Baculovirus and Insect Cell Expression Protocols 3rd Edition David W. Murhammer (Ed.)
Baculovirus and Insect Cell Expression Protocols 3rd Edition David W. Murhammer (Ed.)
Baculovirus and Insect Cell Expression Protocols 3rd Edition David W. Murhammer (Ed.)



















![3
David W. Murhammer (ed.), Baculovirus and Insect Cell Expression Protocols, Methods in Molecular Biology,
vol. 1350, DOI 10.1007/978-1-4939-3043-2_1, © Springer Science+Business Media, LLC 2016
Chapter 1
Useful Tips, Widely Used Techniques, and Quantifying
Cell Metabolic Behavior
David W. Murhammer
Abstract
The insect cell culture/baculovirus system has three primary applications: (1) recombinant protein synthesis,
(2) biopesticide synthesis, and (3) as a model system (e.g., for studying apoptosis). The fundamental tech-
niques involved in these applications are described throughout this book. In this chapter the most widely
used techniques are summarized and the reader is directed to detailed information found elsewhere in this
book. Furthermore, many useful tips and my personal preferences that are rarely published are discussed
in this chapter along with quantitative methods to characterize cell growth, baculovirus infection, and
metabolism.
Key words Cell growth, Baculovirus infection, Population doubling time, Exponential growth, Cell
metabolism, Specific utilization rates, Specific production rates
1 Introduction
The most commonly used lepidopteran insect cell lines are isolates
from Spodoptera frugiperda (Sf-9 and Sf-21) and Trichoplusia ni
(Tn-5, which is commonly known as BTI-Tn-5B1-4 and is com-
mercially known as High Five™ cells (Invitrogen); see Note 1).
See Chapter 6 for an extensive list of available insect cell lines and
Chapter 7 for methods involved in new cell line development. The
Sf-9, Sf-21, and Tn-5 cells grow optimally at 27–28 °C and do not
require CO2 (due to the buffer system used in the medium). Thus,
temperature is the only parameter that needs to be controlled in
incubators used to grow these cells.
Following subculture, insect cells in batch culture (see Note 2)
proceed through the typical growth phases [1] (see Note 3 and
Fig. 1): (1) lag phase (see Note 4), (2) exponential growth phase
(see Note 5), (3) stationary phase (see Note 6), and (4) decline
(or death) phase. The lag phase is the time period between sub-
culturing and the exponential growth phase in which the cell
growth rate reaches its maximum. Lag phase is usually caused by
1.1 Cell Growth](https://image.slidesharecdn.com/84287-250511220407-b12cb833/85/Baculovirus-and-Insect-Cell-Expression-Protocols-3rd-Edition-David-W-Murhammer-Ed-19-320.jpg)
![4
an environmental change, e.g., using cells from the stationary
phase to start a new culture [1] (see Note 7). During the station-
ary phase the net growth rate is zero, i.e., the cell growth rate is
not necessarily zero, but it is equal to the cell death rate. Finally,
during the decline phase the cell death rate exceeds the cell
growth rate and the viable cell number declines.
The major parameters that are used to characterize cell
growth are specific growth rate (μ), population doubling time
(PDT), lag time, and maximum cell density. All of these parame-
ters are functions of the specific cell line and growth environment
(dissolved oxygen concentration, nutrient concentrations, temper-
ature, pH, etc.). Furthermore, % cell viability is a measure of cell
quality. Typical ranges of μ and PDT are 0.029–0.035 h−1
and
20–24 h, respectively, for the Sf-9, Sf-21, and Tn-5 cell lines [2].
The lag phase can be eliminated by using cells from the exponen-
tial growth phase (see Note 4). Finally, the maximum cell density
in batch culture varies considerably depending upon the cell line
and the medium composition, and can exceed 107
cells/mL in
some cases [3]. Additional details about these parameters are given
under Subheading 3.1.
Cell growth during the exponential growth phase can be rep-
resented by
N N t
= 0 exp( ),
m (1)
where N=cell density (cells/mL) at a given time t, N0 =cell density
(cells/mL) at time t=0, μ=specific growth rate (h−1
), and t=time
in culture (h).
Stationary
Phase
Decline
Phase
Exponential
Phase
Lag
Phase
L
Log
(viable
cell
density)
Time
Fig.1A typical cell growth curve (cell density vs.time) showing the lag,exponential,
stationary, and decline phases. The length of the lag phase (L) can be found by
extending the lag and exponential phase lines and finding the time at which they
intersect (as shown)
David W. Murhammer](https://image.slidesharecdn.com/84287-250511220407-b12cb833/85/Baculovirus-and-Insect-Cell-Expression-Protocols-3rd-Edition-David-W-Murhammer-Ed-20-320.jpg)
![5
Insect cells are infected with either a recombinant baculovirus (see
Chapter 4) to produce a recombinant protein or with a wild-type
baculovirus to produce biopesticides (recombinant baculovirus can
also be used as biopesticides, see Chapters 19 and 21). In all of these
cases it is critical that a high quality baculovirus (i.e., a pure baculo-
virus absent of mutants, see Subheading 3.2) and healthy cells (i.e.,
viability>95 %) in exponential growth be used. The most impor-
tant parameter involved in baculovirus infection is the multiplicity
of infection (MOI), which is the ratio of infectious baculovirus par-
ticles to cells. Details about the MOI and other important issues
involved in baculovirus infection are given under Subheading 3.2.
Cell metabolism involves characterizing how cells grow, utilize
nutrients, and produce products and byproducts. Cell growth, as
indicated under Subheadings 1.1 and 3.1, can be characterized
by μ, PDT, lag time, and maximum cell density. Other parame-
ters that can be used to characterize cell metabolism are specific
utilization (for nutrients) and specific production (for byproducts)
rates. For example, we found the specific utilization rates for
glucose in uninfected Sf-9 and Tn-5 cells to be −2.4 × 10−17
and
−2.8 × 10−17
mol/cell-s (the negative sign in indicative of con-
sumption that results in a reducing the glucose concentration),
respectively [2]. Furthermore, it was found that Sf-9 cells did
not produce measurable amounts of lactate and ammonium ions
when sufficient dissolved oxygen (DO) was present. In contrast,
the uninfected Tn-5 cells produced lactate and ammonium ions
at rates of 0.7 × 10−17
and 5.1 × 10−17
mol/cell-s, respectively,
when sufficient DO was present. Both the Sf-9 and Tn-5 cells
produced alanine (another common byproduct in insect cell cul-
ture) at rates of 1.1 × 10−17
and 1.5 × 10−17
mol/cell-s, respec-
tively. Information about other specific uptake and production
rates in these cell lines, both in uninfected and infected cells, can
be found in Rhiel et al. [2]. Details about evaluating these
parameters are given under Subheading 3.3.
2 Materials
1. Supplies and equipment listed in Chapters 9 and 10 for small-
scale cultures or Chapters 11 and 12 for large-scale culture.
2. Coulter particle counter/sizer (Beckman Coulter, Inc.,
Fullerton, CA) (see Note 8).
1. Supplies and equipment listed in Chapter 10 for small-scale
cultures or Chapters 11 and 12 for large-scale culture.
2. Coulter particle counter/sizer (Beckman Coulter, Inc.,
Fullerton, CA) (see Note 8).
1.2 Baculovirus
Infection
1.3 Cell Metabolism
2.1 Cell Growth
2.2 Baculovirus
Infection
Widely Used Techniques](https://image.slidesharecdn.com/84287-250511220407-b12cb833/85/Baculovirus-and-Insect-Cell-Expression-Protocols-3rd-Edition-David-W-Murhammer-Ed-21-320.jpg)
![7
underlying principle is that the membranes of the dead cells will be
leaky and therefore will readily take up the blue dye and appear blue
under the microscope. In contrast, viable (i.e., live) cells have intact
membranes that will exclude the dye. While trypan blue dye exclu-
sion is commonly referred to as a measure of cell viability, it is more
accurately a measure of membrane integrity. Nonetheless, this is by
far the most commonly used method to determine cell viability due
to its simplicity and the fact that results can be obtained in ~5 min.
The MTT (3-(4,5-
dimethylthiazolyl-2)-2,5-diphenyltetrazolium
bromide) cell proliferation assay [4] is a more elaborate assay that
can be used to provide a more authentic measure of cell viability.
The principle behind this assay is that the yellow tetrazolium MTT
is reduced by metabolically active cells to produce purple formazan
that can be quantified by spectroscopic methods. MTT cell prolif-
eration assay kits are available through many vendors, e.g., the
American Type Culture Collection (Manassas, VA; http:/
/www.
atcc.org).
I prefer using a Coulter particle counter (see Note 8) and the
trypan blue dye exclusion method for determining the total cell
density and cell viability, respectively. (Unfortunately, the Coulter
particle counter is a relatively expensive piece of equipment that
may be beyond the financial means of some laboratories.) The via-
ble cell density can then be determined by multiplying the total cell
density by the fraction of viable cells. Manufacturer instructions
can be followed to determine cell densities (and mean cell size)
with the Coulter particle counter.
Conducting cell counts in suspension cultures (i.e., spinner
flasks, shaker flask, and bioreactors) is much easier than in attached
cultures (i.e., tissue culture flasks). Specifically, cell samples can be
directly withdrawn from the cultures at the desired times, i.e.,
every 12–24 h, and continued until the cells are at least in the sta-
tionary growth phase (Fig. 1). In contrast, when conducting cell
counts for monolayer cultures, the cells must first be detached
from the growth surface (see Subheading 3.1 in Chapter 9). When
obtaining a growth curve for cells in monolayer culture the follow-
ing procedure should be followed.
1. Equally seed a number of tissue culture flasks equal to the
number of time points at which cell counts will be taken
(see Note 11).
2. Detach cells from one tissue culture flask.
3. Determine the cell density (cells/mL) and viability.
4. Repeat steps 2 and 3 at each time point at which a cell count
is to be taken. There should be a sufficient number of flasks
seeded to obtain a growth curve into the stationary growth
phase (Fig. 1).
Widely Used Techniques](https://image.slidesharecdn.com/84287-250511220407-b12cb833/85/Baculovirus-and-Insect-Cell-Expression-Protocols-3rd-Edition-David-W-Murhammer-Ed-23-320.jpg)



![12
(GFP) (Subheading 3.2 in Chapter 22). The anti-gp64 antibody
procedure is based on the principle that the gp64 protein is
expressed on the surface of baculovirus infected cells. The GFP
procedure is based on detecting GFP (this method requires insert-
ing the GFP gene into the baculovirus genome as described under
Subheading 3.1 in Chapter 22) following baculovirus infection.
Another method of tittering baculovirus is through flow cytome-
try following binding of a fluorescent dye to baculovirus DNA (see
Note 20 in Chapter 11). This procedure provides a total baculo-
virus concentration in contrast to the plaque assay and end point
dilution procedures that provide the desired infectious baculovi-
rus concentration.
The baculovirus tittering methods described above can best be
verified by infecting cell cultures at a range of concentrations and
observing cell growth or the lack thereof (see Subheadings 3.2.4
and 3.2.6).
The Poisson distribution [5] expresses the probability of a number
of events occurring during a given time period and can be used to
model the baculovirus infection process. First, the fraction of cells
infected by a specific number of baculovirus particles at a given
MOI can be predicted from
F n
n
n
( )
( ) exp( )
!
,
,MOI
MOI MOI
=
-
(4)
where F(n, MOI) is the fraction of cells infected with n baculovirus
particles when a multiplicity of infection of MOI is used.
The fraction of baculovirus infected cells can be predicted by
subtracting the fraction of uninfected cells (i.e., using n=0 in
Eq. 4) from 1.0:
( )
Fraction of insect cells infected with Baculovirus at a given MOI
= 1
1- -
exp( ).
MOI
(5)
Some issues to consider when infecting an insect cell culture
include (1) the cell line to use, (2) the cell density at which the cells
should be infected, (3) the MOI to use, and (4) the product har-
vest time.
Sf-9 and Tn-5 are the most commonly used host cell lines
used to express recombinant proteins with the BEVS. It has been
demonstrated that the Tn-5 cell line expresses higher levels of
recombinant proteins in many, but not all, cases [6] and therefore
is usually a good choice for producing recombinant proteins. The
Mimic™ Sf-9 cell line commercially available from Life
Technologies (see Note 14) is a good choice if the recombinant
protein is glycosylated. The reader is also referred to Chapter 6
regarding the wide range of available insect cell lines and Chapter 7
regarding methods to develop new cell lines, as well as Chapter 18
3.2.4 Poisson
Distribution
3.2.5 Infection of Insect
Cell Cultures: Cell Line
Selection
David W. Murhammer](https://image.slidesharecdn.com/84287-250511220407-b12cb833/85/Baculovirus-and-Insect-Cell-Expression-Protocols-3rd-Edition-David-W-Murhammer-Ed-28-320.jpg)



![16
is in the gas phase at 27 °C is ~0.26 mM [7]. Thus, 100 % air
saturation corresponds to ~0.26 mM oxygen. If desired, cor-
rections can be made for the effect of salts (i.e., actual medium
vs. pure water) in the solution on oxygen solubility [8]; how-
ever, the correction is usually small and can be ignored for
most applications.
6. Example: assume that we find a slope, i.e., a volumetric OUR,
of −2.8 % DO/min from step 3 at a cell density of 2.5×106
cell/mL; the specific OUR can then be calculated:
( )
.
min
min .
Specific OUR
%DO
s
mM
%DO
=
-
æ
è
ç
ö
ø
÷
æ
è
ç
ö
ø
÷
æ
è
ç
ö
2 8
60
0 26
100 ø
ø
÷
æ
è
ç
ö
ø
÷
´
( )( )
= - ´ -
mol L
mM
cells mL mL L
m
/
. / /
.
10
2 5 10 1000
4 9 10
3
6
17
o
ol cell s
-
/
1. If a DO electrode is not present, then a DO monitor can be
used.
2. Remove the appropriate volume of cell suspension from the
culture (appropriate for the DO monitor chamber being used)
and place in the DO monitor chamber.
3. Record the DO concentration as a function of time. Note that
the DO concentration can be monitored to lower DO concen-
trations than in cell culture (Subheading 3.3.1) since the cells
will be discarded when the test is completed and it does not
matter if the cell metabolism is affected.
4. Follow steps 2–5 given under Subheading 3.3.1 to determine
the specific OUR.
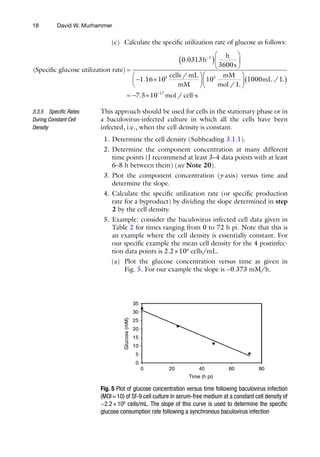
As shown below, the methods used to calculate the specific nutri-
ent utilization and byproduct accumulation rates are different dur-
ing exponential growth than when the cell density is constant (i.e.,
in stationary growth phase and in baculovirus infected cultures).
1. Determine the viable cell density (Subheading 3.1.1) and
nutrient (e.g., glucose) and/or byproduct (e.g., lactate ion)
concentration as a function of time in culture (see Note 20).
2. Determine the range of times over which cells are in the expo-
nential phase.
3. Determine the specific growth rate (Subheading 3.1.2).
4. Plot cell density (y-axis) versus component of interest concen-
tration (x-axis) using exponential phase data only and deter-
mine the slope. For a nutrient this slope is referred to cell yield
coefficient.
5. The specific nutrient utilization rate (or specific byproduct
production rate) is then evaluated by dividing the specific
3.3.2 Specific Oxygen
Utilization Rate: Using
a Dissolved Oxygen (DO)
Monitor
3.3.3 Specific Nutrient
Utilization and Byproduct
Accumulation Rates
3.3.4 Specific Rates
During Exponential
Growth Phase
David W. Murhammer](https://image.slidesharecdn.com/84287-250511220407-b12cb833/85/Baculovirus-and-Insect-Cell-Expression-Protocols-3rd-Edition-David-W-Murhammer-Ed-32-320.jpg)




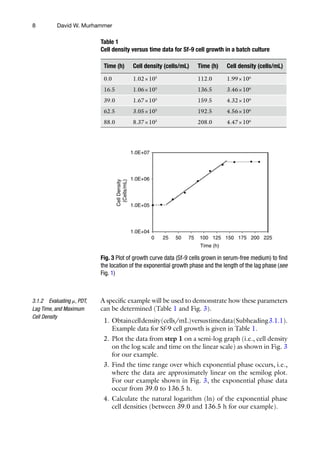
![22
20. Many of the nutrients and byproducts can be determined using
a bioprofile analyzer by following the manufacturer directions.
Furthermore, an HPLC can be used to determine amino acid
concentrations as described in reference [2].
References
1. Pirt SJ (1975) Principles of microbe and cell cul-
tivation. Wiley, New York
2. Rhiel M, Mitchell-Logean CM, Murhammer
DW (1997) Comparison of Trichoplusia ni BTI-
Tn-5B1-4 (High Five™) and Spodoptera frugi-
perda Sf-9 insect cell line metabolism in
suspension cultures. Biotechnol Bioeng 55:
909–920
3. Schmid G (1996) Insect cell cultivation: growth
and kinetics. Cytotechnology 20:43–56
4. Hansen MB, Nielsen SE, Berg K (1989)
Re-examination and further development of a
precise and rapid dye method for measuring cell
growth/cell kill. J Immunol Methods 119:
203–210
5. Devore JL (2004) Probability and statistics
for engineering and the sciences, 6th edn.
Brooks/Cole – Thomson Learning, Belmont,
CA
6. Davis TR, Wickham TJ, McKenna KA et al
(1993) Comparative recombinant protein pro-
duction of eight insect cell lines. In Vitro Cell
Dev Biol Anim 29:388–390
7. Perry RH, Green DW (eds) (1997) Perry’s
chemical engineers’ handbook, 7th edn.
McGraw-Hill, New York
8. Onda K, Sada E, Kobayashi T et al (1970)
Salting-out parameters of gas solubility in aque-
ous salt solutions. J Chem Eng (Japan)
3:18–24
David W. Murhammer](https://image.slidesharecdn.com/84287-250511220407-b12cb833/85/Baculovirus-and-Insect-Cell-Expression-Protocols-3rd-Edition-David-W-Murhammer-Ed-38-320.jpg)



![26
The many commercial kits now available have brought the system
within the capabilities of any competent molecular biologist. Most
ofthesekitsarebasedontheprototypememberoftheBaculoviridae,
namely Autographa californica multiple nucleopolyhedrovirus
(AcMNPV) and utilize the highly expressed polyhedrin gene pro-
moter. The simplicity of culturing insect cells such as Spodoptera
frugiperda or Trichoplusia ni makes scale up of recombinant pro-
tein production feasible for most laboratories. The problems expe-
rienced with spinner or suspension cultures where shear forces
limited viability have largely been solved with the introduction of
serum-free media allied with antifoam and protective (e.g.,
Pluronic®
F-68) agents. The exploitation of baculoviruses as bio-
safe insecticides has also benefited from the work on expression
vectors and fundamental studies on virus gene function. While
most baculovirus expression vectors lack the original polyhedrin
gene required for making occluded viruses, recombinant virus
insecticides can preserve this process by utilizing nonessential
regions of the virus genome for the insertion of foreign genes
encoding insecticidal proteins.
2 Classification
The Baculoviridae are a family of DNA viruses with circular dou-
ble stranded genomes that only infect arthropods. They are char-
acterized by their ability to occlude virions in a crystalline protein
matrix to form either polyhedra or granules. Viruses forming
polyhedra are known as nucleopolyhedroviruses (NPVs) and
those forming granules as granuloviruses. This subdivision is
based on a number of criteria, including occlusion body mor-
phology and the mechanism by which nucleocapsids are envel-
oped in infected cells [1]. Granuloviruses produce small occlusion
bodies (OBs) (0.16−0.30 μm×0.30−0.50 μm) normally con-
taining one or occasionally two virions encapsulated in a protein
called granulin. Nucleopolyhedroviruses produce larger occlu-
sion bodies (0.5–1.5 μm in diameter) composed of polyhedrin
protein, which contain many virions. The family is divided into
four genera that only infect arthropods. The Alphabaculoviruses
and Betabaculoviruses comprise nucleopolyhedroviruses (NPVs)
and granuloviruses (GVs) that are isolated from Lepidoptera
(butterflies and moths). Figure 1 illustrates an example of each of
these genera. Deltabaculoviruses and Gammabaculoviruses com-
prise NPVs and infect dipteran (flies) and hymenopteran (sawflies)
species, respectively. The NPVs pathogenic for members of the
order Lepidoptera have been further subdivided into groups I
and II based on molecular phylogenies [1, 2]. An occluded virus
also infects the pink shrimp Penaeus monodon [3–5]. Partial
sequence analysis showed that it encodes proteins similar to those
produced by baculoviruses.
Barbara J. Kelly et al.](https://image.slidesharecdn.com/84287-250511220407-b12cb833/85/Baculovirus-and-Insect-Cell-Expression-Protocols-3rd-Edition-David-W-Murhammer-Ed-42-320.jpg)
![27
3 Baculovirus Structure
The double stranded, supercoiled, circular DNA genome of
AcMNPV is highly condensed within a nucleocapsid. This con-
densed packaging is facilitated by the core protein p6.9, a 54
amino acid protein rich in arginine [6]. A major protein of the
capsid, vp39, has also been identified. This 39 kDa protein has
been found to be distributed randomly over the surface of the
nucleocapsid [7].
There are two types of baculovirus progeny, budded virus
(BV) and occlusion-derived virus (ODV), which are genetically
identical [8]. However, there are differences in morphology, tim-
ing and cellular site of maturation, structural proteins, source of
viral envelopes, antigenicity, and infectivity [9–13].
BV particles possess spike-like structures known as peplomers,
composed of the glycoprotein GP64 for group I NPVs, at one end
of the virion [14]. The GP64 protein is incorporated throughout
the virus envelope, albeit at lower concentrations than at the
peplomers [14]. During infection, GP64 localizes to discrete areas
of the plasma membrane at which points budding of virions takes
place [15]. Thus, as BV particles bud from the plasma membrane
they acquire a plasma membrane-derived envelope containing the
GP64 glycoprotein.
Granuloviruses and group II NPVs lack GP64 but instead
encode a homolog of another envelope protein, LD130 [16].
While the LD130-encoding baculoviruses do not contain gp64
Fig. 1 Electronmicrographs of Autographa californica multiple nucleopolyhedrovirus (AcMNPV) polyhedron (a)
and Plodia interpunctella granulovirus (PiGV) granules (b). Note the multiple virus particles (vps) in longitudinal
or cross section in AcMNPV, whereas in PiGV each granule contains a single virus particle (vp). Virus particles
are occluded within polyhedrin protein (P) or granulin protein (G). Polyhedra are bounded by an envelope (PE),
which contains protein and carbohydrate. Scale bar is 200 nM
Baculovirus Molecular Biology](https://image.slidesharecdn.com/84287-250511220407-b12cb833/85/Baculovirus-and-Insect-Cell-Expression-Protocols-3rd-Edition-David-W-Murhammer-Ed-43-320.jpg)
![28
homologs, those encoding GP64 also contain a version of LD130.
The OpMNPV LD130 homolog was found to be a glycosylated
protein associated with BV particles and the plasma membrane of
cells at similar locations as the GP64 protein [17].
One of the major differences between BV and ODV is that the
latter do not contain GP64 or LD130. Instead a virus-encoded
envelope protein P74 is associated with the outside of the virus
[18, 19]. The presence of P74 on the outside of the virion enve-
lope suggests that it may play a role in ODV attachment to midgut
cells. Another structural protein of the ODV envelope of Spodoptera
littoralis (Spli) NPV has been identified. The gene encoding this
protein was termed per os infectivity factor (pif) and is homologous
to ORF 119 of AcMNPV [20]. A third gene product inferred to
be a structural protein is encoded by pif-2, which was identified in
Spodoptera exigua NPV and is present in AcMNPV as ORF22 [21].
Recent studies have also identified pif-4 [22] and pif-5 [23, 24] as
additional envelope proteins. The PIFs form a complex on the sur-
face of ODV [25].
Other ODV-specific envelope proteins include ODV-E25
[26], ODV-E66 [27], ODV-E56 [28], ODV-E18, and ODV-E35
[29]. These proteins, along with P74, have been found to associate
with intranuclear microvesicle structures, which appear in the
nucleus during infection [29, 30]. This association has led to the
suggestion that these microvesicles play a role in ODV envelop-
ment [27, 28]. Invagination of the inner nuclear membrane
observed during baculovirus infection has raised the possibility
that it may act as the source of the microvesicles [31–33].
Another ODV-specific protein, GP41, has been identified as
an O-linked glycoprotein, predicted to localize to the region
between the virus envelope and the nucleocapsid, an area referred
to as the tegument [34]. Although GP41 is not present in the bud-
ded form of the virus, it has been shown to be required for release
of nucleocapsids from the nucleus during BV production [35]. An
additional protein, ODV-EC27, has been identified as a structural
protein of the ODV envelope and capsid, and may be present in a
modified form in BV [29]. This protein may be a cyclin homolog,
involved in host cell cycle arrest during baculovirus infection [36].
An envelope protein of both BV and ODV has also been identified
(BV/ODV-E26) and found to be associated with intranuclear
microvesicles [37]. A seminal study of protein composition of
ODV was conducted by using a combination of techniques [38].
Screening expression libraries with antibodies generated to ODV
or BV and mass spectroscopic (MS) analysis of ODV protein
extracts identified up to 44 potential components unique to
ODV. This topic was revisited recently when the protein content of
BV was reexamined [39].
The major component of the NPV occlusion body is the
29 kDa polyhedrin protein. While the protein is thought to be small
Barbara J. Kelly et al.](https://image.slidesharecdn.com/84287-250511220407-b12cb833/85/Baculovirus-and-Insect-Cell-Expression-Protocols-3rd-Edition-David-W-Murhammer-Ed-44-320.jpg)
![29
enough to diffuse through nuclear pores, a nuclear localization
signal (KRKK) has been identified at amino acids 32–35 [40].
Another region of the protein (amino acids 19–110) is required for
assembly into large occlusion-like particles [40]. The polyhedrin
protein is resistant to solubilization except under strongly alkaline
conditions and functions to shield virions from physical and bio-
chemical decay while outside the insect host, as well as protecting
against proteolytic decay during late stages of infection [41].
Surrounding the polyhedral matrix of a mature occlusion body
is the polyhedron envelope (PE), also known as the polyhedron
membrane or calyx. The PE was originally reported to be com-
posed of carbohydrate [42]; however, a 34 kDa phosphorylated
protein (PP34) has since been found to be covalently bound to the
PE of AcMNPV via a thiol linkage [43]. The polyhedron envelope
is thought to increase the stability of the occlusion body in the
environment and has been found to protect occlusion bodies from
physical stress [43, 44]. A recombinant virus unable to produce
PP34 was found to be deficient in polyhedron envelope formation
and electron dense spacers, which are thought to be precursors of
the envelope structure [44, 45].
Both electron dense spacers and the polyhedron envelope have
been found in association with fibrillar structures composed of the
P10 protein [44, 46]. This protein, like polyhedrin, is produced in
large quantities during the occlusion phase. The association of the
polyhedron envelope with the P10-containing fibrillar structures
suggests a role for the protein in assembly and proper association
of the polyhedron envelope around the occlusion body matrix
[46]. Comparative analysis of P10 protein sequences revealed that
they appear to have an amphipathic alpha-helical terminus that
condenses as coiled-coil multimers [47]. The condensation of P10
monomers to coiled-coil multimers may be a step leading to fibrous
body formation in virus-infected cells.
4 Baculovirus Replication In Vivo
Within a cell, baculoviruses have a biphasic cycle of replication pro-
ducing two structurally distinct viral phenotypes responsible for
specialized functions within the insect host. Occlusion-derived
virus (ODV) initiates the primary infection in the gut epithelial
cells, while budded virus (BV) is responsible for secondary rounds
of infection in the insect host. Infection begins with ingestion of
occlusion bodies by a suitable insect host larval stage, followed by
dissolution of the occlusion body matrix in the alkaline midgut.
Occlusion-derived virus is released from dissolved occlusion bod-
ies, passes through the peritrophic membrane of the midgut and
infects the columnar epithelial cells that border the gut lumen.
It has been suggested that entry occurs by direct membrane fusion,
Baculovirus Molecular Biology](https://image.slidesharecdn.com/84287-250511220407-b12cb833/85/Baculovirus-and-Insect-Cell-Expression-Protocols-3rd-Edition-David-W-Murhammer-Ed-45-320.jpg)
![30
involving interaction between virion surface proteins and midgut
cell surface proteins [48]. Removal of P74, PIF1 or PIF2 from
virus particles by deletion of the appropriate gene has been dem-
onstrated to prevent infection of insect larvae via the oral route
[18–21]. Deletion of pifs-3-5 has a similar effect [22–24, 49].
Midgut regenerative cells have also been found to be sites of pri-
mary infection, albeit at a lower frequency than columnar cells
[50]. After replication in the epithelial cells, virus buds in a polar
manner, exiting the basal membrane into the hemolymph, thereby
allowing the infection to spread throughout the insect host. Early
synthesis of the BV envelope fusion protein GP64 seems to allow a
“pass through” strategy for the virus, whereby uncoated virus
nucleocapsids can rapidly exit the newly infected midgut cells to
accelerate the onset of systemic infections [51]. The insect tracheal
system has been identified as the main route used by the virus to
spread from one tissue to another [52]. Efficient BV formation in
AcMNPV requires Ac92 and Ac79 [53, 54].
In addition to enlargement of the nucleus upon baculovirus
infection, cells become rounded due to rearrangement of the cyto-
skeleton. A distinct structure termed the virogenic stroma develops
in the nucleus. This is a chromatin-like network of electron-dense
filaments. It is the predominant structure in the nucleus from 8 to
48 h postinfection (h pi) [55], and is thought to be the site of viral
DNA replication and late gene transcription, as well as nucleocap-
sid formation [56]. Once assembled, nucleocapsids are released
from the nucleus, gaining a nuclear membrane-derived envelope
that is subsequently lost during transit through the cytoplasm. In
the case of group I NPVs, the envelope glycoprotein GP64 local-
izes to discrete areas of the plasma membrane during infection and
it is at these sites that budding of nucleocapsids takes place [15].
The BV envelope is therefore derived from the plasma membrane
and contains the viral glycoprotein GP64. Group II NPVs, and
GVs, encode homologs of the envelope fusion (F) protein LD130
[16], which associate with BV particles, and the plasma membrane
of infected cells, at similar positions to GP64 in group I NPV-
infected cells [17]. These F proteins are functionally analogous to
GP64 since they can restore infectivity to mutant viruses lacking
gp64 [57]. GP64 is essential for cell-to-cell BV transmission, since
AcMNPV particles containing the protein, but not those lacking
the protein, are able to disseminate virus to other cells [58].
Later in infection (~24 h pi), BV production is reduced and
nucleocapsids are transported to sites of intranuclear envelopment
and incorporation into OBs [44]. The source of the ODV enve-
lope is unclear. A number of ODV envelope proteins have been
found to associate with intranuclear microvesicle structures that
appear in the nucleus during infection, thereby suggesting they
may be the source of the envelope [27–29].
Barbara J. Kelly et al.](https://image.slidesharecdn.com/84287-250511220407-b12cb833/85/Baculovirus-and-Insect-Cell-Expression-Protocols-3rd-Edition-David-W-Murhammer-Ed-46-320.jpg)
![31
Occlusion bodies accumulate until the terminal stages of infection
when the insect liquefies and the occlusion bodies are released
into the environment. Liquefaction appears to be caused by the
production of both virus-encoded chitinase and cathepsin genes
[59–61] and may also involve the fp25k product [62]. Deletion of
the cathepsin gene from Bombyx mori NPV reduced recombinant
protein degradation in virus-infected insects, presumably because
of the reduction in proteinase activity [63]. Just prior to death
many Lepidopteran species crawl to the top of the vegetation on
which they were feeding and hang from this elevated position,
facilitating dissemination of the virus as the cadaver decomposes
[64]. This enhanced locomotory activity (ELA) may well be virus-
induced, since insects infected with a B. mori NPV mutant lacking
the protein tyrosine phosphatase gene showed dramatically reduced
ELA before death after about 5 days [65]. However, removal of
ecdysteroid glucosyltransferase gene (egt) from a baculovirus was
found to reduce vertical movement of virus-infected insects on
plants [66]. Occlusion bodies serve as survival vehicles for the virus
when outside the insect host, as well as acting as dispersal agents
between individual insects. They may also protect the virus against
proteolytic decay during the end stages of infection [41].
5 Baculovirus Gene Expression and Replication
The complete sequence of AcMNPV clone 6 has been determined.
The original analysis of the 133,894 bp genome suggested that
the virus encodes 154 methionine-initiated, potentially expressed
open reading frames (ORFs) of 150 nucleotides or more [67].
However, resequencing of various regions of the genome and
comparison with other virus isolates has corrected a number of
errors and suggests that AcMNPV may only encode 150 genes.
Other baculoviruses have also been sequenced. An up-to-date
summary of completed genomes is available from the National
Center for Biotechnology Information (NCBI). Baculovirus gene
expression is divided into four temporal phases: immediate-early,
delayed-early, late, and very late, although the first two are often
considered as one. Immediate-early genes are distinguished from
delayed-early by their expression in the absence of de novo protein
synthesis. Expression of delayed-early genes, however, appears to
be dependent on the presence of immediate-early gene products.
Transcription of late genes occurs concurrently with the onset of
viral DNA replication at about 6 h pi. Very late gene transcription
begins at about 20 h pi and involves high levels of expression from
the polyhedrin and p10 promoters, two proteins involved in the
occlusion of virions. While levels of late gene mRNA transcripts
decrease at very late times in infection, very late polyhedrin and
p10 gene transcript levels continue to remain high [68].
Baculovirus Molecular Biology](https://image.slidesharecdn.com/84287-250511220407-b12cb833/85/Baculovirus-and-Insect-Cell-Expression-Protocols-3rd-Edition-David-W-Murhammer-Ed-47-320.jpg)
![32
Analysis of the AcMNPV genome has revealed that RNA
transcripts are not clustered according to their temporal expres-
sion. Instead, early and late genes are found distributed through-
out the genome. All AcMNPV RNAs are both 5′ methyl capped
and 3′ polyadenylated [69, 70]. Only one transcript, which is that
of the immediate-early gene ie-1, is known to be spliced [71].
However, transcriptional units involving overlapping RNAs have
been identified in the AcMNPV genome, which may provide an
alternative means of introducing a variety of expression [72, 73].
Overlapping transcripts composed of early and late RNAs with a
common 3′ end have been identified in a number of regions [72,
73]. The HindIII-K fragment of the AcMNPV genome produces
five overlapping RNAs, two immediate-early, one delayed-early,
and two late gene transcripts, transcribed in the same direction and
terminating at a common 3′ end [73]. This arrangement has been
implicated in the temporal regulation of these genes. It has been
suggested that the longer, later 5′ extended transcripts serve to
repress transcription of earlier genes, located downstream, proba-
bly by means of promoter occlusion, as well as acting as mRNAs
for late viral products [73].
The polyhedrin and p10 genes are also transcribed as several
overlapping RNAs. In the case of these two genes, however, tran-
scripts have common 5' ends, with longer RNAs being derived
from read through of termination signals at the 3′ end of the
smaller RNAs [72–74]. Four overlapping transcripts were mapped
to the p10 gene region and were found to comprise two sets: a late
phase pair of transcripts (1100 and 1500 bases) sharing a common
5′ end that are most abundant at 12 h pi and a very late phase pair
(750 and 2500 bases) that also have a common 5′ end and are
most abundant at 24 h pi [74]. Promoter occlusion may also be
operating in this region. Synthesis of earlier transcripts may prevent
RNA polymerase initiation at the p10 promoter located down-
stream. At later times, when transcription from upstream promot-
ers has ceased, RNA polymerase may be able to initiate at the
downstream p10 promoter [74].
Baculovirus early genes are transcribed before the onset of viral
DNA replication. Expression of many early genes begins
immediately after cell infection, with some transcripts, for exam-
ple the anti-apoptotic p35 gene early mRNAs, appearing within
the first 2 h [75]. Genes in this phase of the virus lifecycle are
transcribed by the host RNA polymerase II, as demonstrated by
the fact that early transcription is sensitive to α-amanitin [76], a
compound that binds to and inactivates the large subunit of RNA
polymerase II.
Most early baculovirus promoters contain a TATA element which, as
well as regulating the rate of transcription initiation, also establishes
5.1 Early Gene
Expression
5.1.1 Promoter Elements
Barbara J. Kelly et al.](https://image.slidesharecdn.com/84287-250511220407-b12cb833/85/Baculovirus-and-Insect-Cell-Expression-Protocols-3rd-Edition-David-W-Murhammer-Ed-48-320.jpg)

























![STATUTE.
9 Hen. 5.
[Cited by Sir Wm. Browne in his Vindication of the College of
Physicians. Quarto, London 1753.]
Ex Bundello petitionum de ano
. 9o
. H. 5. in Parliamento.
Hey and most mighty prince noble and worthy lords spirituelx and
temporelx and worshipfull comunes, for so moche as a man hath
thre things to governe, that is to say soule, body, and worldly goods,
the which ought and shulde ben principaly reweled by thre sciences,
that ben divinitie, fisyk, and lawe, the soule by divinitie, the body by
fisyk, worldly goods by lawe, and those conynges should be used
and practised principally by the most connyng men in the same
sciences, and most approved in cases necessaries to encrese of
virtue, long life, and gouds of fortune, to the worship of God and
comyn profit.
But worthi soveraines hit is known to your hey discretion, meny
uncunning and unaproved in the aforesaide science practiseth, and
specialy in fysyk, so that in this realme is every man be he never so
lewed taking upon him practice y suffered to use it to grete harm
and slaughtre of many men, where if no man practiced therein but al
only connynge men and approved sufficiently y learned in art,
filosofye, and fysyk, as it is kept in other londes and roialmes ther
shuld many man that dyeth for defaute of helpe lyve, and no man
perish by unconning.](https://image.slidesharecdn.com/84287-250511220407-b12cb833/85/Baculovirus-and-Insect-Cell-Expression-Protocols-3rd-Edition-David-W-Murhammer-Ed-74-320.jpg)



















































