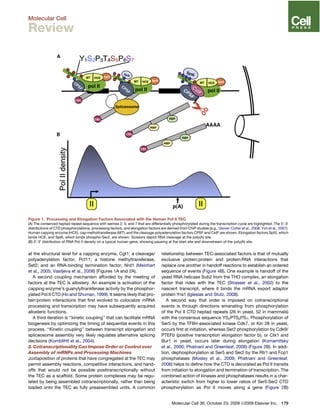
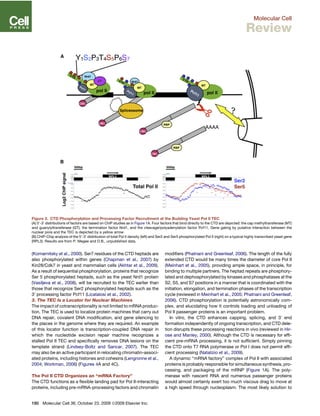
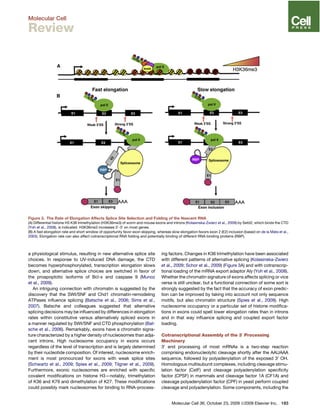
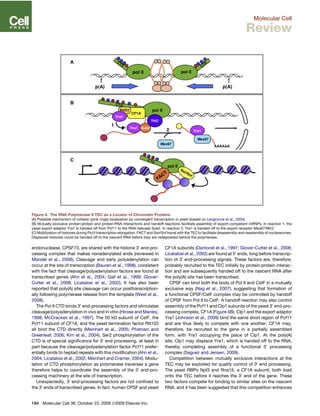
This document discusses how the transcription elongation complex (TEC), comprising the growing nascent RNA and RNA polymerase traversing chromatin, serves as a nexus for various nuclear processes including mRNA biogenesis, DNA repair, chromatin modification, and gene silencing. The TEC allows these processes to occur cotranscriptionally by localizing various protein complexes and factors to the site of transcription. In particular, the phosphorylated C-terminal domain of RNA polymerase II functions as a landing pad that recruits mRNA processing factors and chromatin modifiers to couple transcription with downstream events. This cotranscriptional coupling enhances the efficiency and coordination of gene expression and nuclear transactions organized around the transcription machinery.