Molecular control of male fertility for crop hybrid breeding
•Download as PPTX, PDF•
1 like•867 views

Harnessing hybrid vigor or heterosis is a promising approach to tackle the current challenge of sustaining enhanced yield gains of field crops. More than half of the production of major crops such as maize, rice, sorghum, rapeseed, and sunflower comes from hybrid varieties.

Recommended
Recommended
More Related Content
What's hot
What's hot (20)
Similar to Molecular control of male fertility for crop hybrid breeding
Similar to Molecular control of male fertility for crop hybrid breeding (20)
More from Suresh Antre
More from Suresh Antre (14)
Recently uploaded
Recently uploaded (20)
Molecular control of male fertility for crop hybrid breeding
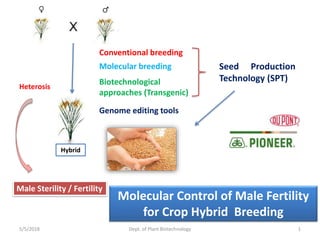
- 1. 5/5/2018 Dept. of Plant Biotechnology 1 Hybrid Heterosis Conventional breeding Molecular breeding Biotechnological approaches (Transgenic) Genome editing tools Seed Production Technology (SPT) Male Sterility / Fertility Molecular Control of Male Fertility for Crop Hybrid Breeding
- 2. 5/5/2018 Dept. of Plant Biotechnology 2 Outline Hybrids cops to increase agricultural yield Types of male sterility Approaches to create male-sterile female lines Genetic resources to exploit heterosis in crop production Mitochondrial genes that control CMS Cytoplasmic–Nuclear interaction in CMS restoration The Seed Production Technology (SPT) platform Case study Summary
- 3. 5/5/2018 Dept. of Plant Biotechnology 3 Charles Darwin Best known for his contributions to the science branching pattern of evolution. Resulted from a process that he called natural selection, in which the struggle for existence has a similar effect to the artificial selection involved in selective breeding. In the 1860s, first described heterosis in maize (Zea mays). Gregor Mendel Genetic basis for inherited traits. Maize became the first commercialized hybrid seed crop. Successively rice, barley, rapeseed etc. Joseph Gottlieb Kölreuter The first documentation of male sterility in 1763. Observed heterosis, that hybrids surpassed their parents.
- 4. 5/5/2018 Dept. of Plant Biotechnology 4 Tremendous contribution to increases agricultural yield. Hybrid plants possess superior traits such as better adaptability, greater uniformity, and improved abiotic and biotic stress tolerance. Wheat (Triticum spp.) 3.5–15% Barley 11% Rice 55% Common bean (Proteus vulgaris) 47% Foxtail millet (Setaria italica) 68% Brassica oilseed crops 200% (Kim and Zhang., 2018)
- 5. 5/5/2018 Dept. of Plant Biotechnology 5 Producing hybrid seeds requires - 1. Cross-pollination of parental lines with genetic diversity. 2. The male-sterile plants must be maintained and propagated for large-scale use as the male-sterile crossing partner. 3. Plant male sterility must be overcome in the F1 hybrid generation to achieve a higher yield for crops whose seeds are harvested.
- 6. 5/5/2018 Dept. of Plant Biotechnology 6 Types of male sterility 1. Genetic Male Sterility (GMS) 1.1 Environmental Sensitive (EGMS) a) Thermo sensitive genetic male sterility (TGMS) b) Photoperiod sensitive genetic male sterility (PGMS) 1.2 Environmental non-sensitive 2. Cytoplasmic Male Sterility (CMS) 3. Cytoplasmic Genetic Male Sterility (CGMS) 4. Transgenic Male Sterility (TMS)
- 7. 5/5/2018 Dept. of Plant Biotechnology 7 Approaches to create male-sterile female lines 1. Chemicals : Auxins, anti-auxins, halogenated aliphatic acids, gibberellins, arsenicals, ethepon applied as foliar sprays prior to flowering have been investigated (McRae, 1985). Drawbacks : Impacts on genotype and environment, as well as the logistical challenges of accurate and timely chemical application. Some chemical applications reduced seed yields in excess of 20% because of damage to the female flower (Newhouse et al., 1996).
- 8. 5/5/2018 Dept. of Plant Biotechnology 8 Approaches to create male-sterile female lines 2. Genetic manipulations : First biotechnology-based approaches to male sterility was proposed by Mariani et al., (1990) and included tapetal-specific expression of a ribonuclease gene, barnase, to cause complete male sterility. This technology is linked with the selectable marker phosphinothricin acetyltransferase (PAT) conferring tolerance to the herbicidal active ingredient glufosinate ammonium. Drawbacks : Timely application of herbicide is essential to remove non transgenic plants, which adds to costs and complexity.
- 9. 5/5/2018 Dept. of Plant Biotechnology 9 Glyphosate-resistant corn was developed via expression of a glyphosate insensitive 5-enolpyruvyl-shikimate 3- phosphate synthase enzyme (CP4-EPSPS). The resulting plant demonstrated sterile tassel following glyphosate application with little to no injury to the rest of the plant. Roundup Hybridization System (RHS) replaces mechanical detasseling with glyphosate spray and greatly simplifies the process of hybrid seed corn production.
- 10. 5/5/2018 Dept. of Plant Biotechnology 10 Genetic Resources to Exploit Heterosis in Crop Production CMS and EGMS lines do not require emasculation and therefore are ideal female lines for hybrid seed production. Three Line Hybrid Seed Production System (Kim and Zhang., 2018)
- 11. 5/5/2018 Dept. of Plant Biotechnology 11 Two-Line Hybrid Seed Production System Nongken 58S (NK58S), O. sativa ssp. Japonica in 1973 completely male sterile when grown under long-day conditions but male fertile when grown under short-day conditions (Shi, M.S., 1985). (Kim and Zhang., 2018)
- 12. 5/5/2018 Dept. of Plant Biotechnology 12 Mitochondrial Genes that Control CMS The mitochondrion is a semiautonomous organelle and power house of cell. The reported CMS DNA sequences encode small transmembrane proteins that are 10–35 kDa in size and cytotoxic. The specific accumulation of cytotoxic CMS proteins within the tapetum, frequently causes altered mitochondrial function such as ATP synthesis and abnormal programmed cell death (PCD) in the sporophytic or gametophytic cells, leading to male sterility. Until now 28 CMS genes have been identified from 13 crop species. e.g. in Rice – orf79, orfH79 etc. (Chen and Liu., 2014)
- 13. 5/5/2018 Dept. of Plant Biotechnology 13( Luo et al., 2013) CMS-related cytoplasmic-nuclear incompatibility is driven by a detrimental interaction between mitochondrial gene and a conserved, essential nuclear gene. Wild Abortive CMS WA 352
- 14. 5/5/2018 Dept. of Plant Biotechnology 14 How CMS genes affect only male development . . . ? It may result from one of three possible types of expression patterns of CMS genes : 1. Male organ (anther)–specific mRNA expression and protein accumulation – NO CMS system 2. Constitutive mRNA expression and protein accumulation throughout plant tissues – orf 129 expression in flowers, roots and leaves. But functions only in male organ (Energy Deficiency). 3. Constitutive mRNA expression but specific or preferential protein accumulation in the male organs (anthers).
- 15. 5/5/2018 Dept. of Plant Biotechnology 15 A model for mitochondrial-nuclear gene interactions in CMS/Rf systems. (Chen and Liu., 2014) CMS restoration by RF proteins can be achieved at the G, R, P, or M level.
- 16. 5/5/2018 Dept. of Plant Biotechnology 16 Proposed a model: “Multi-recombination/protogene formation /functionalization” mechanism . Involves- • Variations in the structure • Sequence • Copy number • Function. Formation and evolution of new CMS genes
- 17. 5/5/2018 Dept. of Plant Biotechnology 17 (Tang et al. 2017) Multi-recombination/protogene formation /functionalization mechanism . Darwin’s Principle of descent with modification Microevolution
- 18. 5/5/2018 18Dept. of Plant Biotechnology Multiple rounds of recombination of the mitochondrial genome during evolution (Kim and Zhang., 2018)
- 19. 5/5/2018 Dept. of Plant Biotechnology 19 Cont. . . PCMS Norin 26 Murai et al., 2016
- 20. 5/5/2018 Dept. of Plant Biotechnology 20 Cytoplasmic–Nuclear Interaction in CMS Restoration Rf genes from the restorer lines are nuclear genes that encode proteins localized to mitochondria that play a retrograde regulatory role by suppressing CMS defects. About half of the identified Rf genes encode pentatricopeptide repeat (PPR)- containing proteins that are localized to mitochondria or plastids. In most cases each Rf functions specifically in a particular type of CMS system, which has a unique phenotype caused by the specific type of mitochondrial DNA and corresponding nuclear gene(s).
- 21. 5/5/2018 Dept. of Plant Biotechnology 21 Until now 14 Rf genes have been isolated from 9 crop plants (Chen and Liu., 2014)
- 22. 5/5/2018 Dept. of Plant Biotechnology 22 Fig - A typical structural arrangement of a PPR protein. (Saha et al., 2007) PPR proteins A typical PPR protein is targeted to mitochondria or chloroplasts, binds one or several organellar transcripts, and influences their expression by altering RNA sequence, turnover, processing, or translation. Pentatricopeptide repeat (PPR) proteins constitute one of the largest protein families in land plants, with more than 400 members in most species. Functional studies on different PPR proteins have revealed their role fertility restoration in CMS plants, embryogenesis, and plant development.
- 23. 5/5/2018 Dept. of Plant Biotechnology 23 Major functions of PPR proteins in plants (Saha et al., 2007)
- 24. 5/5/2018 Dept. of Plant Biotechnology 24 Characterized cytoplasmic male sterility (CMS)/restorer (Rf ) gene systems in major crops (Chen and Liu., 2014)
- 25. 5/5/2018 Dept. of Plant Biotechnology 25 Cont. . . Most CMS genes encodes : • Mitochondrial ETC pathways • Transmembrane proteins (Chen and Liu., 2014)
- 26. 5/5/2018 Dept. of Plant Biotechnology 26 The Seed Production Technology (SPT) Platform The Barnase-Barstar system (Bayer) has been tried in many crops but has encountered several problems. A transgenic construct-driven non- GM (genetically modified) system called ‘seed production technology’ (SPT) was recently developed by DuPont Pioneer. Developed a hybrid seed system that exploits a transgene but generates non- transgenic seeds. The SPT platform consists of a transgenic maintainer line capable of maintaining and propagating non transgenic nuclear male-sterile female lines for hybrid seed production. This novel hybridization system allows the use of nuclear male sterility to produce hybrid seeds in maize, rice, and other cross-pollinating crops.
- 27. 5/5/2018 Dept. of Plant Biotechnology 27 Case Study The maize SPT maintainer line DP-32138-1 is a homozygous recessive male sterile transformed with a SPT construct containing: (i) A complementary wild-type male fertility gene to restore fertility (ii) An a-amylase gene to disrupt pollination (iii) Seed colour marker gene
- 28. 5/5/2018 Dept. of Plant Biotechnology 28 Experimental procedure 1. Maize transformation and molecular analyses of transformants Male-sterile transformation-amenable maize lines, which contain the ms450- 9301 allele were developed by backcrossing with pollen from a transformable maize genotype. Immature embryos that segregated for the ms450-9301 allele were used for transformation. Agrobacterium-mediated transformation 2. Histochemical analysis, pollen in vivo germination and Microscopy 3. Seed sorting and sorter specifications (ScanMasterII 200 high-volume colour sorter) 4. Real-time PCR of male-sterile progeny plants
- 29. 5/5/2018 Dept. of Plant Biotechnology 29 Isolation of the Ms45 male fertility gene Gene Construct used pPG47::Bt1:zm-aa1 linked to DsReds under a constitutive maize ubiquitin (Ubi) Promoter.
- 30. 5/5/2018 Dept. of Plant Biotechnology 30 Fig.- Pollen from transgenic plant at different developmental stages Results : FDA KI Absence of detectable starch in pollen with the presence of pPG47::Bt1:zm-aa1 DsReds
- 31. 5/5/2018 Dept. of Plant Biotechnology 31 Fig.- Pollen kernel characters of transgenic plant Transgenic Non Transgenic
- 32. 5/5/2018 Dept. of Plant Biotechnology 32 Schematic representation of the Seed Production Technology (SPT) process
- 33. 5/5/2018 Dept. of Plant Biotechnology 33 Extremely high efficacy of the SPT genetic process in preventing the transfer of the SPT transgenes to the male-sterile female parents. Large-scale automated seed sorting can be employed to further help ensure the purity of the nontransgenic male-sterile inbred parent seed being increased and subsequently used for commercial scale maize hybrid seed production. The SPT process has the potential to dramatically impact hybrid maize seed production as it eliminates a need for mechanical and/or hand detasseling. Conclusion:
- 34. 5/5/2018 Dept. of Plant Biotechnology 34 Summary CMS proteins may interact with nucleus-encoded mitochondrial factors to induce abnormal programmed cell death in tapeta and microspores, leading to male sterility. CMS/Rf genes may be formed during evolution through gene duplication and multi-recombination. The SPT process has potentially important applications for hybrid seed production in other major crops including rice, sorghum and wheat, which have flowers that are not amenable to manual emasculation. More CMS systems in staple crops need to be studied and applied in agriculture to avoid genetic vulnerability in hybrid crop production that relies on few CMS cytoplasms, such as CMS-WA in rice.
- 35. 5/5/2018 Dept. of Plant Biotechnology 35 https://www.pioneer.com/home/site/about/template.CONTENT/news-media/media- kits/seed-production-technology/guid.B84F55B0-BE0B-6774-F216-69CE365F59A7 For more details on SPT -
- 36. 5/5/2018 Dept. of Plant Biotechnology 36
Editor's Notes
- (a) The three-line system requires a CMS line, containing sterile cytoplasm (S) and a nonfunctional (recessive) restorer (rf ) gene or genes; a maintainer line, containing normal cytoplasm (N) and a nuclear genome identical to that of the CMS line;and a restorer line, with normal (N) or sterile (S) cytoplasm and a functional (dominant) restorer (Rf ) gene or genes. The CMS line is propagated by crossing with the maintainer line; the maintainer and restorer lines can produce seeds by self-pollination. The CMS line is crossed with the restorer line to produce male-fertilehybrids. (b) In the two-line system, an EGMS [photoperiod-sensitive GMS (PGMS), reverse PGMS, or temperature-sensitive GMS (TGMS)] mutant (MT) line is propagated by self-pollination when grown under permissive conditions (PC) (short-day conditions for PGMS, long-day conditions for reverse PGMS, or low-temperature conditions for TGMS). The EGMS line is male sterile under restrictive conditions (RC) (long-day conditions for PGMS, short-day conditions for reverse PGMS, or high-temperature conditions for TGMS) and thus serves as the female parent for crossing with a wild-type (WT) line to produce hybrid seeds.
- (A) Male fertility in EGMS lines containing a mutated nuclear gene (mt) is switched on/off by permissive photoperiod (PGMS) or temperature (TGMS) conditions. EGMS lines are maintained by self-pollination and are used for hybrid seed production through pollination with a restorer line that contains the wild-type (WT) gene.
- Here we report that a new mitochondrial gene, WA352, which originated recently in wild rice, confers CMS-WA because the protein it encodes interacts with the nuclear-encoded mitochondrial protein COX11. In CMS-WA lines, WA352 accumulates preferentially in the anther tapetum, thereby inhibiting COX11 function in peroxide metabolism and triggering premature tapetal programmed cell death and consequent pollen abortion. WA352-induced sterility can be suppressed by two restorer-of-fertility (Rf) genes, suggesting the existence of different mechanisms to counteract deleterious cytoplasmic factors. Thus, CMS-related cytoplasmic-nuclear incompatibility is driven by a detrimental interaction between a newly evolved mitochondrial gene and a conserved, essential nuclear gene. A model for the mechanism of the CMS-WA system. WA352 originated in the mitochondrial genome of wild rice by multiple recombination events. Rf3 and Rf4 suppress WA352 expression at different steps. In CMS-WA plants, the expression of WA352 protein is regulated to accumulate preferentially in the anther tapetum at the MMC stage. WA352 interacts with the nuclear-encoded COX11 (OsCOX11) to inhibit COX11 function in peroxide metabolism, leading to ROS burst and Cyt c release, which cause premature tapetal PCD and consequent sporophytic male sterility.
- The nucleus-encoded mitochondrial-sorting gene (MSG) products—including RF proteins and tissue-specific regulatory factors (TSRFs) and subunits of the mitochondrial electron transfer chain (mtETC) complexes—target to the mitochondria for anterograde regulation. TSRFs may regulate, at the translational or posttranslational level, the male organ (anther)–specific accumulation of CMS proteins for male specificity. The CMS proteins may interact with the mtETC subunits to affect their functionsor redox status or ATP biogenesis, leading to production of retrograde signals (such as reactive oxygen species and cytochrome c release) that trigger aberrant programmed cell death (PCD) in tapeta or microspores. CMS restoration by RF proteins can be achieved at the genomic level (G), mRNA level (R), protein level (P), or metabolic level (M).
- Figure 7 A model of multi-step origination of new CMS genes in plant mitochondrial genomes by a “multi-recombination/protogene formation/functionalization” mechanism. The dynamic, multiple steps of rearrangements among the promoter-active sequence and other mitochondrial genomic source sequences at the specific site (downstream of rpl5 in this case) generated the transcription-competent pre-structures and function-inhibited protogenes, then new functional genes and the consequent new traits and better fitness evolved by acquiring the biological functions via further sequence variations, rearrangements, substoichiometric shifting (SSS), and natural selection.
- Multiple rounds of recombination of the mitochondrial genome during evolution have caused structural mutations in rice. Substoichiometric shifting by changes in gene copy number results in the generation of a functional CMS gene. Diversifying pentatricopeptide repeat (PPR) gene clusters led to functional Rf alleles for CMS restoration. CMS-causing genes identified so far are chimeric mitochondrial genes or genes of unknown origin. Nuclear Rf genes reverse CMS gene function at transcriptional (Tc) or protein (P) level, while the rf17 recessive allele is retrogradely (R) upregulated by CMS gene. Unlike most CMS lines, wheat CMS lines such as CMS-Norin 25 were created by the introduction of cytoplasm from a related wild grass, Aegilops, and show a photoperiod-sensitive phenotype (PCMS) caused by unprocessed mitochondrial orf25, which can be restored by Rfd1.
- Fig. 1. (A) A typical structural arrangement of a pentatricopeptide repeat protein.).
- Figure 1 A schematic representation of a system for propagation of recessive genetic male-sterile (ms) plants. Ms, male fertility gene; P, pollination disruption gene; S, seed screenable marker gene; SPT, Seed Production Technology