Intended Learning outcomes
Bythe end of the session and the associated learning resources,
the student should be able to:
Highlight the typical strategies for viral persistence.
Describe mechanism of persistence in Herpes viruses and HIV
3.
Patterns of viralinfection
Two different approaches for infections
endured by viruses
Acute infection followed by clearance.
Acute infection followed by persistence:
The infection last for long period.
Chronic infection.
Latent infection.
4.
General Principles ofViral persistence
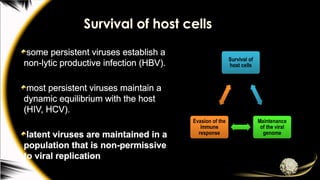
conditions need to be met in order for the virus to avoid clearance;
Survival of
host cells
Maintenance
of the viral
genome
Evasion of the
immune
response
5.
Survival of hostcells
some persistent viruses establish a
non-lytic productive infection (HBV).
most persistent viruses maintain a
dynamic equilibrium with the host
(HIV, HCV).
latent viruses are maintained in a
population that is non-permissive
to viral replication
6.
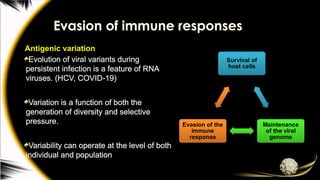
Evasion of immuneresponses
Antigenic variation
Evolution of viral variants during
persistent infection is a feature of RNA
viruses. (HCV, COVID-19)
Variation is a function of both the
generation of diversity and selective
pressure.
Variability can operate at the level of both
individual and population
7.
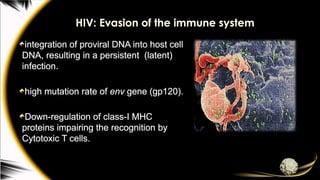
HIV: Evasion ofthe immune system
integration of proviral DNA into host cell
DNA, resulting in a persistent (latent)
infection.
high mutation rate of env gene (gp120).
Down-regulation of class-I MHC
proteins impairing the recognition by
Cytotoxic T cells.
8.
HIV
HIV persists ina number of compartments:
as a dynamic infection of CD4 positive T
cells.
as a low level infection of macrophages.
as partially or fully reverse transcribed,
non-integrated DNA in inactive T cells.
as an integrated but genetically silent pro-
virus in CD4 positive memory T cells
9.
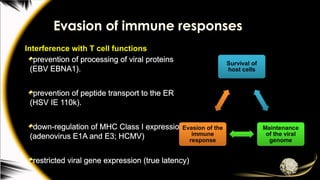
Evasion of immuneresponses
Interference with T cell functions
prevention of processing of viral proteins
(EBV EBNA1).
prevention of peptide transport to the ER
(HSV IE 110k).
down-regulation of MHC Class I expression
(adenovirus E1A and E3; HCMV)
restricted viral gene expression (true latency)
10.
Maintenance of viralgenome
Models:
Alpha-herpesviruses (HSV)
Gamma-herpesviruses (EBV)
11.
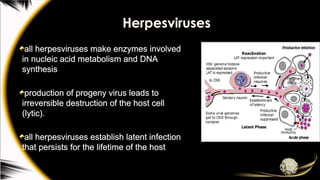
Herpesviruses
all herpesviruses makeenzymes involved
in nucleic acid metabolism and DNA
synthesis
production of progeny virus leads to
irreversible destruction of the host cell
(lytic).
all herpesviruses establish latent infection
that persists for the lifetime of the host
12.
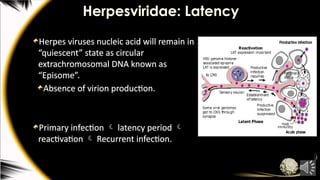
Herpesviridae: Latency
Herpes virusesnucleic acid will remain in
“quiescent” state as circular
extrachromosomal DNA known as
“Episome”.
Absence of virion production.
Primary infection latency period
reactivation Recurrent infection.
13.
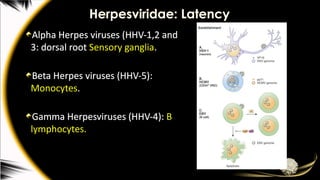
Alpha-herpesviruses
HSV is endemicin all human
populations.
primary infection occurs by intimate
contact with an individual shedding
the virus.
HSV and all other alpha-
herpesviruses establish latent
infection in sensory neurones
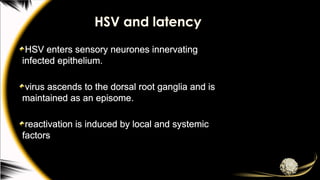
HSV and latency
HSVenters sensory neurones innervating
infected epithelium.
virus ascends to the dorsal root ganglia and is
maintained as an episome.
reactivation is induced by local and systemic
factors
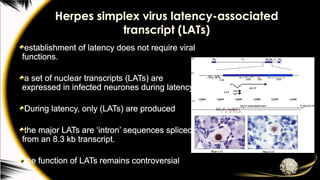
Herpes simplex viruslatency-associated
transcript (LATs)
establishment of latency does not require viral
functions.
a set of nuclear transcripts (LATs) are
expressed in infected neurones during latency.
During latency, only (LATs) are produced
the major LATs are ‘intron’ sequences spliced
from an 8.3 kb transcript.
the function of LATs remains controversial
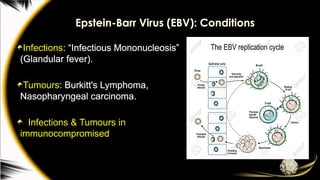
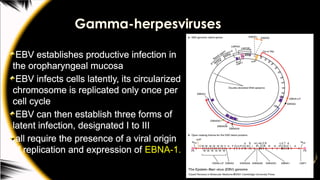
Gamma-herpesviruses
EBV establishes productiveinfection in
the oropharyngeal mucosa
EBV infects cells latently, its circularized
chromosome is replicated only once per
cell cycle
EBV can then establish three forms of
latent infection, designated I to III
all require the presence of a viral origin
of replication and expression of EBNA-1.
22.
OriP
OriP contains twoessential elements
DS: contains 4 low affinity binding
sites for EBNA-1 and is the origin of
replication (kid).
FR: contains 20 copies of a 30 bp
sequence, each containing a high
affinity site for EBNA-1
23.
EBNA-1
is expressed inlatently infected B
lymphocytes that persist for life in healthy
virus carriers and is the only viral protein
EBNA-1 is the only EBV encoded protein
to bind OriP.
the NH2 terminus of EBNA1 has a Gly-
Gly-Ala repeat that inhibits ubiquitinisation.
24.
EBV latency I
Expressionof EBV EBNA-1 (and
LMP-2A) only.
B cells supporting this type of
infection are a major reservoir of
EBV in healthy individuals.
latency I is the pattern found in
EBV positive Burkitt’s lymphoma,
NK-cell lymphoma and gastric
cancer
25.
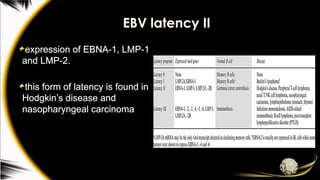
EBV latency II
expressionof EBNA-1, LMP-1
and LMP-2.
this form of latency is found in
Hodgkin’s disease and
nasopharyngeal carcinoma
26.
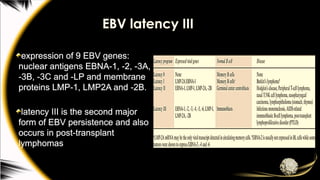
EBV latency III
expressionof 9 EBV genes:
nuclear antigens EBNA-1, -2, -3A,
-3B, -3C and -LP and membrane
proteins LMP-1, LMP2A and -2B.
latency III is the second major
form of EBV persistence and also
occurs in post-transplant
lymphomas
27.
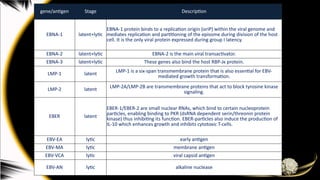
gene/antigen Stage Description
EBNA-1latent+lytic
EBNA-1 protein binds to a replication origin (oriP) within the viral genome and
mediates replication and partitioning of the episome during division of the host
cell. It is the only viral protein expressed during group I latency.
EBNA-2 latent+lytic EBNA-2 is the main viral transactivator.
EBNA-3 latent+lytic These genes also bind the host RBP-Jκ protein.
LMP-1 latent LMP-1 is a six-span transmembrane protein that is also essential for EBV-
mediated growth transformation.
LMP-2 latent LMP-2A/LMP-2B are transmembrane proteins that act to block tyrosine kinase
signaling.
EBER latent
EBER-1/EBER-2 are small nuclear RNAs, which bind to certain nucleoprotein
particles, enabling binding to PKR (dsRNA dependent serin/threonin protein
kinase) thus inhibiting its function. EBER-particles also induce the production of
IL-10 which enhances growth and inhibits cytotoxic T-cells.
EBV-EA lytic early antigen
EBV-MA lytic membrane antigen
EBV-VCA lytic viral capsid antigen
EBV-AN lytic alkaline nuclease
28.
Further Reading
Notes onMedical Virology. 11th Edition. Morag Timbury. Churchill
Livingstone. ISBN 0-443-05845-8.
The Biology of Viruses. Bruce Voyles.Mosby Publishers. ISBN 0-
8016-6391-1. 1993.
https://www.ncbi.nlm.nih.gov/books/NBK8174/
Bashaw JM, Yates JL. Replication from oriP of Epstein-Barr virus
requires exact spacing of two bound dimers of EBNA1 which bend
DNA. J Virol. 2001 Nov;75(22):10603-11. doi:
10.1128/JVI.75.22.10603-10611.2001. PMID: 11602702; PMCID:
PMC114642.
Editor's Notes
#19 Herpes simplex virus types 1 and 2 (HSV1 and HSV2) enter and reactivate from latency in sensory neurons, although the events governing these processes are little understood. During latency, only the latency-associated transcripts (LATs) are produced. However, although the LAT RNAs were described approximately 10 years ago, their function remains ambiguous. Mutations affecting the LATs have minimal effects other than a small reduction in establishment of and reactivation from latency in some cases. Mutations in putative LAT-contained open reading frames (ORFs) have so far shown no effect. The LATs consist of a large species from which smaller (approximately 2 kb), nuclear, nonlinear LATs which are abundant during latency are spliced. Thus, translation of ORFs in these smaller LATs would not usually be expected to be possible, and if expressed at all, their expression might be tightly regulated. Here we show that deregulated expression of the largest HSV1 2-kb LAT-contained ORF in various cells of neuronal and nonneuronal origin greatly enhances virus growth in a manner specific to HSV1-the HSV1 LAT ORF has no effect on the growth of HSV2. Similar results of enhanced growth were found when the HSV1 LAT ORF was constitutively expressed from within the HSV1 genome. The mechanism of LAT ORF action was strongly suggested to be by substituting for deficiencies in immediate-early (IE) gene expression (particularly ICP0), because deregulated LAT ORF expression, as well as enhancing wild-type virus growth, was also found to allow efficient growth of viruses with mutations in ICP0 or VMW65. Such viruses otherwise exhibit considerable growth defects. IE gene expression deficiencies are often the block to productive infection in nonpermissive cells and are also evident during latency. These results, which we show to be protein- rather than RNA-mediated effects, strongly suggest a function of the tightly regulated expression of a LAT ORF-encoded protein in the reactivation from HSV latency.
#23 The Epstein-Barr virus (EBV) encoded nuclear antigen (EBNA) 1 is expressed in latently infected B lymphocytes that persist for life in healthy virus carriers and is the only viral protein regularly detected in all EBV associated malignancies. The Gly-Ala repeat domain of EBNA1 was shown to inhibit in cis the presentation of major histocompatibility complex (MHC) class I restricted cytotoxic T cell epitopes from EBNA4. It appears that the majority of antigens presented via the MHC I pathway are subject to ATP-dependent ubiquitination and degradation by the proteasome















![Epstein barr virus [autosaved]](https://cdn.slidesharecdn.com/ss_thumbnails/epsteinbarrvirusautosaved-211025044258-thumbnail.jpg?width=640&height=640&fit=bounds)







































![ONFH[AVN HIP] -TRIPLE REGIME -A NOVAL SURGICAL CONCEPT .pptx](https://cdn.slidesharecdn.com/ss_thumbnails/onfhavnhip2026koaconcalicutdrgokuldevdrmashraf-260210064517-213ec005-thumbnail.jpg?width=640&height=640&fit=bounds)

![PERI-PROSTHETIC FRACTURE NAIL-PLATE CONSTRUCT [NPC].pptx](https://cdn.slidesharecdn.com/ss_thumbnails/drarunkumardrmohamedashrafperiprostheticfrasturenail-plateconstructnpc-260209164459-7e9d15a1-thumbnail.jpg?width=640&height=640&fit=bounds)




