Download to read offline



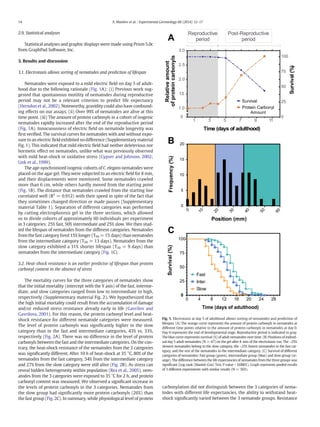
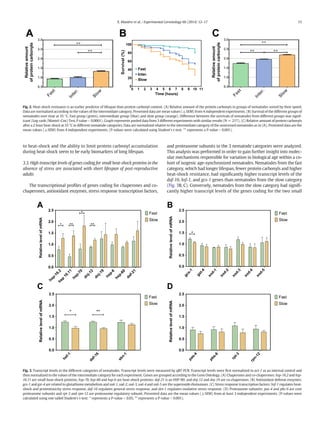



This study used an electrotaxis assay to separate Caenorhabditis elegans nematodes into groups based on their crawling speed in response to an electric field. Nematodes that crawled more slowly had shorter lifespans, higher levels of protein damage, and lower heat shock resistance than faster nematodes. Gene expression analysis found that slow nematodes had higher transcript levels of heat shock genes, which correlated with their poorer stress response and shorter lifespan. The results suggest that accumulation of early-life damage leads to faster age-related deterioration and a shorter lifespan.












































![[Name Sponged] Consulting Project (2)](https://cdn.slidesharecdn.com/ss_thumbnails/d43b862b-7ff4-4ce2-9f91-c72b4f73476a-151017140420-lva1-app6891-thumbnail.jpg?width=640&height=640&fit=bounds)





















