Downloaded 444 times






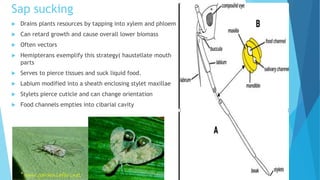












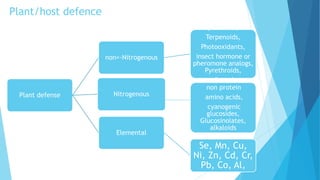



























This document provides an outline and overview of plant-insect interactions. It discusses different types of interactions like herbivory, mutualism, and parasitism. It describes how insects feed on plants through chewing, mining, boring, and sap sucking. It also discusses how plants defend against insects through production of secondary compounds, physical defenses, and induced responses. The document outlines how plants and insects have co-evolved over time through an evolutionary arms race of offense and defense traits.






















































































