2
Receptors
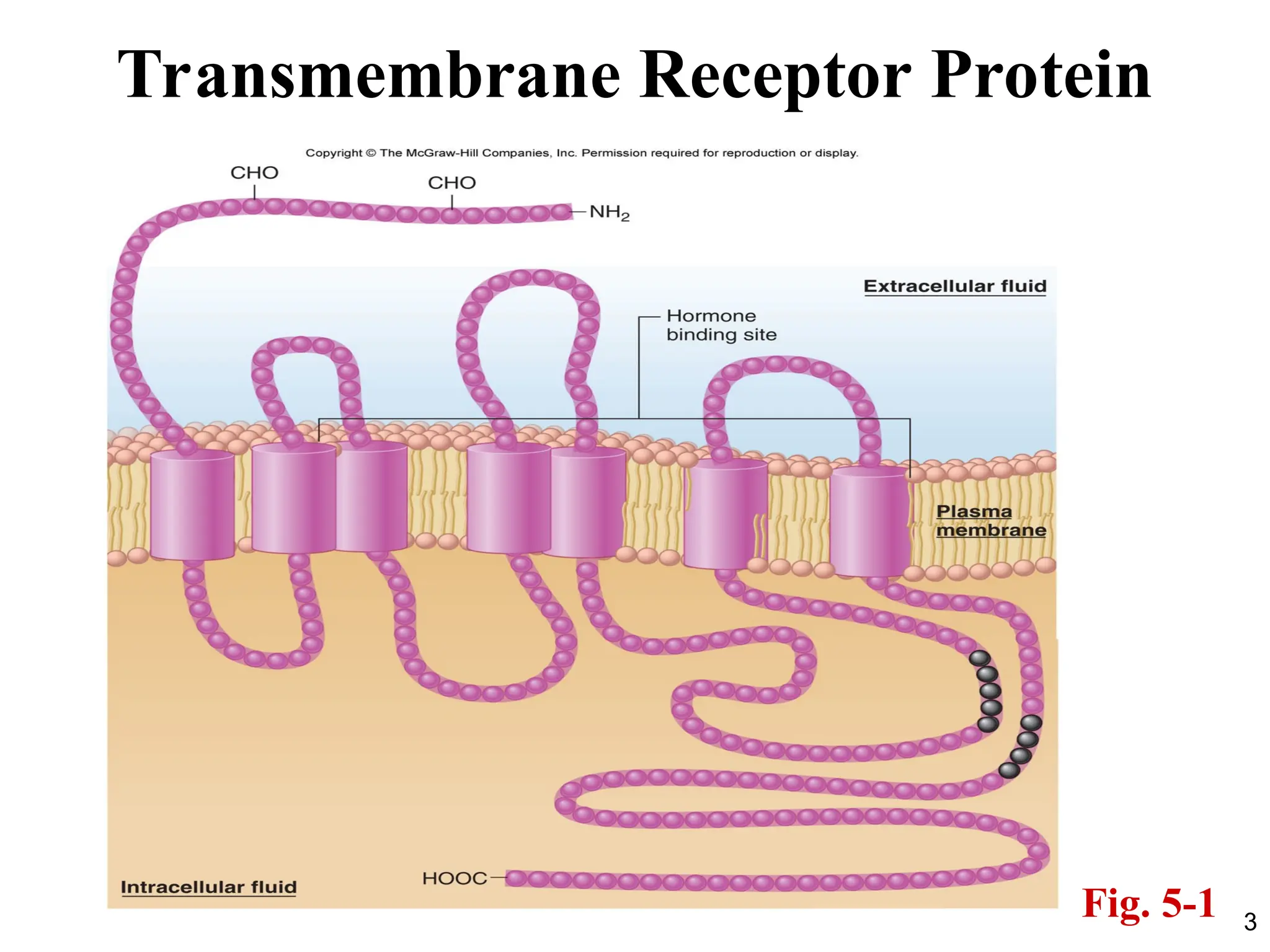
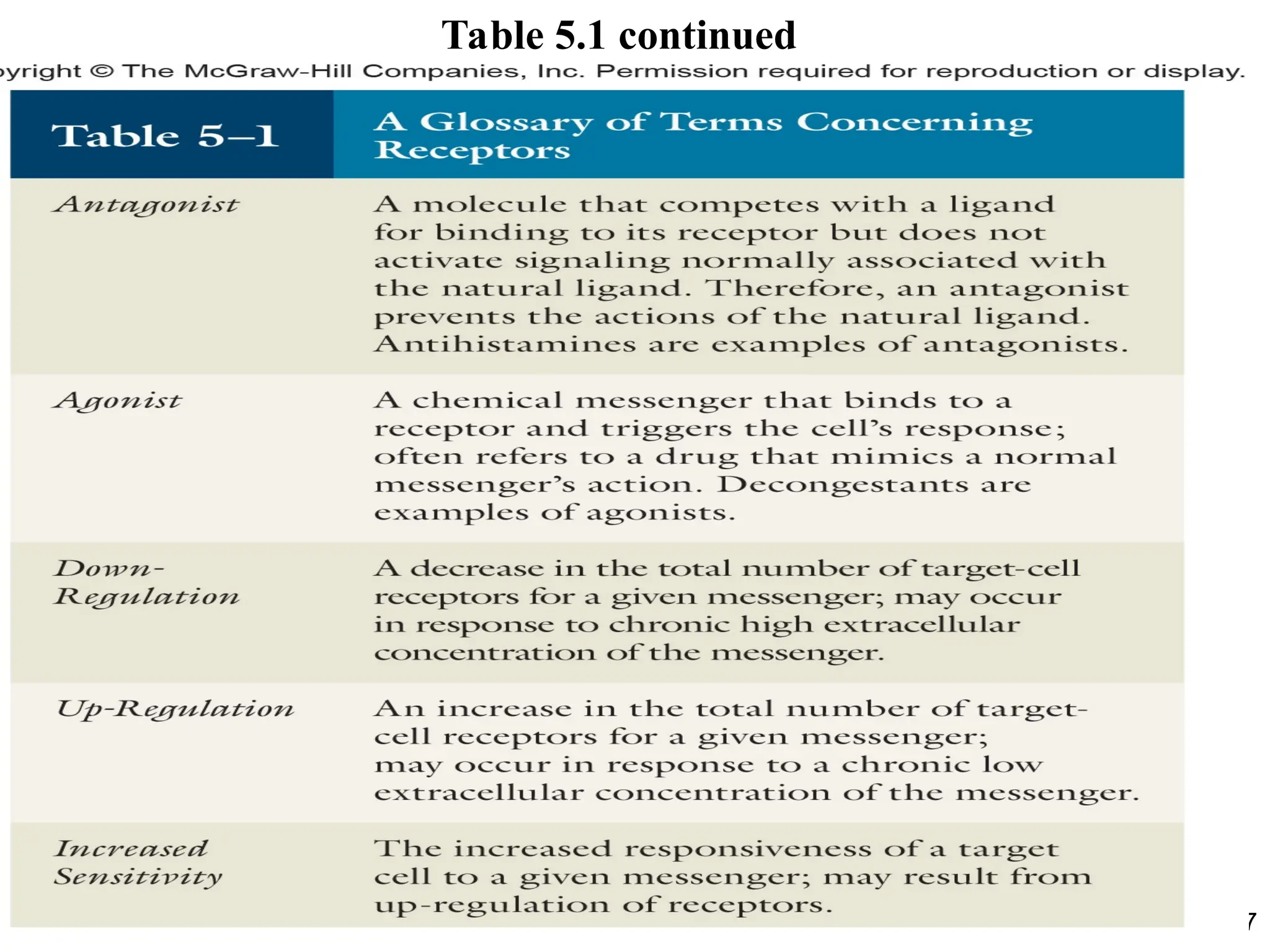
• Chemical messengersbind to proteins called
receptors.
• Most chemical messengers are water-soluble
and bind to receptors located at the plasma
membrane.
• Some messengers like steroids are lipid-
soluble and bind to an intracellular receptor.
4
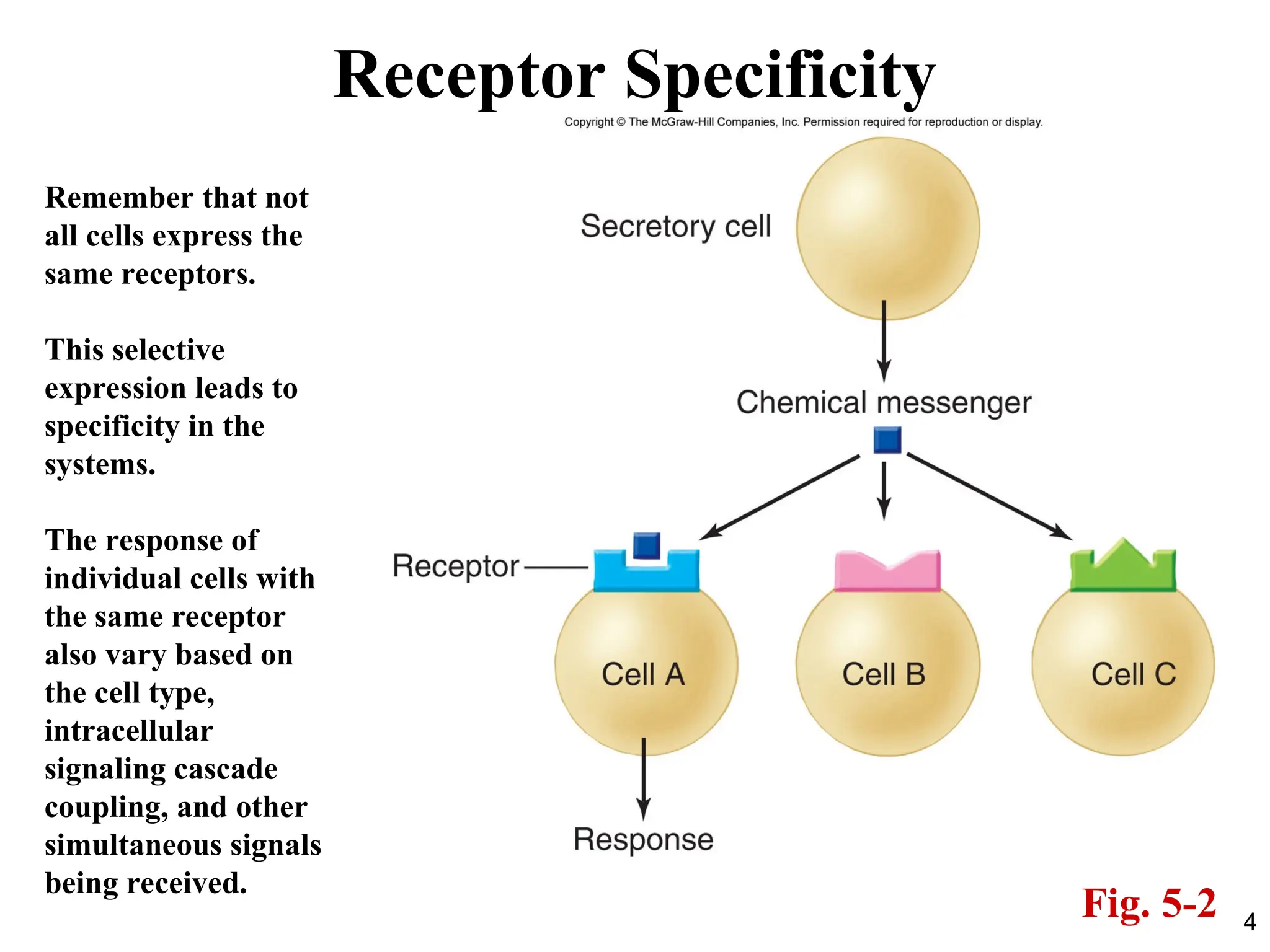
Receptor Specificity
Fig. 5-2
Rememberthat not
all cells express the
same receptors.
This selective
expression leads to
specificity in the
systems.
The response of
individual cells with
the same receptor
also vary based on
the cell type,
intracellular
signaling cascade
coupling, and other
simultaneous signals
being received.
8
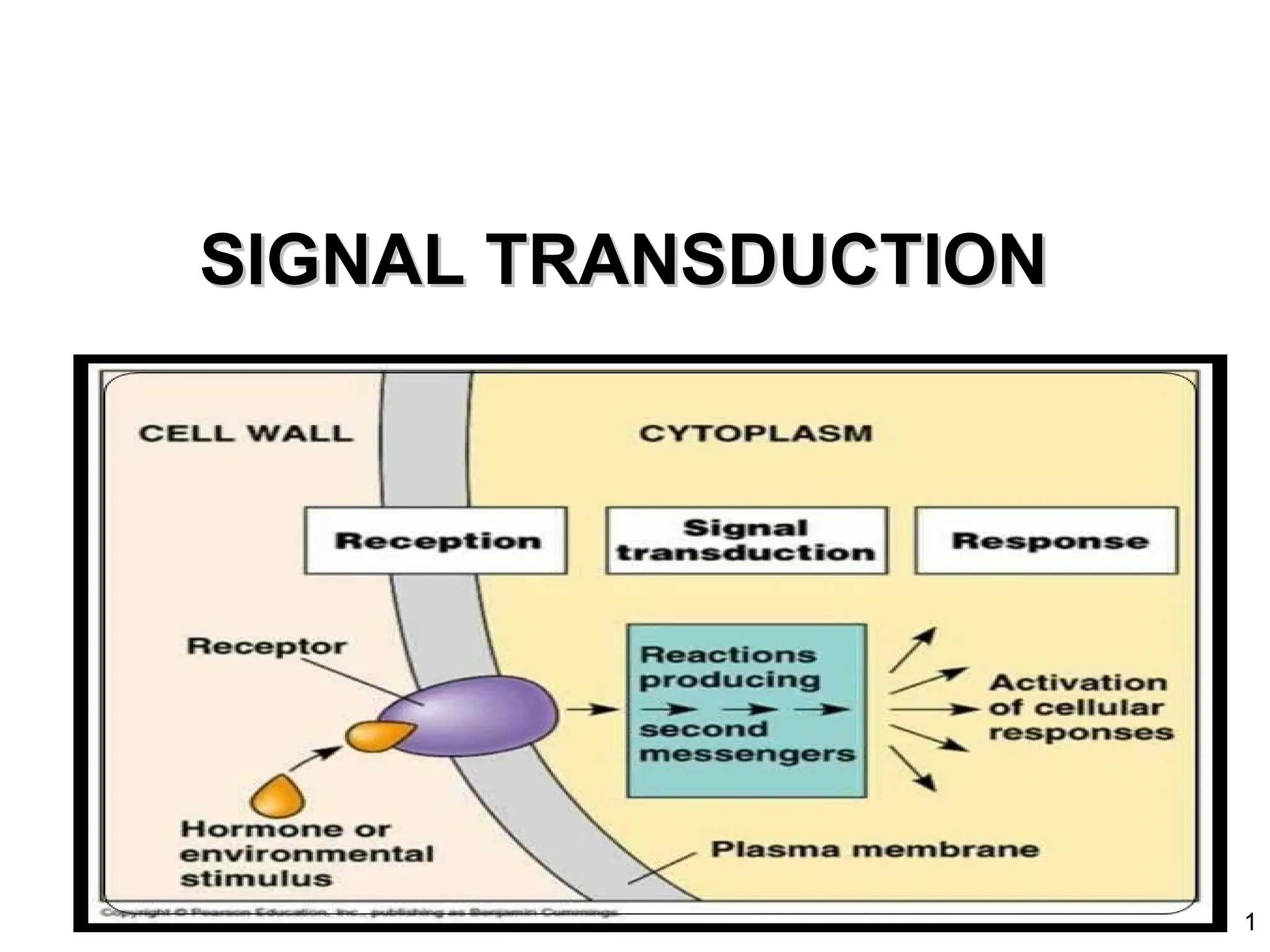
Signal Transduction Pathways
•Asignal transduction pathway is a series of steps by which a
signal on a cell’s surface is converted into a specific cellular
response
10
Pathways Initiated byLipid-Soluble Messengers
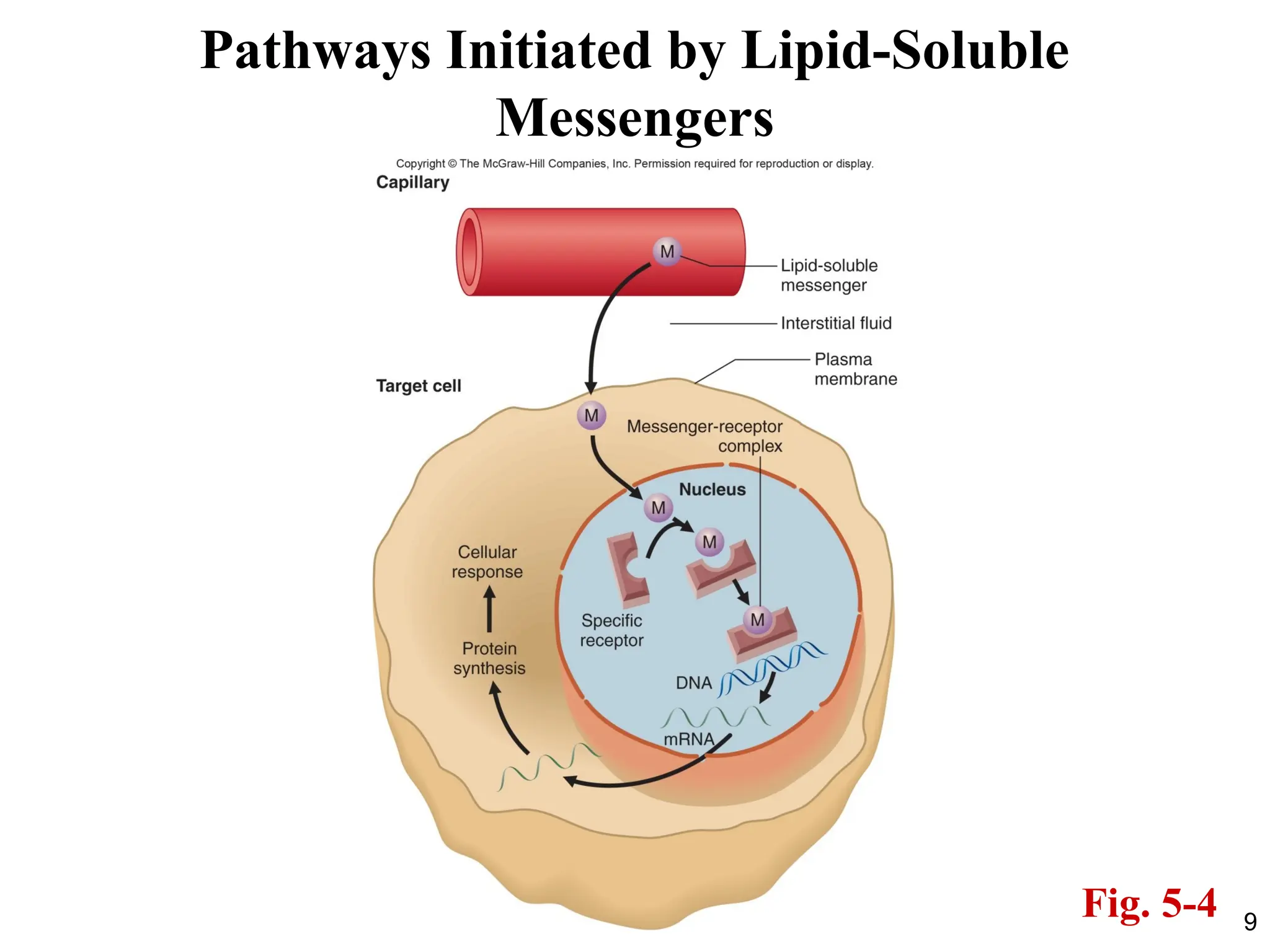
• Critical Points to remember:
1. Lipid messengers can diffuse through the plasma
membrane.
2. They have intracellular receptors.
3. The receptors bind directly to recognized
sequences in the DNA and alter gene
transcription.
4. This is a slower response compared to membrane
receptors, but it is a sustained response.
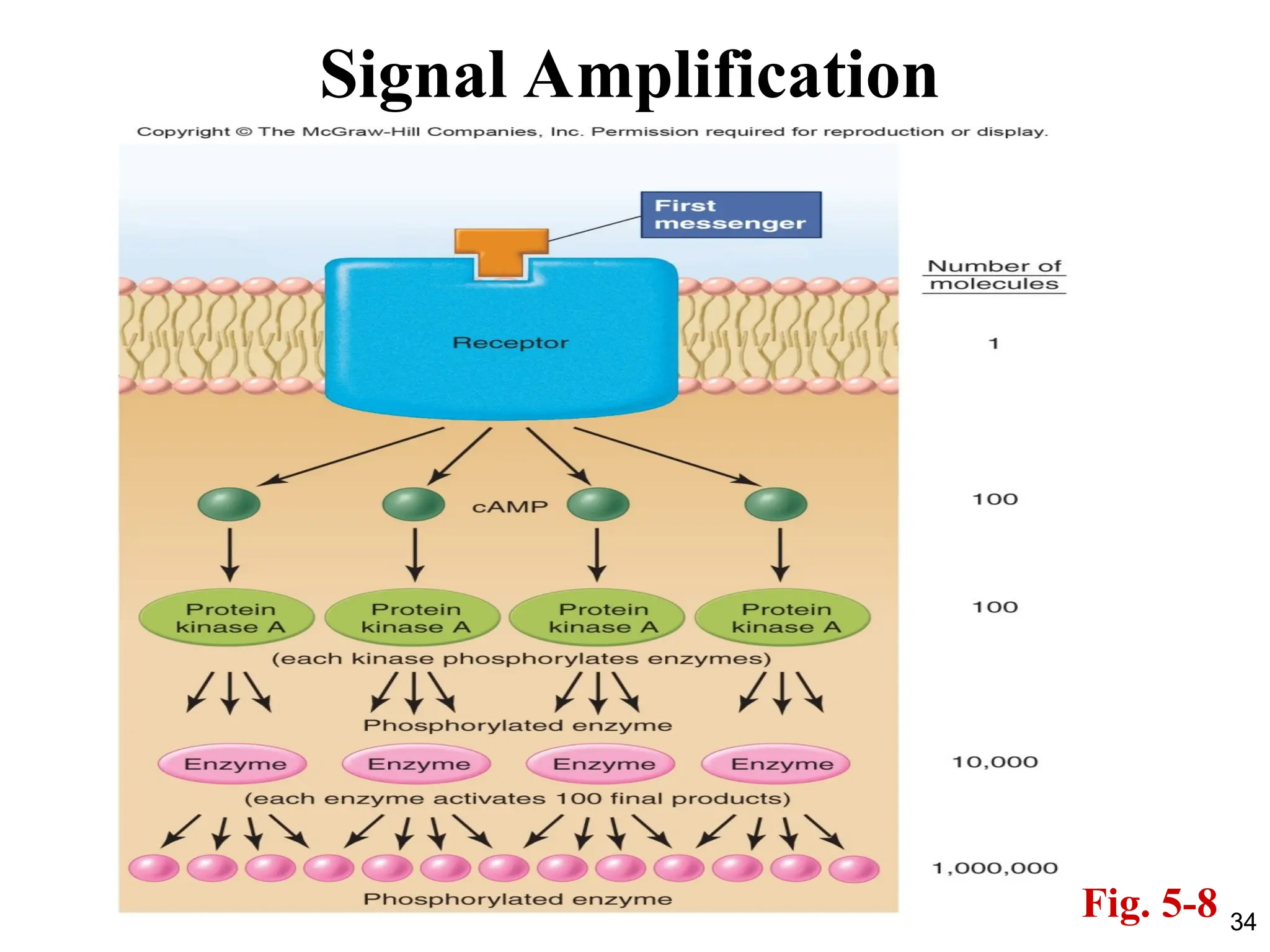
SECONDARY MESSENGERS
• Secondmessengers are molecules that relay
signals from receptors on the cell surface to
target molecules inside the cell.
• They greatly amplify the strength of the signal,
cause some kind of change in the activity of
the cell
• They are a component of cell signaling
pathways
12
14
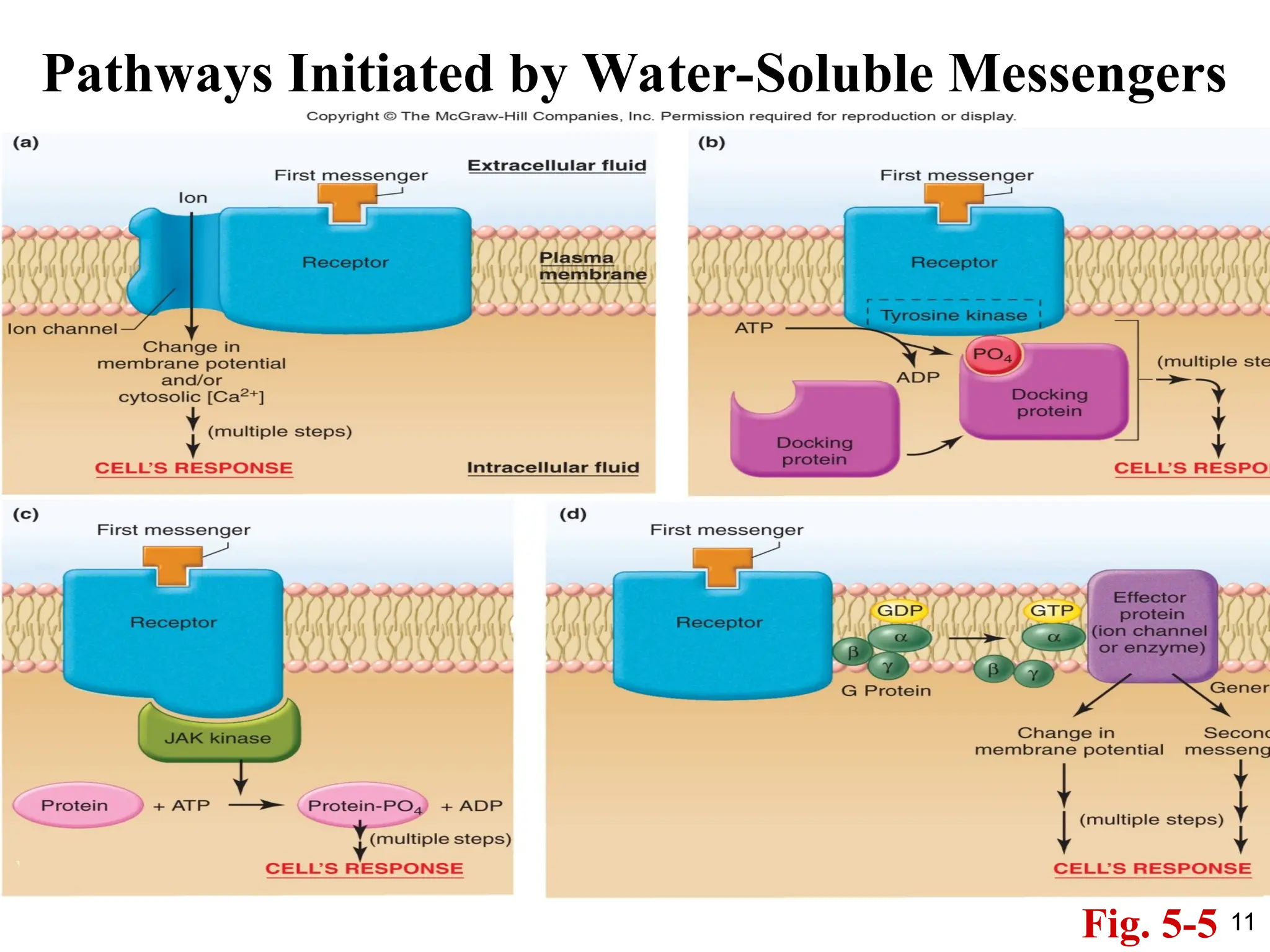
Pathways Initiated byMembrane-Bound Receptors
1. This is a broad range of receptors: ion channels,
G-protein coupled receptors, receptors with
intrinsic kinase activity, etc.
2. These receptors activate intracellular signaling
cascades that affect cell function.
3. These receptors can activate downstream
mediators which affect DNA transcription but
also have many other effects in the cell.
4. This is a faster response compared to
lipid/steroid receptors, but it is a less sustained
response.
16
Receptors that Functionas Enzymes
• Some receptors ( like the insulin receptor) have intrinsic
enzyme activity.
• Most of these receptors that possess intrinsic enzyme activity
are all protein kinases that specifically phosphorylate the
amino acid tyrosine (receptor tyrosine kinases).
• The typical sequence of events for receptors with intrinsic
tyrosine kinase activity is:
1. The binding of a specific messenger to the receptor changes
the conformation of the receptor so that its enzymatic portion,
located on the cytoplasmic side of the plasma membrane, is
activated.
17.
17
2. This resultsin autophosphorylation of the receptor (the
receptor phosphorylates its own tyrosine groups).
3. The newly created phosphotyrosines on the cytoplasmic
portion of the receptor then serve as docking sites for
cytoplasmic proteins.
4. The bound docking proteins then bind and activate other
proteins, which in turn activate one or more signaling
pathways within the cell.
• The common denominator of these pathways is that they all
involve activation of cytoplasmic proteins by phosphorylation.
Receptors that Function as Enzymes
18.
18
Signaling
• The numberof kinases that mediate these
phosphorylations can be very large, and their
names constitute a veritable alphabet soup—
RAF, MEK, MAPKK, and many others.
• Most of the receptors with intrinsic tyrosine
kinase activity bind ligands that typically
influence cell proliferation and differentiation,
and are often called growth factors.
19.
19
cGMP
• The onemajor exception to the generalization is Guanylyl
Cyclase.
• Guanylyl cyclase is a receptor that acts to catalyse the
formation, in the cytoplasm, of a molecule known as
cyclic GMP (cGMP).
• In turn, cGMP functions as a second messenger to
activate a protein kinase called cGMP-dependent
protein kinase.
• This kinase phosphorylates specific proteins that then
mediate the cell’s response to the original messenger.
20.
20
cGMP cont’d
• Receptorsthat function both as ligand-binding molecules
and as guanylyl cyclases are present in high amounts in
the retina of the vertebrate eye, where they are important
for processing visual inputs.
• This signal transduction pathway is used by only a small
number of messengers and should not be confused with
the much more prevalent cAMP system.
• Also, in certain cells, guanylyl cyclase enzymes are
present in the cytoplasm. In these cases, a first messenger
—nitric oxide—diffuses into the cell and combines with
the guanylyl cyclase there to trigger the formation of
cGMP.
22
Receptors that Interactwith Cytoplasmic Kinases
• There are several families of cytoplasmic protein kinases:
src, JAKs, etc.
• These receptors do not have intrinsic kinase activity, but
must use a cytoplasmic kinase.
• The binding of a ligand to the receptor causes a
conformational change in the receptor that leads to
activation of the JAK kinase.
• Janus kinases (JAK) are a commonly used cytoplasmic
kinase.
The Janus kinases are a family of 4 kinases that are all tyrosine
kinases. They are differentially expressed among the tissues in
the body.
23.
23
JAK Kinases
• Differentreceptors associate with different members of the JAK
kinase family, and the different JAK kinases phosphorylate
different target proteins, many of which act as transcription
factors.
• JAK’s traditional targets are the Signal Transducers of Activated
transcription (STATs). However, they have also been shown to
interact with other proteins.
• The result of these pathways is the synthesis of new proteins,
which mediate the cell’s response to the first messenger.
• Signaling by cytokines—proteins secreted by cells of the immune
system that play a critical role in immune defenses—occurs
primarily via receptors linked to JAK kinases.
25
G Protein-Coupled Receptors
•Bound to the inactive receptor is a protein complex
located on the cytosolic surface of the plasma membrane
and belonging to the family of heterotrimeric (containing
three different subunits) proteins known as G proteins.
• All G proteins contain three subunits, called the alpha,
beta and gamma subunits. The alpha subunit can bind
GDP and GTP. The beta and gamma subunits help anchor
the alpha subunit in the membrane.
• The binding of a ligand to the receptor changes the
conformation of the receptor.
26.
26
G Protein-Coupled Receptors
•This activated receptor increases the affinity of the alpha
subunit of the G protein for GTP.
• When bound to GTP, the alpha subunit dissociates from
the beta and gamma subunits.
• This dissociation allows the activated alpha subunit to
link up with still another plasma membrane protein, either
an ion channel or an enzyme.
• These ion channels and enzymes are termed plasma
membrane effector proteins because they mediate the next
steps in the sequence of events leading to the cell’s
response.
28
G Protein-Coupled Receptors
•There are several subfamilies of plasma membrane G
proteins, each with multiple distinct members, and a
single receptor may be associated with more than one
type of G protein.
Examples: Gs, Gi, Gq, Gα
• Moreover, some G proteins may couple to more than one
type of plasma membrane effector protein.
• G-protein coupled receptors are the most numerous type
of receptor family and have a large variety of signaling
pathways associated with them.
29.
29
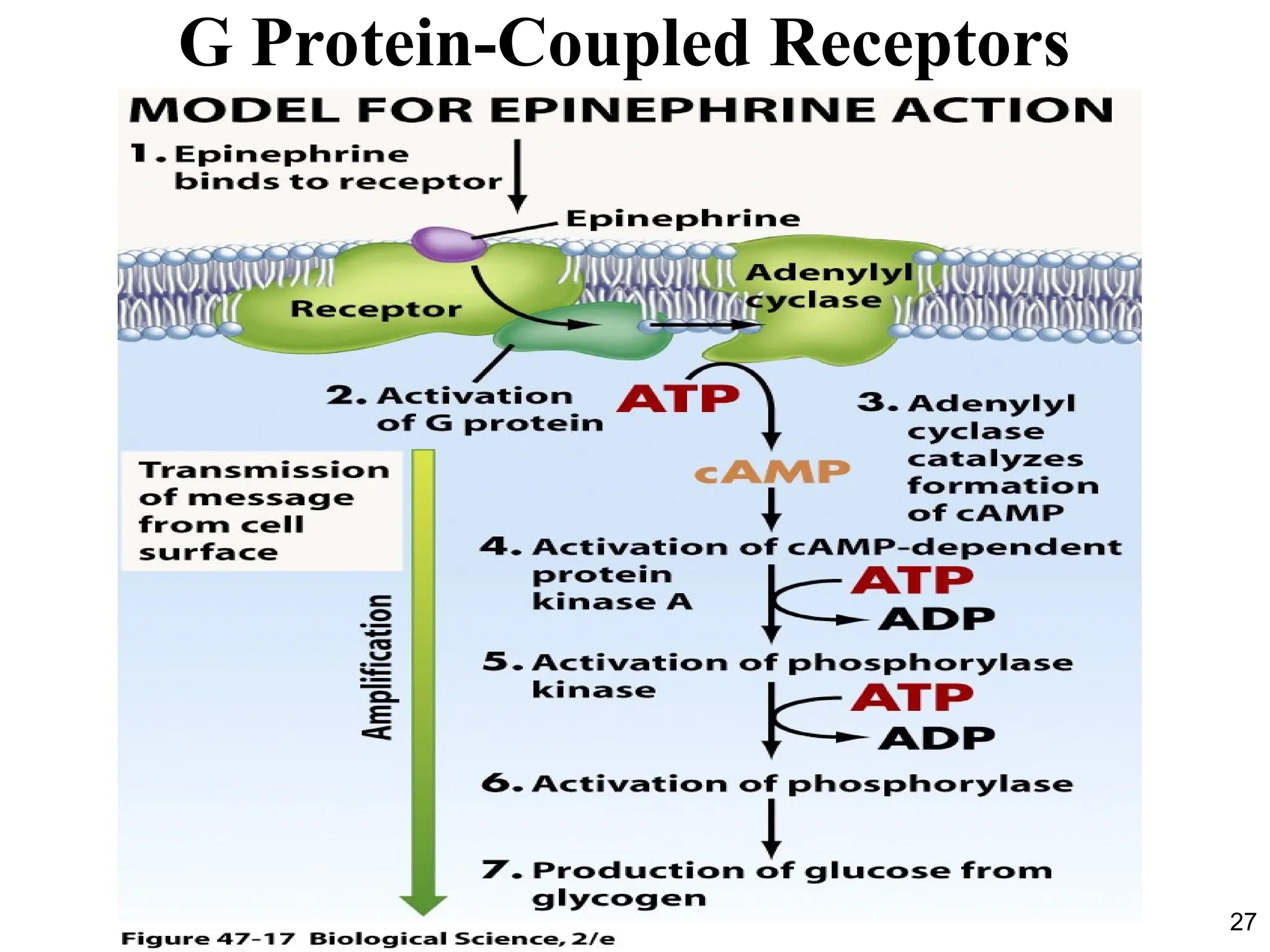
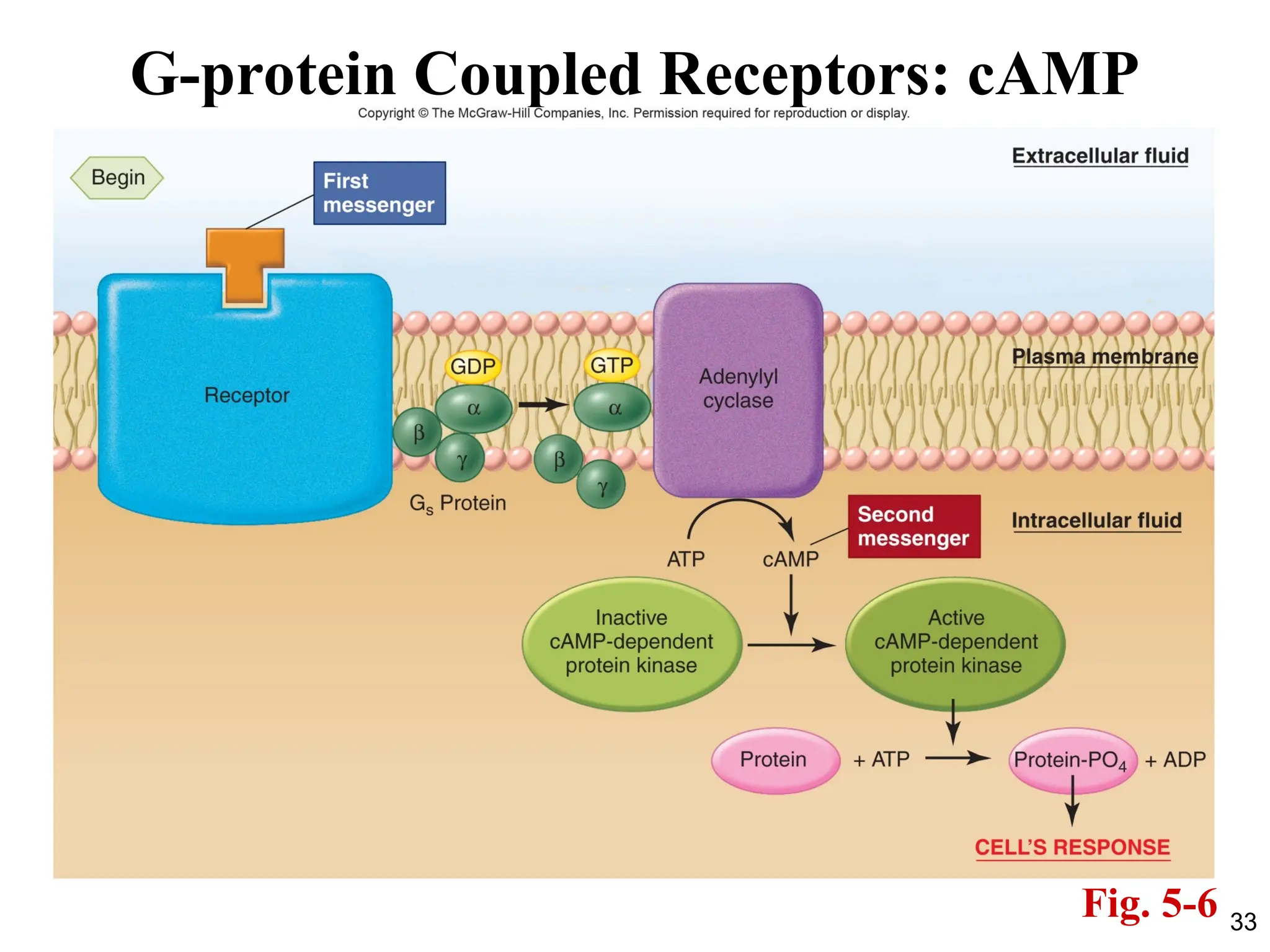
Adenylyl Cyclase andCyclic AMP
• Activation of the receptor by the binding of ligand (for
example, the hormone epinephrine) allows the receptor to
activate its associated G protein (Gs ; “stimulatory”).
• This causes Gs to activate its effector protein, the membrane
enzyme called adenylyl cyclase (also known as adenylate
cyclase).
• The activated adenylyl cyclase catalyzes the conversion of
cytosolic ATP molecules to cyclic 3´,5´-adenosine
monophosphate, or cyclic AMP (cAMP).
• Cyclic AMP then acts as a second messenger.
30.
30
Adenylyl Cyclase andCyclic AMP
• The action of cAMP eventually terminates when it is
broken down to noncyclic AMP, a reaction catalyzed by
the enzyme cAMP phosphodiesterase.
• Thus, the cellular concentration of cAMP can be changed
either by altering the rate of its messenger-mediated
generation or the rate of its phosphodiesterase-mediated
breakdown.
• Caffeine and theophylline, the active ingredients of coffee
and tea, are widely consumed stimulants that work partly
by inhibiting phosphodiesterase activity, which results in
prolonging the actions of cAMP within a cell.
31.
31
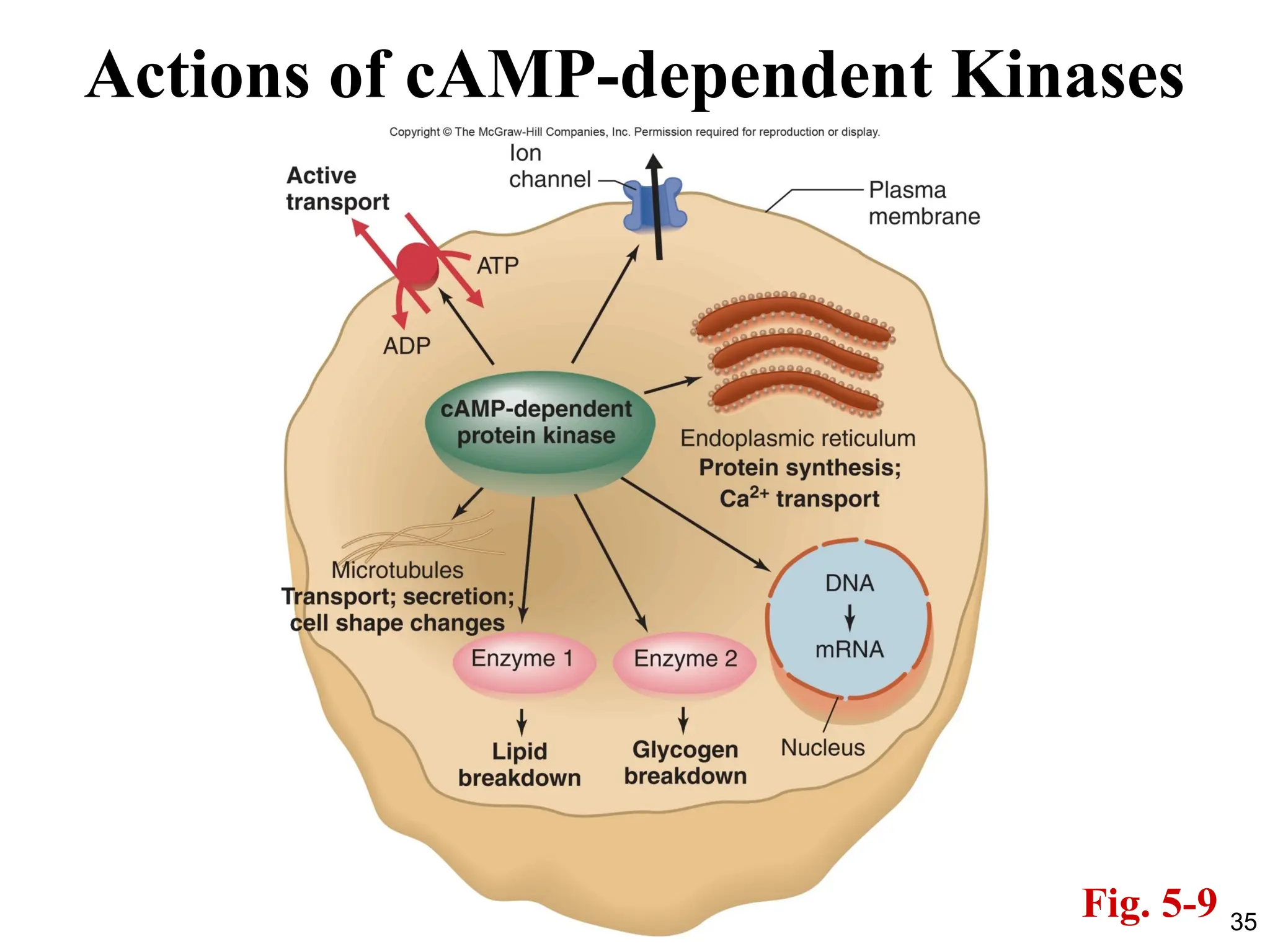
Adenylyl Cyclase andCyclic AMP
• Inside the cell, cAMP binds to and activates an enzyme
known as cAMP-dependent protein kinase (PKA).
PKA then phosphorylates downstream targets.
• Examples: epinephrine acts via the cAMP pathway on fat
cells to stimulate the breakdown of triglyceride, a process
that is mediated by one particular phosphorylated
enzyme. In the liver, epinephrine acts via cAMP to
stimulate both glycogenolysis and gluconeogenesis,
processes that are mediated by phosphorylated enzymes
that differ from those in fat cells.
32.
32
Gi Proteins
• Notall G proteins stimulate cAMP; some inhibit adenylyl
cyclase.
• This inhibition results in less, rather than more, generation of
cAMP.
• This occurs because these receptors are associated with a
different G protein known as Gi (“inhibitory’’).
• Activation of Gi causes the inhibition of adenylyl cyclase. The
result is to decrease the concentration of cAMP in the cell and
thereby the phosphorylation of key proteins inside the cell.
38
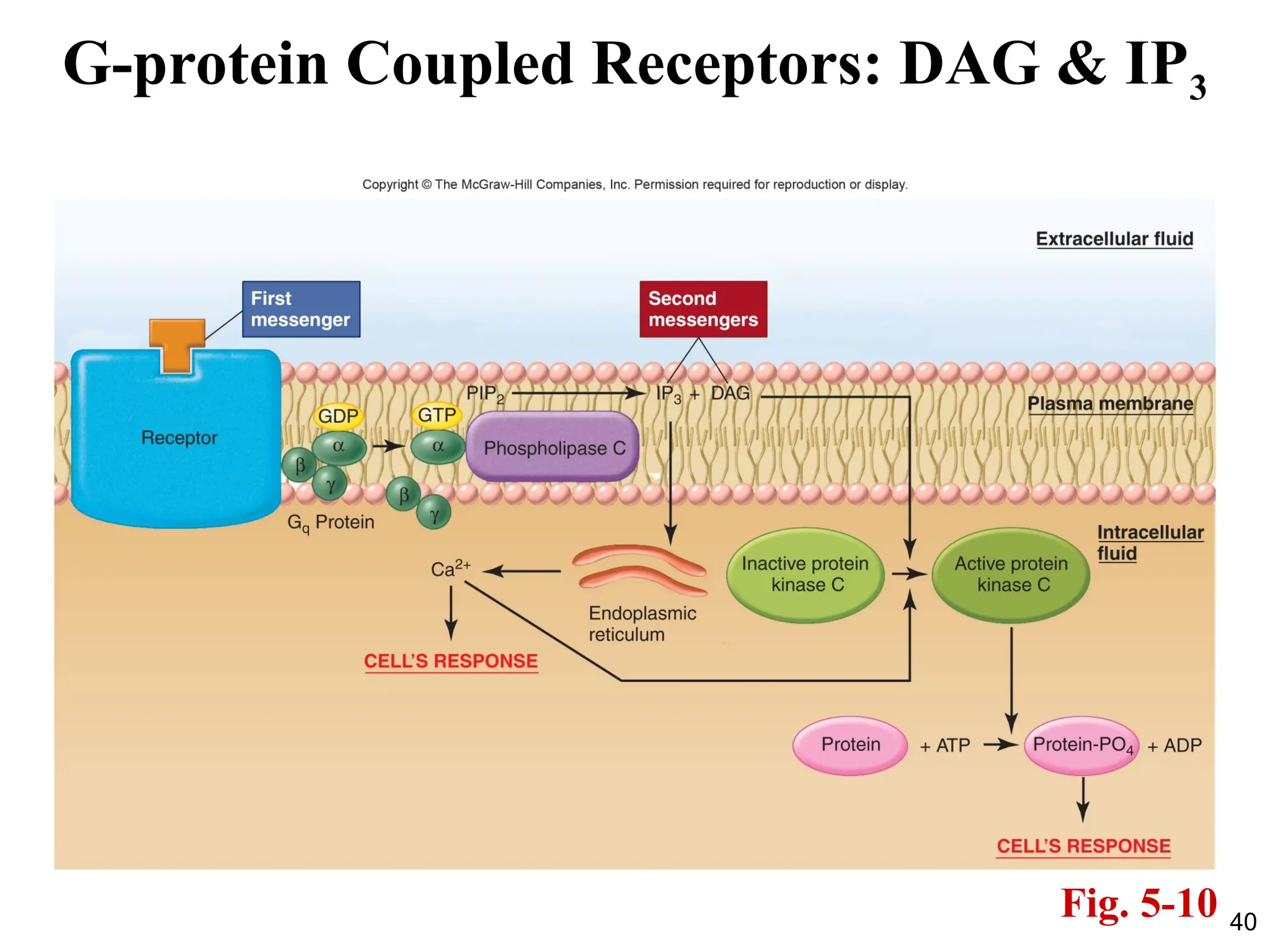
PLC, DAG, IP3
•Both DAG and IP3 then function as second messengers.
• DAG activates a class of protein kinases known
collectively as protein kinase C (PKC), which then
phosphorylate a large number of other proteins, leading
to the cell’s response.
• There are currently 13 known isoforms of PKC which
contribute to the large variety of cellular responses
observed.
39.
39
IP3
• IP3 bindsto receptors located on the endoplasmic
reticulum.
• These receptors are ligand-gated Ca2+
channels which when
bound to IP3 open and result in increased cytosolic Ca2+
concentration.
• This increased Ca2+
concentration then continues the
sequence of events leading to the cell’s response.
• One of the actions of Ca2+
is to help activate some forms of
protein kinase C.
41
Ca2+
as a SecondMessenger
• Ca2+
functions as a second messenger in many pathways.
• Ca2+
can be either increased or decreased cytosolically to elicit a
cellular response (change membrane potential). Ca2+
also has
direct actions on other signaling proteins.
• By means of active-transport systems in the plasma membrane
and cell organelles, Ca2+
is maintained at an extremely low
concentration in the cytosol.
• Consequently, there is always a large electrochemical gradient
favoring diffusion of Ca2+
into the cytosol via Ca2+
channels found
in both the plasma membrane and the endoplasmic reticulum.
42.
42
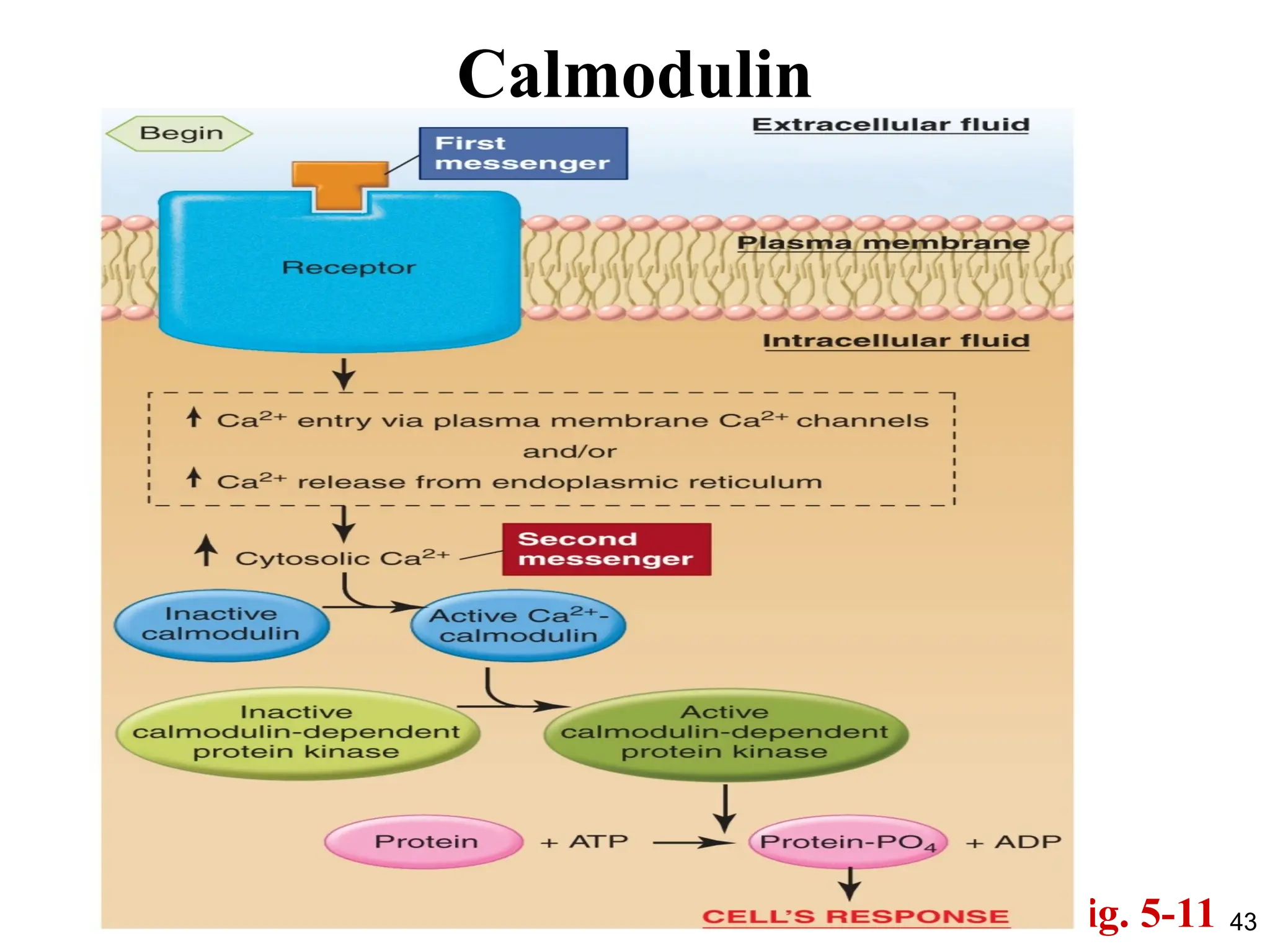
Ca2+
as a SecondMessenger
• Besides the changes to the membrane potential, Ca2+
also acts by
its ability to bind to various cytosolic proteins, altering their
conformation and thereby activating their function.
• One of the most important of these is a protein, found in virtually
all cells, known as calmodulin.
• On binding with Ca2+
calmodulin changes shape, and this allows
calcium-calmodulin to activate or inhibit a large variety of
enzymes and other proteins, many of them protein kinases.
• Other proteins Ca2+
binds include: troponin, nitric oxide synthase,
PYK2.
44
Cessation of Activityin Signal Transduction Pathways
• Once initiated, signal transduction pathways are eventually shut off
because chronic overstimulation of a cell can in some cases be
detrimental.
• The key event is usually the cessation of receptor activation. Responses to
messengers are transient events that persist only briefly, and subside when
the receptor is no longer bound to the ligand.
• A major way that receptor activation ceases is by a decrease in the
concentration of first messenger molecules in the region of the receptor.
• This occurs as enzymes in the vicinity metabolize the first messenger, as
the first messenger is taken up by adjacent cells, or as it simply diffuses
away.
45.
45
Cessation of Activityin Signal
Transduction Pathways
• Receptors can be inactivated in at least three other ways:
1. The receptor becomes chemically altered (usually by
phosphorylation), which may lower its affinity for a first
messenger, and so the messenger is released.
2. Phosphorylation of the receptor may prevent further G protein
binding to the receptor.
3. Plasma membrane receptors may be removed when the
combination of first messenger and receptor is taken into the
cell by endocytosis.
46.
46
Interactions of SignalTransduction Pathways
• It is essential to recognize that the pathways do not exist in
isolation but may be active simultaneously in a single cell,
undergoing complex interactions.
• This is possible because a single first messenger may trigger
changes in the activity of more than one pathway and, much more
importantly, because many different first messengers—often
dozens—may simultaneously influence a cell.
• Moreover, a great deal of “cross-talk” can occur at one or more
levels among the various signal transduction pathways. For
example, active molecules generated in the cAMP pathway can
alter the of receptors and signaling molecules generated by other
pathways.