The document discusses receptor-mediated activation of nitric oxide (NO) synthesis by arginine in the vasculature. It first provides historical background on the discovery of NO and its role in vasodilation. It then presents evidence that arginine can activate NO synthesis through receptor-mediated mechanisms involving imidazoline/alpha-2 adrenergic receptors, G-proteins, and intracellular calcium signaling. Specifically, it shows that arginine and the endogenous agonist agmatine can directly stimulate NO production in endothelial cells, and this effect is blocked by receptor and G-protein antagonists. It also demonstrates that arginine and agonists elicit vasodilation through receptor-dependent mechanisms. This work helps





![Arginine Paradox
Intracellular [Arginine] = 0.8-2.0 mM
Plasma [Arginine] = 100-150 µM
Km of Arginine for cNOS = 2.9 µM
Exogenous Arginine is beneficial in endothelial-dependent vasodilation.
• Intravenous infusion of L-arginine in healthy humans significantly increased
vasodilation (Bode-Boger et al., 1998)
• Oral L-arginine improves endothelial function in healthy adults older than
70 years (Bode-Boger et al., 2003)
• Oral L-arginine improves endothelial dysfunction in patients with essential
hypertension (Lekakis et al., 2002)
• In cystic fibrosis patients, oral L-arginine was shown to enhance lower
airway NO formation (Grasemann et al., 2005)](https://image.slidesharecdn.com/03a13fa6-b0f1-4f08-b6e0-7071fb43c937-160317233444/85/Agmatine-7-320.jpg)









![In vivo studies in humans
(Bode-Boger et al., 1998)
L-arginine [Plasma arg], mM Peripheral resistance
6 g 0.82 ± 0.059 No effect
30 g 6.22 ± 0.4 Significant decrease](https://image.slidesharecdn.com/03a13fa6-b0f1-4f08-b6e0-7071fb43c937-160317233444/85/Agmatine-17-320.jpg)

![L-arginine triggered [Ca2+
]i release
0 400 800 1200 1600
0.75
0.80
0.85
0.90
0.95
1.00
1.05
0 300 600 900 1200 1500
0.75
0.80
0.85
0.90
0.95
1.00
1.05
Fluor.ratio(340nm/380nm)
5 mM L-arginine
Time, sec Time, sec
2 nM Rauwolscine
L-arg + Rauwolscine](https://image.slidesharecdn.com/03a13fa6-b0f1-4f08-b6e0-7071fb43c937-160317233444/85/Agmatine-20-320.jpg)














![Conclusions
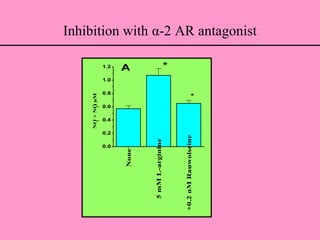
• The NO formation (from both L-arginine and D-arginine) was inhibited by I-
receptor and α-2 AR antagonists, indicating the possible involvement of
receptor(s) in the NO formation.
• The involvement of G-proteins was indicated by the inhibition of NO by
pertussis toxin, a specific inhibitor of Gi/o-proteins.
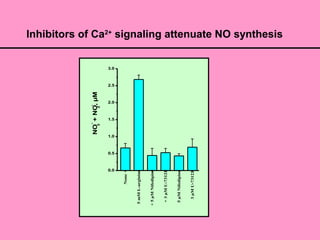
• The addition of L-arginine yielded a transient increase in [Ca2+
]i within 1 min
and Ca2+
dependence of NO formation was demonstrated using inhibitors of
intracellular Ca2+
signaling.
• The agmatine directly activated endothelial NO at low concentrations as
compared to arginine and this could be attenuated by α-2 AR antagonist,
rauwolscine.
• L-arginine-initiated dilation of isolated gracilis vessels is being inhibited by α-
2 AR antagonist, rauwolscine and thus establishing the physiological
relevance of our central hypothesis.](https://image.slidesharecdn.com/03a13fa6-b0f1-4f08-b6e0-7071fb43c937-160317233444/85/Agmatine-37-320.jpg)








































![ARGININE ,HISTIDINE AND BRANCHED CHAIN AMINO ACIDS [Autosaved] HISTIDINE AND ...](https://cdn.slidesharecdn.com/ss_thumbnails/argininehistidineandbranchedchainaminoacidsautosavedhistidineandproline-250321163551-9c372010-thumbnail.jpg?width=640&height=640&fit=bounds)














