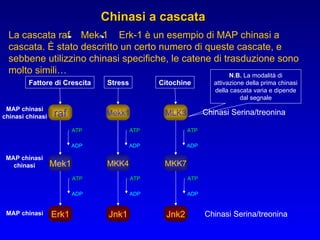
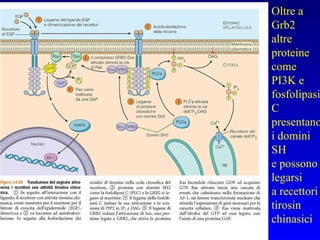
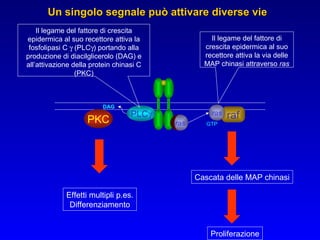
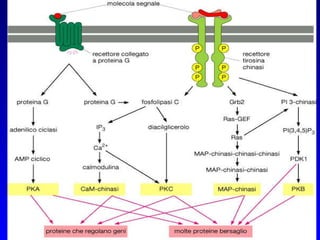
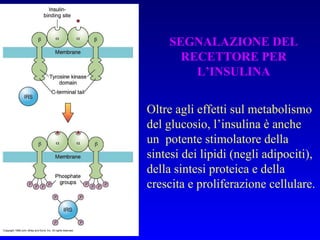
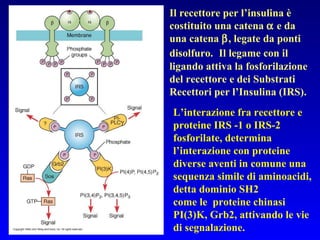
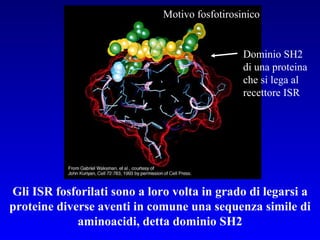

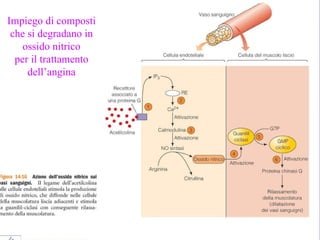


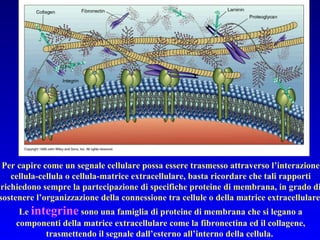
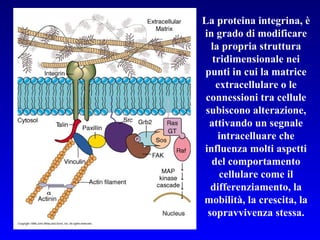
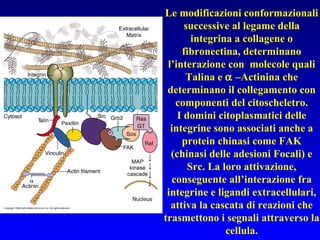
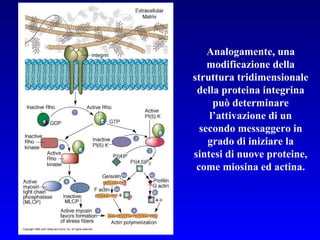

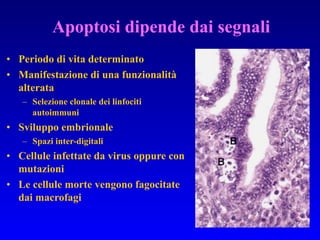
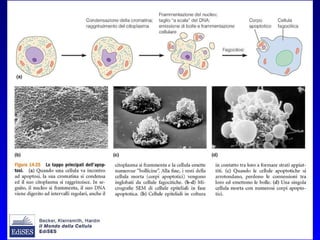


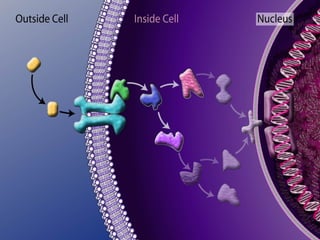
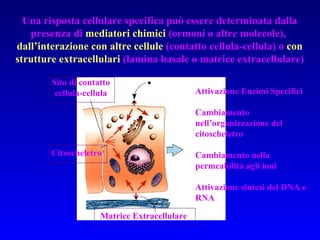
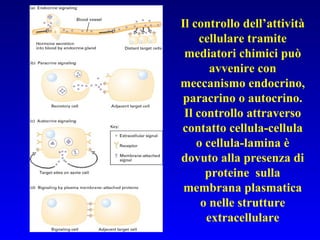
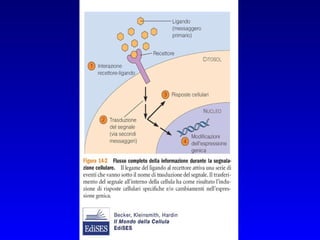
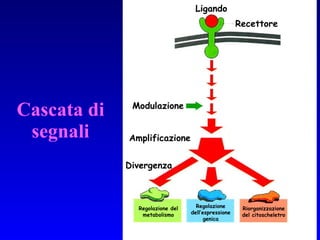
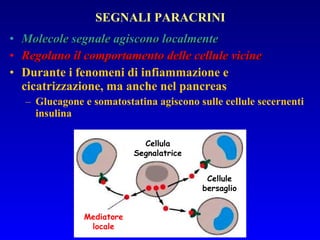
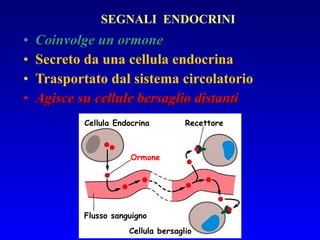
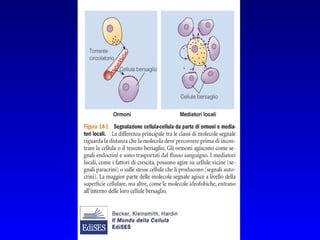
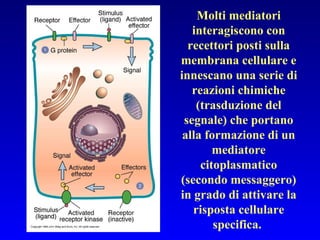
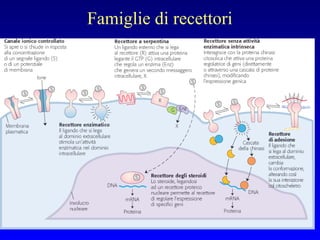
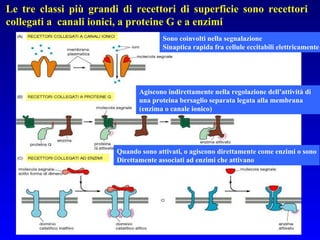
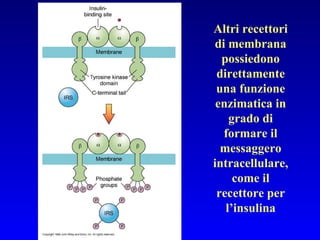
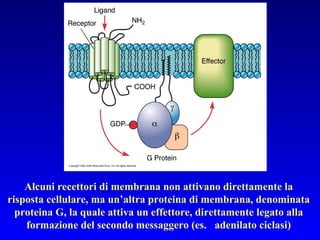
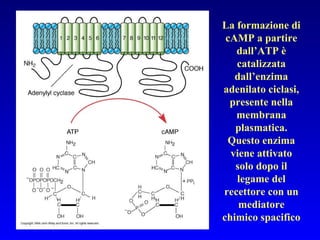
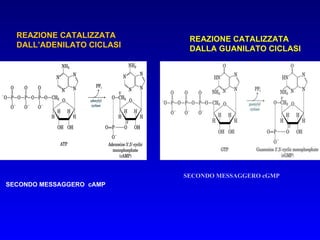
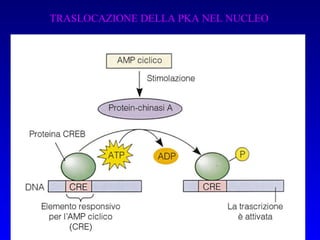
Il documento tratta della segnalazione cellulare e della trasduzione del segnale, evidenziando come le cellule possano adattarsi a cambiamenti ambientali grazie a meccanismi molecolari. Descrive i diversi tipi di segnali e recettori, spiegando il funzionamento delle vie di segnalazione intracellulari e l'importanza di messaggeri secondari come cAMP e calcio. Infine, sottolinea le conseguenze di alterazioni nei processi di segnalazione, come il possibile sviluppo di tumori.




























































