Transcription and synthesis of different RNAs
Processing of RNA transcript
Catalytic RNA
RNA splicing and Spliceosome
Transport of RNA through nuclear pore
Translation and polypeptide synthesis
Posttranslational modification
Protein trafficking and degradation
Antibiotics and inhibition of protein synthesis.
Stunning ➥8448380779▻ Call Girls In Panchshil Enclave Delhi NCR
Protein Synthesis in Prokaryotes and Eukaroytes
1. Protein Synthesis in Pks & Eks
The simple Story that started complex LIFE!!
Prepared by
BIR BAHADUR THAPA
CDB, TU
NEPAL
Guided by
Dr. GIRI PD. JOSHI
CDB, TU
NEPAL
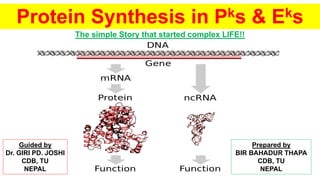
2. About picture, which did not tell itself is: -
A non-coding RNA (ncRNA) is an RNA molecule that is not translated into a
protein. The DNA sequence from which a functional non-coding RNA is transcribed
is often called an RNA gene. Abundant and functionally important types of non-
coding RNAs include transfer RNAs (tRNAs) and ribosomal RNAs (rRNAs), as well
as small RNAs such as microRNAs, siRNAs, piRNAs, snoRNAs, snRNAs, exRNAs,
scaRNAs and the long ncRNAs such as Xist and HOTAIR.
Non-coding RNAs contribute to diseases including cancers, autism, and
Alzheimer's.
Many of the newly identified ncRNAs have not been validated for their function. It is
also likely that many ncRNAs are non functional (sometimes referred to as Junk
RNA), and are the product of false transcription.
3. About sub-heading “The simple Story that started LIFE!!”
• The origins of life? The central dogma is so central to all living things, but one
wonders how it may have evolved.
• Life requires both storage and replication of genetic information, and the ability to
catalyze specific reactions.
• RNA has both of these abilities.
• RNA thought to be the original molecule of life, carrying both genetic info and
performing chemical reactions (ribozymes).
• Life then shifted to a DNA platform for the storage of the genetic information because
of its increased chemical stability and double-stranded format that enables
proofreading
• Life then shifted to a protein platform for chemical processes ->broader chemical
functionality
4. Contents under
Protein synthesis in Prokaryotes and Eukaryotes:
• Transcription and synthesis of different RNAs
• Processing of RNA transcript
• Catalytic RNA
• RNA splicing and Spliceosome
• Transport of RNA through nuclear pore
• Translation and polypeptide synthesis
• Posttranslational modification
• Protein trafficking and degradation
• Antibiotics and inhibition of protein synthesis.
6. Transcription and synthesis of diff. RNAs
• What is transcription?
RNA synthesis (=Transcription) is the process of copying information in DNA
sequences into RNA sequences.
• How is transcription different from replication of DNA?
DNA replication serves to copy all the genetic material of the cell and occurs before a
cell divides.
Transcription copies short stretches of the coding regions of DNA to make RNA.
Different genes may be copied into RNA at different times in the cell's life cycle.
• What enzyme carries out transcription?
This process is catalyzed by the enzyme RNA Polymerase. "RNA polymerase" is a
general term for an enzyme that makes RNA. There are many different RNA
polymerases. The basic transcription process is more or less similar in prokaryotes and
eukaryotes, though the regulation of transcription is much more elaborate in
7. Transcription and synthesis of diff. RNAs
• How does RNA Polymerase carry out its function?
To carry out RNA synthesis, all RNA Polymerases, prokaryotic and eukaryotic. must do the following:
1. Search the DNA template for promoters (sites on the DNA where the polymerase binds to start
transcription.)
2. Interact with other proteins that regulate transcription.
3. Unwind a short stretch of the DNA to expose single stranded DNA to copy into RNA
4. Select the correct RNA nucleotides, based on the DNA sequence, and assemble the RNA chain.
5 Recognize termination signals and stop synthesizing RNA when a termination signal is detected.
• How is RNA Polymerase like DNA Polymerase?
Like DNA polymerase, RNA Polymerase synthesizes new strands only in the 5' to 3' direction.
• How is RNA Polymerase different from DNA Polymerase?
RNA Polymerase doesn't require a primer to start making RNA.
RNA Polymerase uses ribonucleotides, not deoxyribonucleotides.
8. Different RNAs
• The major RNAs can be assigned to three major classes:
(1) The cytoplasmic messenger RNAs (mRNAs) and their nuclear precursors (pre-
mRNAs) carry the information that is used to specify the sequence, and therefore
ultimately the structure, of all proteins in the cell.
(2) Other RNAs do not encode protein but function directly, playing major roles in
various metabolic pathways, including protein synthesis.
-These include the ribosomal RNAs (rRNAs) and transfer RNAs (tRNAs), which are
the key components of the protein synthesis machinery;
-the small nuclear RNAs (snRNAs), which form the core of the pre-mRNA splicing
system; &
-the small nucleolar RNAs (snoRNAs), which are important factors in ribosome
biogenesis. These RNAs are generally much longer-lived than mRNAs and therefore
often are referred to as stable or noncoding RNAs (ncRNAs).
9. Transcription and synthesis of diff. RNAs
• (3) The third and most recently identified class of RNA comprises several structurally related groups of
very small (21 to 25 nucleotides) RNA species that play important roles in regulating gene expression.
Base pairing between endogenous micro-RNAs (miRNAs) and target mRNAs in the cytoplasm
represses their translation into protein. The packaging of DNA into a nontranscribed form termed
heterochromatin is promoted by a class of nuclear, small centromeric RNAs (ncRNAs). The
introduction of small double-stranded RNAs into many cell types and organisms results in cleavage of
the target mRNA and consequent silencing of gene expression. This phenomenon is described as
RNA interference (RNAi), and the RNAs are referred to as small interfering RNAs (siRNAs). In
addition, a heterogeneous set of longer ncRNAs (lncRNAs) have been implicated in a variety of
nuclear events.
• How does an RNA polymerase know where to start copying DNA to make a transcript?
Signals in DNA indicate to RNA polymerase where it should start and end transcription. These signals
are special sequences in DNA that are recognized by the RNA polymerase or by proteins that help
RNA polymerase determine where it should bind the DNA to start transcription. A DNA sequence at
which the RNA polymerase binds to start transcription is called a promoter.
10. Transcription and synthesis of diff. RNAs
What does a promoter look like
in prokaryotes?
A typical prokaryotic promoter has
three recognizable elements:
1. The transcription start site
(this the base in the DNA
across from which the first
RNA nucleotide is paired).
Sequences that are before the
start site are said be
"upstream" sequences.
2. The -10 sequence: this is a 6
bp region centered about 10
bp upstream of the start site.
The consensus sequence at
this position is TATAAT (i.e.,
this is the sequence found at
this position in the majority of
promoters studied)
3. The -35 sequence: this is a 6 bp sequence at about 35 basepairs upstream
from the start of transcription. The consensus sequence at this position is
TTGACA.
The sequences at -10 and -35 are recognized and bound by the RNA
11. Transcription and synthesis of diff. RNAs
• How does RNA polymerase
bind and carry out transcription
in prokaryotes?
• Prokaryotic RNA polymerases
have 2 components, a core
enzyme and a sigma factor
(a.ka. sigma subunit).
• The sigma factor is necessary for
the RNA polymerase to bind
tightly to the promoter and initiate
transcription.
• Once transcription starts, the
sigma factor falls off, and the core
enzyme continues copying the
DNA into RNA till it reaches a
terminator. A terminator is a
sequence of DNA that signals
RNA polymerase to stop
transcribing.
12. Transcription and synthesis of diff. RNAs
• In what ways does transcription differ in prokaryotes and eukaryotes?
1. In eukaryotes, the DNA template exists as chromatin, not as free DNA. The packaging of the DNA
must therefore be "opened up" to allow access for transcription. We have already considered how
chromatin remodeling complexes and histone modifications can make DNA regions accessible for
transcription.
2. Eukaryotes have three RNA polymerases, not one as in bacterial cells.
3. All three eukaryotic RNA polymerases need additional proteins to help them get transcription
started. In prokaryotes, RNA polymerase by itself can initiate transcription.
4. In addition to promoters, eukaryotic genes often have extra regulatory sequences many kilobases
away from the transcription start site. In bacteria, regulatory sequences are generally adjacent to the
gene that they control.
5. In eukaryotic cells, transcription is separated in space and time from translation:
-Transcription happens in the nucleus, and the RNAs produced are processed further before they are
sent into the cytoplasm.
-Protein synthesis (translation) happens in the cytoplasm.
In prokaryotic cells, RNAs can be translated as they are coming off the DNA template, and because
there is no nucleus, transcription and protein synthesis occur in a single cellular compartment.
13. Eukaryotic Protein Synthesis vs. Prokaryotic Protein Synthesis
Eukaryotic Protein Synthesis
• mRNA molecules are monocistronic, containing the coding sequence only f
or one polypeptide.
• protein synthesis occurs in the cytoplasm.
• most of the gene have introns or non-
coding sequences along with exons or coding sequences. The exons are join
ed together and introns are removed during mRNA processing.
• The primary mRNA transcript undergoes processing and splicing to change
into a functional mRNA.
• mRNA molecules are modified by the addition of a 5’G cap formed of methyla
ted guanosine triphosphate.
• A poly A tail formed of about 200 adenine nucleotides is added at the 3’end of
mRNA.
• 5’cap initiates translation by binding the mRNA to small ribosomal subunit us
ually at the first codon AUG.
• The first amino acid methionine entering the ribosome is not formylated.
• The pre imitation complex formation is initiated by nine initiated factors.
• No. of initiating factors is much more than prokaryotes. About ten IFs have be
en identified in reticulocytes an RBC. These are eIF1, eIF2, eIF3, eIF4 , eIF5
, eIF6 ,eIF4B, eIF4C,eIF4D, eIF4F
• small subunit of ribosome (40 S) gets dissociated with the initiator amino acyl
tRNA (MettRNA Met) without the help of mRNA. The complex joins mRNA la
ter on.
Prokaryotic Protein Synthesis
• mRNA molecules are
polycistronic containing the coding sequence of several genes of a part
icular metabolic pathway.
• protein synthesis begins
even before the transcription of mRNA molecule is completed. This is cal
led coupled transcription - translation.
• do not have introns (Except
Archaebacteria). Therefore mRNA processing is not required.
• splicing of mRNA transcript does not occur.
• No such cap is formed at 5’end of bacterial mRNA.
• No poly A tail is added to bacterial mRNA.
• translation begins at an AUG codon preceded by a special nucleotide se
quence.
• The first amino acid methionine is formylated into N formyl methionine.
• Only two initiating factors are involved.
• Three initiating factors found in prokaryotes. PIF-1 , PIF-2 , PIF-3
• 30 S subunit first complexes with mRNA (30S-
mRNA) when then joins with f Met tRNA f-
14. Transcription and synthesis of diff. RNAs
• What do the three RNA Polymerases do?
RNA polymerase I- transcribes ribosomal RNA genes
RNA polymerase II- transcribes protein coding genes (that is, it makes mRNA)
RNA polymerase III- transcribes transfer RNA genes.
We will focus on RNA Polymerase II (sometimes referred to as Pol II) which transcribes messenger
RNAs. As always, Pol II must find and bind a promoter to initiate transcription.
• What does a eukaryotic promoter look like?
Eukaryotic promoters have some recognizable features:
1. The start site for transcription.
2. The TATA-box: This is a sequence about 25 basepairs upstream of the start of transcription.
3. Variable numbers of upstream elements: these are short DNA sequences that are within 100 bp
upstream of the start of transcription.
15. Transcription and synthesis of diff. RNAs
• What are the additional proteins needed to start
transcription?
General transcription factors are proteins that help eukaryotic
RNA polymerases find transcription start sites and initiate RNA
synthesis.
For RNA polymerase II these transcription factors are named TFIIA,
TFIIB and so on (TF= transcription factor, II=RNA polymerase II, and
the letters distinguish individual transcription factors).
The complex composed of RNA polymerase and the general
transcription factors bound at the TATA box is called the basal
transcription complex or transcription initiation complex. It is
the minimum requirement for any gene to be transcribed.
16. Processing of RNA transcript
• The newly made RNA, also known as the primary transcript (the product of transcription is known as
a transcript) is further processed before it is functional. Both prokaryotes and eukaryotes process their
r & t-RNAs.
• The major difference in RNA processing, however, between prokaryotes and eukaryotes, is in the
processing of mRNAs.
• in bacterial cells, the mRNA is translated directly as it comes off the DNA template. In eukaryotic cells,
RNA synthesis, which occurs in the nucleus, is separated from the protein synthesis machinery, which
is in the cytoplasm. In addition, eukaryotic genes have introns, noncoding regions that interrupt the
gene. The mRNA copied from genes containing introns will also therefore have noncoding regions that
interrupt the information in the gene. These noncoding regions must be removed before the mRNA is
sent out of the nucleus to be used for protein synthesis. The process of removing the introns and
rejoining the coding sections or exons, of the mRNA, is called splicing.
• Once the mRNA has been capped, spliced and had a polyA tail added, it is sent from the nucleus into
the cytoplasm for translation. The lifespan of RNAs varies greatly. Some RNAs are longlived, others
are rapidly degraded. RNA degradation is another step at which gene expression can be regulated
after the initial transcriptional control step.
17. Processing of RNA transcript
• What are the processing steps
for messenger RNAs?
Messenger RNAs are processed in
eukaryotic cells, not in bacterial cells.
The three main processing steps are
1. Capping at the 5' end
2. Addition of a polyA tail at the 3'
end and
3. Splicing to remove introns
18. Processing of RNA transcript
• What is the initial transcript of mRNA
called?
The initial product of transcription of an mRNA
is called the pre-mRNA or primary transcript.
After it has been processed and is ready to be
exported from the nucleus, it is called the
mature mRNA or processed mRNA.
• What happens in the capping step?
In the capping step of mRNA processing, a 7-
methyl guanosine is added at the 5' end of
the mRNA.
19. Processing of RNA transcript
• What is the function of capping?
The cap protects the 5' end of the mRNA from degradation by nucleases and also helps to position
the mRNA correctly on the ribosomes during protein synthesis.
• What happens at the 3' end of the mRNA?
The 3' end of a eukaryotic mRNA is first trimmed, then an enzyme called PolyA Polymerase adds a
"tail" of about 200 As to the 3' end.
• What is the function of the tail?
Evidence indicates that the polyA tail plays a role in efficient translation of the mRNA, as well as in the
stability of the mRNA. The cap and the polyA tail on an mRNA are also indications that the mRNA is
complete (i.e., not defective).
• How are introns removed from the pre-mRNA?
Introns are removed from the pre-mRNA by the activity of a complex called the spliceosome. The
spliceosome is made up of proteins and small RNAs that are associated to form protein-RNA
enzymes called small nuclear ribonucleoproteins or snRNPs (pronounced SNURPS).
20. Processing of RNA transcript
• What signals indicate where an intron starts and ends?
The base sequence at the start (5' end) of an intron is GU while the sequence at
the 3' end is AG. There is also a third important sequence within the intron, called
a branch point, that is important for splicing.
21. Processing of RNA transcript
• The newly made RNA, also known as the primary transcript (the product of transcription is known as a
transcript) is further processed before it is functional. Both prokaryotes and eukaryotes process their r &
t-RNAs.
• The major difference in RNA processing, however, between prokaryotes and eukaryotes, is in the
processing of mRNAs.
• in bacterial cells, the mRNA is translated directly as it comes off the DNA template. In eukaryotic cells,
RNA synthesis, which occurs in the nucleus, is separated from the protein synthesis machinery, which is
in the cytoplasm. In addition, eukaryotic genes have introns, noncoding regions that interrupt the gene.
The mRNA copied from genes containing introns will also therefore have noncoding regions that
interrupt the information in the gene. These noncoding regions must be removed before the mRNA is
sent out of the nucleus to be used for protein synthesis. The process of removing the introns and
rejoining the coding sections or exons, of the mRNA, is called splicing.
• Once the mRNA has been capped, spliced and had a polyA tail added, it is sent from the nucleus into
the cytoplasm for translation. The lifespan of RNAs varies greatly. Some RNAs are longlived, others are
rapidly degraded. RNA degradation is another step at which gene expression can be regulated after the
22. Catalytic RNA
• cRNA are RNA molecules that have enzyme activity. The classic
example is the hammerhead ribozyme. Catalytic RNAs are involved in a
number of biological processes, including RNA processing and protein
synthesis. Discovery of catalytic RNA contributed to the hypothesis of an
'RNA world', describing the origin of life as starting from RNA.
• Some RNA molecules can function like enzymes and exert a catalytic
action on themselves or on other molecules, known as Catalytic RNA,
presenting a new target for drugs and can be used for inactivating
unwanted RNA or DNA molecules by a specific cleavage reaction.
23. Catalytic RNA
• Discovery- 1980s
Until recently, protein enzymes were thought to be the only biologically active
catalysts. It was believed that DNA stored the genetic information and that RNA played
the role of an intermediate courier between the genetic messages contained in the
DNA and the ribosomes, where proteins are synthesized. Other RNAs, like transfer
RNAs (tRNAs) and ribosomal RNAs (rRNAs), were considered as helper molecules to
assist the function of proteins.
• Today, RNA molecules are the only molecules known both to store genetic
information (as in the RNA viruses and viroids or during transport in the form of
mRNA) and to exert biological catalysis.
• nondispensable biological activity- In the tRNA-processing enzyme ribonuclease
P (RNase P), the RNA moiety contains the catalytic activity.
• Self-splicing introns often occur within ribosomal genes, as in the rRNA
24. Catalytic RNA
• Very recently, it has been established that protein synthesis on the ribosome is
catalysed by the 23S ribosomal RNA compound and, thus, that the ribosome is
actually a ribozyme (Nissen et al., 2000).
• Ribozymes need to acquire three-dimensional architectures to promote specific
interactions with cofactors, especially divalent metal ions, and other functional
domains for processing RNA substrates.
• Ribozymes are generally built up of several structural subdomains made of helical
segments connected by tertiary contacts.
-Functional regions are usually located in single-stranded regions, such as internal
loops or bulges.
-Specific tertiary contacts occur between hairpin and internal loops, especially those
positioned on the outside of the molecule.
-The subdomains have various functions and are responsible for substrate recognition,
specific sequence alignment and catalytic activity, leading to a modular and
hierarchically organized architecture.
25. Catalytic RNA
• Some ribozymes, like the hammerhead ribozyme, the hairpin ribozyme and the
RNAase P RNA, are under extensive clinical research for their ability to cleave other
specifically chosen substrate RNAs. The therapeutic applications range from
cleavage of viral RNAs, like the acquired immune deficiency syndrome (AIDS)-
causing human immunodeficiency virus (HIV) RNA, to silencing of carcinogenic or
mutated cellular RNAs, or the control of gene expression in vivo.
Secondary structure of a minimal
hairpin ribozyme with substrate RNA
bound. Circles represent individual
nucleotides and lines indicate
canonical (Watson-Crick) base pairs
Predicted secondary
structure and
sequence
conservation of the
HH9 ribozyme found
conserved from lizard
to human genomes.
26. Catalytic RNA
Sequence and Structure
RNA molecules are built up of RNA helices interlinked by internal loops and multiple
junctions. Double-stranded helical regions are formed through standard Watson–Crick
base pairs.
One groove of the standard type A-form double-stranded RNA helix is deep and
narrow, while the other one is shallow and wide.
Double-stranded regions mainly act as a framework or rigid spacer to organize and to
orientate other structural and functional recognition elements.
Helices can be connected by three-, four-or multi-way junctions, which form a
sequence-dependent joint, which is either flexible or fixed, depending, among other
factors, on the type and concentration of metal ions.
Single-stranded regions comprise a whole range of structural elements, including base
bulges, internal loops or bubbles and terminal or hairpin loops. These can simply serve
as flexible linkers between helical domains, or they interact with other bases, usually
forming non-Watson–Crick pairs to widen or tighten the grooves or to induce kinks and
bends in the RNA backbone.
27. Catalytic RNA
• Catalytic Mechanisms
• RNAase P RNA- an endoribonuclease responsible for the maturation of the 5′ termini
of the majority of all known tRNAs in all cell types studied to date, involves in a site-
specific hydrolysis. The enzyme is conserved in all three kingdoms—bacteria,
archaea, and eukarya—with mitochondria and chloroplasts having activities separate
from the nucleus in eukaryotes.
• Eukaryotes also contain a second RNA-protein enzyme, called RNase MRP, which is
closely related to RNase P. The RNA components share common structural features,
and the complexes share eight common proteins. RNase MRP cleaves the pre-rRNA
between the small and large subunit rRNAs.
• The biological reactions catalysed by ribozymes mainly involve phosphodiester
cleavage or transfer (i.e. relegation to another nucleotide). The 2’ ribose hydroxyl
group, characteristic of RNA, is always directly or indirectly involved in catalysis.
• Ribozymes can be considered as metalloenzymes. Indeed, the folding and/or
catalytic activities of ribozymes often depend on the presence of and interaction with
divalent metal ions (Pyle, 1993).
28. Catalytic RNA
Classes of Catalytic RNAs
1. tRNA processing by ribonuclease P RNA- a true recycling catalyst is the ubiquitous endonuclease enzyme processing
precursor tRNA 5’ ends. Functional groups involved in catalysis are the 2’ ribose hydroxyl group of the conserved pyrimidine
(usually a U) at the position preceding the cleavage and a highly conserved purine at the position of the cleavage on the pre-
tRNA.
2. Intron Splicing- All three major types of cellular RNAs, tRNA, rRNA and mRNA, are known to contain intervening sequences
(IVS) or introns, which have to be precisely excised to produce functional molecules or the right reading frame for protein
synthesis. This process, known as RNA splicing, requires the recognition of the 5’ and 3’ exonic sequences, strand cleavage
and ligation. Although most eukaryotic introns are spliced by a complex machinery, the spliceosome, some introns in
bacteriophages or organelles self-splice.
3. Small Catalytic RNAs- simplest, each undergoing self cleavage and has a well-defined tertiary structure in the presence of
divalent cations.
• The hammerhead ribozyme- smallest known ribozyme. All conserved bases are in the single-stranded regions linking the
helical stems and terminal loops add stability and ensure correct folding. The three dimensional structure obtained by X-ray
diffraction studies (Pley et al., 1994) shows that most of the nucleotides in the single-stranded regions base-pair to form a Y-
shaped structure.
• The hairpin ribozyme- second smallest catalytic RNA molecule with a functional length of 50 nucleotides. It catalyses not only
a cleavage reaction but also its reverse reaction (ligation).
29. Catalytic RNA
• Ribozymes: Catalytic RNAs that cut things, make things, and do odd and
useful jobs- Walter aand Engelke, 2002
• Catalytic RNAs, or ribozymes, are a fossil record of the ancient molecular evolution
of life on Earth and still provide the essential core of macromolecule synthesis in all
life forms today. Are they also an avenue to the development of new catalysts to
recreate evolution, or to use as therapeutics and molecule sensors? (Walter &
Engelke, 2002)
• The original discovery of ribozymes by Cech and Altman (Nobel laureate- 1990)
was twofold – RNA segments that cut themselves out of larger RNAs (self-splicing
introns) and a protein-assisted RNA enzyme (ribonuclease P) that cuts the leader
sequences off all transfer RNAs throughout the three organismal domains.
• Universal use of RNA in synthesis of macromolecules
• The use of RNA in protein synthesis has long been part of the central dogma. Not
only is information carried in messenger RNA (mRNA) triplet codons, but transfer
RNA (tRNA) serves as the adapter to interpret codons into amino acids; and the
enormously complex ribosome, containing both ribosomal RNA (rRNA) and protein
30. Catalytic RNA
• Ribozyme Gene Therapy- Lewin & Hauswirth, 2001
• RNA enzymes – ribozymes – are being developed as treatments for a variety of diseases ranging
from inborn metabolic disorders to viral infections and acquired diseases such as cancer. Ribozymes
can be used both to down regulate and to repair pathogenic genes. In some instances, short-term
exogenous delivery of stabilized RNA is desirable, but many treatments will require viral-mediated
delivery to provide long-term expression of the therapeutic catalyst. Current gene therapy applications
employ variations on naturally occurring ribozymes, but in vitro selection has provided new RNA and
DNA catalysts, and research on trans-splicing and RNase P has suggested ways to harness the
endogenous ribozymes of the cell for therapeutic purposes.
• There are two basic modes for therapy that targets the genetic basis of disease: replace or resect. For
diseases caused by recessive mutations, gene therapists try to complement the defective gene. For
dominant disease mutations, however, introducing a normal gene will not work. Many of these
diseases are associated with hyperactivation (in the case of oncogenes), or aggregation of a mutant
protein (in the case of neurodegenerative diseases), with a normal copy of the same gene being
present on the partner chromosome. In these cases, expression of the defective gene must be
silenced or at least limited. For such autosomal dominant genetic diseases, ribozymes are a
particularly appealing tool: they can be used to reduce expression of a pathogenic gene by digesting
32. Catalytic RNA
• Ribosome is a Ribozyme- Cech, 2000
• The AA we obtained by digestion of steak, salmon or lettuce salad are loaded onto
tRNAs and rebuilt into proteins in the ribosome, the cell’s macromolecular protein
synthesis factory…. Its Key component are so highly conserved among all of the
Earth’s species that a similar entity must have fueled protein synthesis in the
LUCA i.e., Last Universal Common Ancestor i.e. ancestor of all extant life.
• Ribosome task- simple one i.e. joining multiple AA through peptide linkage but it
also performs the remarkable task of choosing the AA to be added to the growing
polypeptide chain by reading successive mRNA.
34. RNA splicing and Spliceosome
• Spliceosomes are dynamic macromolecular multimegadalton organelles, a endogenous
RiboNucleoProteinComplexes (RNPC) composed of small nuclear RNA (snRNA) & approximately 100 other
associated proteins with a ribozyme at its core, that remove intervening noncoding regions- introns in protein-
encoding genes of a precursor messenger RNA (pre-mRNA).
• The assembly depends on both complementary base pairing between the small nuclear RNAs and the intron
and exon substrates, and on extensive protein–RNA and protein–protein interactions. These interactions are
fundamental for proper spliceosomal function.
• At the core of the spliceosome are five small nuclear RNAs (snRNAs) termed U1, U2, U4, U5, and U6. U1-U5
snRNAs are produced by RNA polymerase II transcription and are 5′- capped with 7-methylguanosine, while U6
is transcribed by RNA Pol III and has a different cap structure.
• Each of the snRNAs is associated with a set of 8 Sm proteins, B/B′, D3, D2, D1, E, F, and G.
• The Sm proteins bind to each other and to a highly conserved sequence on U1, U2, U4, and U5 snRNAs.
• The association of Sm proteins and snRNAs occurs in the cytoplasm in a controlled and precise order, and the
5′cap structure of snRNAs must be hypermethylated before the small nuclear ribonucleoprotein (snRNP)
complex is imported back into the nucleus.
• U6 snRNA diverges from this assembly pathway and associates to Sm-like proteins LSm2, LSm3, LSm4, LSm5,
LSm6, LSm7, and LSm8 in the nucleus.
35. RNA splicing and Spliceosome
• How does splicing actually take place?
There are two main steps in splicing-
1st step, the pre-mRNA is cut at the 5' splice site
(the junction of the 5' exon and the intron).
The 5' end of the intron then is joined to the
branch point within the intron. This generates
the lariat-shaped molecule characteristic of the
splicing process.
2nd step, the 3' splice site is cut, and the two
exons are joined together, and the intron is
released.
• https://www.youtube.com/watch?v=FVuAwBGw_pQ
37. RNA splicing and Spliceosome
• If there are multiple exons in an mRNA can they be spliced in
different combinations?
Yes, different combinations of exons can be spliced together to
generate different mature mRNAs (recall that this is called alternative
splicing).
38. RNA splicing and Spliceosome
• What is the advantage of alternative splicing?
Alternative splicing allows the production of many different proteins using relatively
few genes, since a single RNA with many exons can, by mixing and matching its
exons during splicing, create many different protein coding messages. Because of
alternative splicing, each gene in our DNA gives rise, on average, to three different
proteins.
• What happens to RNAs after they have fulfilled their functions? All
RNAs are eventually degraded in the cell. Ribosomal RNAs and transfer RNAs,
which are in constant use in the cell for protein synthesis, are relatively long-lived.
The stability of messenger RNAs varies widely. Some are degraded rapidly, while
others may have a longer half-life. The level of a particular mRNA is determined by
both the rate of its synthesis and the rate of its breakdown by the cell. This provides
the cell with an additional point of control for gene expression.
39. Nuclear Pore Complexes
Nuclear Pore Complexes: Small-sized nuclear
pores keep most large molecules from entering
the nucleus. But some large protein molecules
are required in the nucleus and enter it though a
larger complex protein structure called the
nuclear pore complex (NPC). This complex
consists of at least 456 protein molecules. It
surrounds each pore and allows larger molecules
to exit or enter by expanding the pore openings to
a larger size. NPC's cross the double-walled
nuclear membrane, allowing the transport of
water-soluble molecules through the membrane.
Nuclear pore- Side view.
1. Nuclear envelope.
2. Outer ring.
3. Spokes.
4. Basket.
5. Filaments.
40. Transport of RNA through nuclear pore
• Since the DNA is transcribed into mRNA in the nucleus, and protein synthesis takes
place in the cytoplasm, the mRNA has to exit the nucleus to the cytoplasm. The
environment in the nucleus differs in many ways from that of the cytoplasm. To
separate these two environments from each other the nucleus is enclosed by a
double membrane, and the only connection to the surrounding cytoplasm is
through channels called the nuclear pore complex (NPC).
• When it is time for the mRNA to leave the nucleus, the mRNA is believed to be
"tagged" by proteins which serve as export signals, directing the mRNA to the
nuclear pore complex that the mRNA is to leave. The mRNA and it's bound export
proteins then attaches to export receptors and the whole complex (RNA, export
signal proteins and export receptors) is translocated through the nuclear pore
complex. The mRNA is released into the cytoplasm and is immediately ready for
the next step: translation»
41. Transport of RNA through nuclear pore
a. The mRNA molecule is transported to the nuclear pore. Before the translocation
through the nuclear pore begins, some proteins, for example the splicing
components, disassociate from the mRNA.
b. Export proteins bind to the mRNA and as a first step in the translocation, the
mRNA docks with the cage-like structure of the nuclear pore complex.
c. The mRNA is translocated through the nuclear pore with the 5' end, the CAP
structure, in the lead. The translocation through the nuclear pore is an energy
requiring process, but the mechanism for the transport is not known.
d. When the mRNA arrives at the cytoplasmic side of the nuclear pore, even if parts
of the RNA are still within the pore or on the nuclear side, many proteins
disassociate from the mRNA. Among those are the export proteins which return to
the nucleus. The mRNA is immediately ready for the next step: translation
42. Transport of RNA through nuclear pore
a. The mRNA molecule
is transported to the
nuclear pore. Before the
translocation through the
nuclear pore begins,
some proteins, for
example the splicing
components,
disassociate from the
mRNA.
b. Export proteins bind
to the mRNA and as a
first step in the
translocation, the mRNA
docks with the cage-like
structure of the nuclear
pore complex.
c. The mRNA is translocated
through the nuclear pore with
the 5' end, the CAP
structure, in the lead. The
translocation through the
nuclear pore is an energy
requiring process, but the
mechanism for the transport
is not known.
d. When the mRNA arrives
at the cytoplasmic side of the
nuclear pore, even if parts of
the RNA are still within the
pore or on the nuclear side,
many proteins disassociate
from the mRNA. Among
those are the export proteins
which return to the nucleus.
The mRNA is immediately
45. Translation and Polypeptide synthesis
Once mRNA has passed out of the nuclear pore, it determines the synthesis of a
polypeptide.
1. A ribosome attaches to the starting codon, at one end of an RNA molecule.
2. The tRNA molecule with a complementary anticodon sequence moves to the
ribosome, and pairs up with the sequence on the mRNA.
3. A tRNA molecule with a complimentary anticodon pairs up with the next codon
on the mRNA.
4. The ribosome moves along the mRNA, bringing together two tRNA molecules,
each pairing up with the corresponding two codons on the mRNA.
5. By means of an enzyme, ATP and and two amino acids on the tRNA are joined
by a peptide bond.
6. The ribosome moves onto the third codon, on the mRNA linking the amino
acids.
7. As this happens the first tRNA is released from its amino acid, from the amino
acid pool in the cell.
8. This process continues until a complete polypeptide chain is created.
46. Translation and Polypeptide synthesis
1. Initiation
• Translation initiation operates through a 30S initiation complex
(30S-IC), consisting of the 30S, mRNA, initiator fMet-tRNA, and
three initiation factors, IF1, IF2, and IF3.
• Subsequently, the 30S-IC associates with the 50S, which
releases the Initiation Factors (IFs) and leaves the initiator-
tRNA at the peptidyl-tRNA-binding Site (P site), base-paired to
the start Codon of The mRNA.
47. Translation and Polypeptide synthesis
2. Elongation
The elongation phase involves the movement of tRNAs in a cyclic fashion through the three
tRNA-binding sites (A→P→E) on the ribosome. The first step in the cycle involves the delivery of
the aminoacyl-tRNA (aa-tRNA) to the aa-tRNA-binding site (A site), which is facilitated by the
elongation factor EF-Tu•GTP.
Hydrolysis of GTP by EF-Tu leads to its dissociation from the ribosome, allowing aa-tRNA
accommodation. Peptide-bond formation then proceeds, transferring the entire polypeptide chain
from the P-tRNA to the aa-tRNA in the A site. The ribosome now has a peptidyl-tRNA at the A site
and an uncharged tRNA at the P site.
This ribosomal state is highly dynamic with the tRNAs oscillating between classical (A and P
sites) and hybrid states (A/P and P/E sites on 30S/50S). EF-G binds to the ribosome, which
stabilizes the tRNAs in hybrid states, hydrolyzes GTP to GDP, and catalyzes the translocation
reaction. Translocation shifts the peptidyl-tRNA from the A/P hybrid state to the P site and the
deacylated tRNA from the P/E to the exit site (E site). EF-G•GDP dissociates leaving the A site
48. Translation and Polypeptide synthesis
3. Termination/Recycling
• Arrival of an mRNA stop codon in the A site of the ribosome
signals the termination of protein synthesis.
• Release factor 1 (RF1) or RF2 binds to the ribosome and
hydrolyzes the peptidyl-tRNA bond, releasing the translated
polypeptide chain from the ribosome.
• RF1/2 is recycled from the ribosome by RF3 in a GTP-
dependent fashion.
• The ribosome is then split into subunits by the concerted action
of EF-G and ribosome recycling factor (RRF), thus recycling the
components for the next round of translation.
49.
50. Translation and Polypeptide synthesis
1. tRNAs attach to the correct
amino acid in the cytoplasm –
directed by enzymes
2. Small subunit of the ribosome
binds to the 5’ cap of the mRNA
51. Translation and Polypeptide synthesis
3. The first tRNA attaches to the
first three nucleotides of the
mRNA
The first three nucleotides of every
mRNA is AUG. This codes for the
amino acid Methoinine.
52. Translation and Polypeptide synthesis
4. The large subunit
slides into place so the
tRNA is in the middle
slot of the ribosome,
the P site.
53. Translation and Polypeptide synthesis
5. The next tRNA
with the
appropriate amino
acid enters the A
site matching to
the mRNA codon.
- Determine the
amino acid coming
in based on the
mRNA codon
sequence – use
mRNA codon chart.
54. Translation and Polypeptide synthesis
6. The amino acid on
the tRNA in the P
site attaches to the
amino acid on the
tRNA in the A site
and forms a peptide
bond.
55. Translation and Polypeptide synthesis
7. The ribosome shifts
down the mRNA causing
the tRNAs (which are still
attached to the mRNA) to
translocate (move) into
the adjacent site on the
ribosome.
- the tRNA in the A site
moves to the P site
- the tRNA in P site moves
to the E site and then
leaves
56. Translation and Polypeptide synthesis
8. This process (steps 5 – 7)
repeats over and over
building the amino acid
chain until a STOP codon
is reached.
- STOP codons = UGA,
UAG, UAA
- bring in a protein called a
Release Factor and this
causes the amino acid
chain (protein) to be freed
and the ribosome to detach
from the mRNA.
57. Translation and Polypeptide synthesis
• What is peptide: A Simple Introduction of Peptide Synthesis.
• Peptides and proteins are linear polymers of amino acids linked by
amide "peptide bonds" (Fig. 1). The peptide bond is formed by linking an
amino group to a caroboxyl group on another amino acid. Amino acids
are primary amines that contain an alpha carbon that is connected to an
amino (NH3) group, a carboxyl group (COOH), and a variable side
group (R). Carboxylic acid and carboxylate groups are normally not very
reactive. It requires activating the carboxylic acid for the formation of the
amide peptide bond.
60. Post-translational modification (PTM)
• Newly synthesized polypeptides in the membrane and lumen of the ER
undergo five principal modifications before they reach their final destinations:
1. Formation of disulfide bonds
2. Proper folding
3. Addition and processing of carbohydrates
4. Specific proteolytic cleavages
5. Assembly into multimeric proteins
• The most common are:-
-specific cleavage of precursor proteins;
-formation of disulfide bonds; or
-covalent addition or removal of low molecular weight groups thus leading to
modifications like Phosphorylation, Acetylation, N-linked glycosylation, Amidation,
Hydroxylation, Methylation, O-linked glycosylation, Ubiquitylation, Pyrrolidone
Carboxylic Acid, Sulfation
61. Post-translational modification (PTM)
Disulfide Bonds Are Formed and Rearranged in the ER Lumen
Correct Folding of Newly Made Proteins Is Facilitated by Several ER
Proteins
Assembly of Subunits into Multimeric Proteins Occurs in the ER
Only Properly Folded Proteins Are Transported from the Rough ER to
the Golgi Complex
Many Unassembled or Misfolded Proteins in the ER Are Transported to
the Cytosol and Degraded
ER-Resident Proteins Often Are Retrieved from the Cis-Golgi
62. Post-translational modification (PTM)
• PTM is a biochemical modification that occurs to one or more amino acids on a protein after
the protein has been translated by a ribosome, the covalent and generally enzymatic
modification of proteins following protein biosynthesis. Proteins are synthesized by
ribosomes translating mRNA into polypeptide chains, which may then undergo PTM to form
the mature protein product. PTMs are important components in cell signaling.
• Fundamental role- Regulating the folding of proteins, their targeting to specific subcellular
compartments, their interactions with ligands or other proteins, and their functional states,
such as catalytic activity in the case of enzymes or the signaling function of proteins
involved in signal transduction pathways.
• Study Method- PROTEOMICS i.e. systemic identification of proteins from cells and tissues
that involves separation of proteins and their isoforms by size or charge heterogeneity by 2-
dimensional gel electrophoresis, recovery of the individual spots from gel by mass
spectrometry.
• Post-translational modifications can occur on the amino acid side chains or at the protein's
C- or N- termini. They can extend the chemical repertoire of the 20 standard amino acids by
63. Post-translational modification (PTM)
Phosphorylation is a very common mechanism for regulating the activity of enzymes
and is the most common post-translational modification.
Many eukaryotic proteins also have carbohydrate molecules attached to them in a
process called glycosylation, which can promote protein folding and improve stability as
well as serving regulatory functions.
Attachment of lipid molecules, known as lipidation, often targets a protein or part of a
protein attached to the cell membrane. Varying degrees of posttranslational
modification affect the hydrophobicity by adding a glycophospholipid to the C-terminal
carboxyl group of the protein(s). The lipid allows the enzyme to be tethered to plasma
membranes.
Other forms of post-translational modification consist of cleaving peptide bonds, as in
processing a propeptide to a mature form or removing the initiator methionine residue.
The formation of disulfide bonds from cysteine residues may also be referred to as a
post-translational modification. For instance, the peptide hormone insulin is cut twice
after disulfide bonds are formed, and a propeptide is removed from the middle of the
64. Post-translational modification (PTM)
Glycosylation is a posttranslational modification whereby one or more sugar groups
are covalently linked to a target protein forming a glycoprotein. These sugars are often
essential for the proper structure and function of
the protein.
Some types of post-translational modification are consequences of oxidative stress.
Carbonylation is one example that targets the modified protein for degradation and
can result in the formation of protein aggregates. Specific amino acid modifications can
be used as biomarkers indicating oxidative damage.
66. Antibiotics and Inhibition of Protein Synthesis.
• A protein synthesis inhibitor is a substance that stops or slows the
growth or proliferation of cells by disrupting the processes that lead
directly to the generation of new proteins.
• any antibiotic, usually refers to any substances that act at the ribosome
level (either the ribosome itself or the translation factor), taking
advantages of the major differences between prokaryotic and
eukaryotic ribosome structures.
67. Antibiotics and Inhibition of Protein Synthesis.
1. Inhibitors of both Prokaryotic and Eukaryotic Protein Synthesis:
• Aurintricarboocylic acid inhibits formation of the initiation complex by preventing the
association of mRNA with the small ribosomal subunit.
• Edeine, a polypeptide isolated from Bacillus brevis, inhibits the binding of
aminoacyl-tRNA and N-formylmet-tRNAM
f
et (in prokaryotes) to the small subunit.
• Fusidic acid is a steroidal antibiotic; in prokaryotes, it inhibits the binding of
aminoacyl-tRNA to the ribosome, whereas in eukaryotes, it inhibits translocation by
reacting with elongation factor.
• Puromycin, mimics aminoacyl-tRNA and binds to the free A site of ribosomes
engaged in protein synthesis. Catalytic formation of a bond between the nascent
polypeptide and puromycin is followed by the release of the peptidyl-puromycin from
the ribosome, as no further elongation is possible.
68. Antibiotics and Inhibition of Protein Synthesis.
2. Inhibitors Specific for Prokaryotes:
• Chloramphenicol (Chloromycetin) binds to the large subunit of prokaryotic
ribosomes and interferes with the functioning of peptide synthetase, thereby inhibit-
ing chain elongation.
• Colicin E3 inhibits protein synthesis in prokaryotes by interfering in some manner
with the functioning of the small subunit.
• Erythromycin binds to ribosomes that are not engaged in protein synthesis,
preventing their potential participation, but does not bind to ribosomes containing
nascent chains (i.e., ribosomes that are part of a functioning polysome).
• Streptomycin binds to protein S12 of the small ribosome subunit, causing release of
N-formylmet-tRNAM
f
et from initiation complexes (thereby preventing initiation of
chain growth) and also causing misreading of the codons of mRNA by ribosomes
already involved in chain elongation.
69. Antibiotics and Inhibition of Protein Synthesis.
3. Inhibitors Specific for Eukaryotes:
• Anisomycin is an antibiotic produced by Streptomyces that inhibits peptide bond
formation when bound to the small ribosomal subunit.
• Cycloheximide binds to the large subunit, preventing the translocation of tRNA in
the A site to the P site.
• Diphtheria toxin (produced by a strain of Corynebacterium diphtherial) inhibits
protein synthesis through its action on EF-2 (translocase).
• Ricin acts on the large subunit, preventing formation of the 80 S initiation complex.
• Sparsomycin, produced by Streptomyces, inhibits the association of the amino acid
moiety of aminoacyl-tRNA from binding to the large subunit and, in so doing, blocks
peptide synthetase.
• THchodermin is the only chemical compound so far identified as a specific inhibitor
of the termination stage of polypeptide synthesis.
70. Antibiotics and Inhibition of Protein Synthesis.
4. Inhibitors of Organeiiar Protein Synthesis:
• Chloramphenicol, a strong inhibitor of prokaryote protein synthesis,
blocks synthesis in mitochondria and chloroplasts,
• whereas cycloheximide, which blocks eukaryote cytoplasmic ribosomal
protein synthesis, is without effect on mitochondrial and chloroplast
synthesis.
• These observations provided added credence for the notion that
prokaryotic cells, mitochondria, and chloroplasts have a common
evolutionary origin.
• It is now clear, however, that the picture is considerably more complex.
For example, streptomycin, which inhibits prokaryotic protein synthesis,
fails to inhibit mitochondrial protein synthesis in yeast cells.
• Erythromycin inhibits the synthesis of proteins in prokaryotes, yeast
mitochondria, and chloroplasts but fails to block protein synthesis in
71. Antibiotics and Inhibition of Protein Synthesis.
4. Inhibitors of Organeiiar Protein Synthesis:
• The nature of mitochondrial protein synthesis varies among
different groups of eukaryotes. Mitochondria from higher
eukaryotes are more resistant to inhibitors of prokaryotic
protein synthesis than are mitochondria from lower eukaryotes.
• In chloroplasts, protein synthesis is inhibited by the same
agents that block this process in prokaryotic cells.
72. Antibiotics and Inhibition of Protein Synthesis.
4. Inhibitors of Organeiiar Protein Synthesis:
• The differential sensitivity of eukaryotic cytoplasmic and
mitochondrial ribosomes to specific inhibitors provides a means for
examining the sources of certain mitochondrial proteins.
• The synthesis of a mitochondrial protein in the presence of
cycloheximide (a cytoplasmic inhibitor) indicates that the mitochondria
are the source of the protein, whereas synthesis of the protein in the
presence of chloramphenicol indicates that the mitochondrial protein is
produced in the cytoplasm and then moves to the mitochondria.
73. Antibiotics and Inhibition of Protein Synthesis.
• 1. PCN/pen derived from Penicilium notatum
• Penicillin is a group of antibiotics which include
-penicillin G (intravenous use),
-penicillin V (oral use),
-procaine penicillin etc..
• degrades the cell wall of bacteria which is made up of peptidoglycan without
interfering with the host cell wall
74. Antibiotics and Inhibition of Protein Synthesis.
• 2. Streptomycin
• Derived from actino-bacterium Streptomyces griseus (G+ve bacteria;
with high G and C content in DNA)
• Trisaccharide
• Medically important from aminoglycosides family
• At higher concentration binds with fMet-tRNA to ribosomes and inhibit
initiation
• At lower concentration it leads to the misreading of genetic code
75. Antibiotics and Inhibition of Protein Synthesis.
2. Tetracycline
• Broad spectrum four ring compound produced from Streptomyces
• Blocks A site on the ribosomes so that binding of aminoacyl-tRNA is
inhibited.
3. Chloramphenicol
• Inhibits the peptidyl transferase activity on 70s ribosomes
• Act at low concentration
• Doesn’t affect cytosolic protein synthesis
76. Antibiotics and Inhibition of Protein Synthesis.
4. Cycloheximide
• Inhibitor of protein synthesis in eukaryotes
• Toxic to human and used in in-vitro research application
• Exerts its effect by interfering with translocation step thus blocking
translational elongation
• used as fungicide
5. Erythromycin
• Binds to 50s subunit of prokaryotes and blocks translocation step
thereby freezing the peptidyl –tRNA in the A site
77. Antibiotics and Inhibition of Protein Synthesis.
Stages- Protein
Synthesis
Antibiotics Mechanism
Earlier Rifamycin stage-inhibits prokaryotic DNA transcription into mRNA by inhibiting
DNA-dependent RNA polymerase by binding its beta-subunit.
Initiation Linezolid probably by preventing the formation of the initiation complex.
Ribosome
assembly
Neomycin prevents ribosome assembly by binding to the prokaryotic 30S
ribosomal subunit.
Aminoacyl tRNA
entry
Tetracyclines ,Tigecycline block the A site on the ribosome, preventing the binding of aminoacyl
tRNAs.
Proofreading Aminoglycosides, interfere with the proofreading process, causing increased rate of error
in synthesis with premature termination.
Peptidyl transfer Chloramphenicol blocks the peptidyl transfer step of elongation on the 50S ribosomal
subunit in both bacteria and mitochondria.
Macrolides bind to the 50s ribosomal subunits, inhibiting peptidyl transfer.
Ribosomal
translocation
Macrolides,
clindamycin,,aminoglycosi
de
ribosomal translocation inhibition
78. References
• Walter, F., & Westhof, E. (2001). Catalytic RNA. eLS.
• Walter, N. G., & Engelke, D. R. (2002). Ribozymes: catalytic RNAs that cut
things, make things, and do odd and useful jobs. Biologist (London, England),
49(5), 199.
• Pley HW, Flaherty KM and McKay DB (1994) Three-dimensional structure of
a hammerhead ribozyme. Nature 372: 68–74.
• Pyle AM (1993) Ribozymes: a distinct class of metalloenzymes. Science 261:
709–714.
• ltman S. Nobel lecture. Enzymatic cleavage of RNA by RNA. Biosci Rep.
1990; 10:317–337. [PubMed: 1701103]
• Butcher SE. Structure and function of the small ribozymes. Curr Opin Struct
Biol. 2001; 11:315–320. [PubMed: 11406380]
• Cech TR. Nobel lecture. Self-splicing and enzymatic activity of an intervening
sequence RNA from
• Tetrahymena. Biosci Rep. 1990; 10:239–261. [PubMed: 1699616]
• Cech TR. The ribosome is a ribozyme. Science. 2000; 289:878–879.