Downloaded 169 times



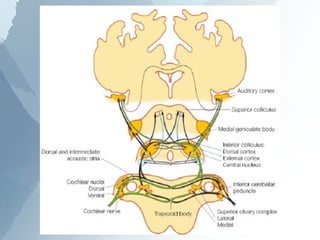


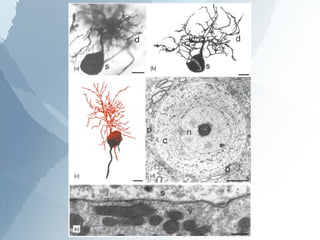


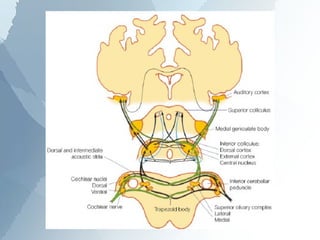




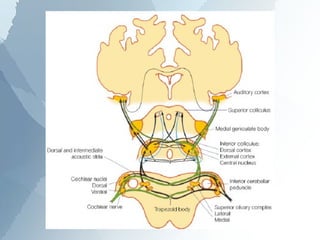

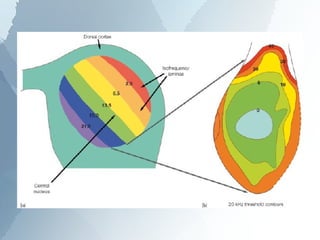



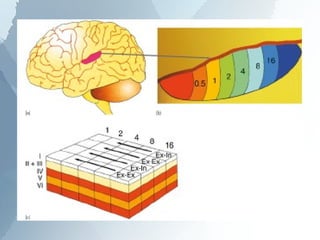

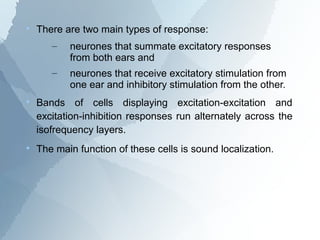










The ascending auditory pathway begins with the cochlea and travels through several regions in the brainstem and midbrain before reaching the auditory cortex. It passes sequentially through the cochlear nuclear complex, superior olivary complex, inferior colliculus, and medial geniculate nucleus. Each region processes auditory information and relays it to the next area, maintaining a tonotopic map of frequencies. The pathway is intricately connected and allows for analysis of binaural cues important for sound localization.



























































![[Neuro] presentation on ear due oct 13 +5](https://cdn.slidesharecdn.com/ss_thumbnails/neuropresentationoneardueoct135-141130235504-conversion-gate02-thumbnail.jpg?width=640&height=640&fit=bounds)













