
Recommended
More Related Content
What's hot
What's hot (17)
Similar to 6 - Транскрипция
Similar to 6 - Транскрипция (15)
More from tophisopam
More from tophisopam (6)
6 - Транскрипция
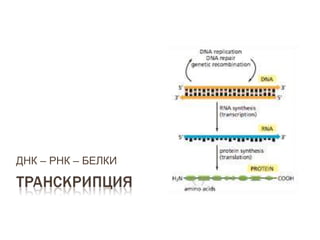
- 1. ДНК – РНК – БЕЛКИ
- 2. Отличия РНК от ДНК • Рибоза вместо 2’-O- дезоксирибозы • Урацил вместо тимина
- 5. WEB-LOGO TATGTT CATGAT TATGCT TACTGT TGTCAT TTAACT TAGACT TAGGTT TATGAT TATAAT TATAAT TATTAT • Высота стэка букв отражает консервативность • Высота индивидуальных буквы отражает частоту встречаемости нуклеотидов (аминокислот)
- 6. Расположение промоторов в ДНК (аденовирус) Важное отличие РНК-полимераз от ДНК-полимераз: им не нужна затравка (праймер)
- 7. Роль σ-субъединицы -35 -10 E.coli 70 TTGACA TATAAT E.coli 32 TCTC-CCCTTGAA CCCCAT-TA E.coli 54 (-24)CTGG-A (-12)TTGCA B. sub A TTGACA TATAAT B. sub AGGTTTAA GGGTAT B. sub D CTAAA CCGATAT B. sub E ATATT ATACA B. sub K AC CATA---T B. sub H CAGGA GAATT—T SPO1gp28 AGGAGA TTT-TTT T4gp55 - TATAAATA
- 9. Уровни регуляции экспрессии генов • На уровне транскрипции • На уровне стабильности мРНК • На уровне трансляции • На уровне стабильности белка
- 10. Анти-σ факторы • Связывают σ-фактор, препятствуя инициации транскрипции • Обычно регулируют специфические σ-факторы, необходимые для споруляции и т.д. • Некоторые бактериофаги кодируют свои анти-σ- факторы, чтобы заблокировать синтез клеточных мРНК
- 11. Транскрипция генов бактериофага SPO1 • Транскрипция ранних мРНК, среди которых есть мРНК σ28, осуществляется клеточным σ-фактором. • σ28 запускает транскрипцию средних генов, в том числе и σ34. • σ34 отвечает за транскрипцию поздних мРНК.
- 12. Элонгация транскрипции •Белок NusA вызывает паузы при элонгации на определенных последовательностях •Белок NusG, наоборот, препятствует паузам •GreA и GreB узнают «арестованные» комплексы, в которых РНК-полимераза «съехала» назад и содержит в активном центре не 3’-конец РНК, а дуплекс РНК-ДНК •Mfd узнает комплексы, в которых полимераза наткнулась на непреодолимое препятствие, например, тиминовый димер.
- 14. • Скорость транскрипции 20-50 нуклеотидов в секунду при 37оС
- 15. Транскрипция и трансляция •У прокариот трансляция начинается еще до того, как синтезируется полноразмерная мРНК. •У эукариот процессы транскрипции и трансляции разделены в пространстве и во времени.
- 16. Регуляция лактозного оперона •Лактоза – менее питательный субстрат для бактерий, чем глюкоза •Белок LacI в отсутствие лактозы связывается с промоторной областью оперона и блокирует транскрипцию. •Когда лактоза есть, LacI связывает ее, и не связывает ДНК. •В отсутствие глюкозы в клетке накапливается cAMP, связывая который активируется CAP (catabolism activation protein). •Активированный САР связывается с с лактозным опероном, и включает его транскрипцию.
- 17. Искусственные системы на основе lac- оперона • Можно заставлять клетку (бактериальную) включать или выключать синтез мРНК с введенного в клетку гена • Для активации можно использовать негидролизуемый аналог лактозы: IPTG
- 18. Оперон • Функциональная единица транскрипции • В одной мРНК закодированы несколько белков, участвующих в одном и том же процессе • У эукариот (за редчайшим исключением!) в одной мРНК всегда закодирован один единственный белок
- 19. Терминация транскрипции (ρ-зависимая) Фактор ρ (ро) – РНК-зависимая АТФаза (хеликаза) Садится на мРНК и «ползет» в направлении 5’->3’ и, когда «догоняет» РНК-полимеразу, терминирует транскрипцию
- 20. Терминация транскрипции (ρ-независимая) В других случаях терминация транскрипции происходит вследствие сигнала в самой мРНК: стабильная шпилька, за которой следует U- богатый участок
- 21. Рибосома и РНК-полимераза. Кто-кого? I При недостатке в клетке UTP рибосома ползет быстрее, но когда UTP много – полимераза успевает синтезировать терминаторный сигнал. Регуляция синтеза мРНК, кодирующей белок, необходимый для синтеза UTP. Соответственно, когда UTP в клетке достаточно - этот белок не нужен.
- 22. Альтернативные структуры в РНК
- 23. Рибосома и РНК-полимераза. Кто-кого? II Когда в клетке мало триптофана, рибосома ползет медленно и образуется более стабильная структура мРНК, предотвращающая терминацию. Регуляция белка, необходимого для синтеза триптофана. Когда триптофана в клетке много - этот белок не нужен.
- 24. Регуляция trp-оперона у Bacillus subtilis
- 26. «Незаряженная тРНК» как сигнал для транскрипции
- 27. Когда вся тРНК проаминоацилирована - транскрипция не идет
- 28. РНК-полимеразы эукариот Полимераза Функция I 18S, 4.5S, 28S rRNA II mRNA, snoRNA, snRNA III 5S rRNA, tRNA, U6 sRNA, vRNA
- 31. Отличия от прокариот • Больше ДНК • Хроматин • Ядро • По умолчанию у прокариот гены включены, у эукариот - выключены Как следствие, больше возможностей для регуляции
- 32. Еще сложнее! • Альтернативные факторы транскрипции • Альтернативные промоторы • Альтернативные терминаторы транскрипции • Альтернативный сплайсинг
- 33. И еще сложнее! • Энхансеры могут регулировать транскрипцию на огромном расстоянии • Иногда находятся вообще на другой хромосоме
- 35. • α-аманитин • Ингибирует РНК- полимеразы II и III • 5-6 мг летальны для человека • Смерть наступает через несколько дней, и это отражает время жизни мРНК у человека