Russian Call Girls Nashik Anjali 7001305949 Independent Escort Service Nashik
Fish_Gill.pptx
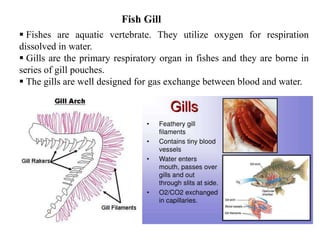
1. Fishes are aquatic vertebrate. They utilize oxygen for respiration
dissolved in water.
Gills are the primary respiratory organ in fishes and they are borne in
series of gill pouches.
The gills are well designed for gas exchange between blood and water.
Fish Gill
4. Gills in agnathous lamperys and hagfishes are in pouches
(Marsipobranch/pouch gills).
Each pouch has an internal opening from the pharynx and one that leads to or
towards the outside.
Other parts of the water route in lamprey and hagfishes include oral
vestibule, the mouth and the pharynx.
In the hagfishes a pituitary duct connects the exterior with the pharynx
dorsally.
In the adult lamprey, the pharynx becomes divided from the dorsal food
pathway anteriorly by a shelf of tissue, the velum.
The ventral respiratory tube thus formed leads to the gill pouches via the
internal branchial apertures.
Lampreys have seven pairs of external gill openings.
Some hagfishes have numerous gill pouches (5-14 in Eptatretus) on each side,
each with an internal and external aperture.
In other hagfishes (e.g. Myxine), the six pairs of gill pouches open on each side
into a common duct, which in turn opens to the outside near the level of sixth
pouch.
Functionally this arrangement parallels the opercular gill covers in Chimaeras
and the bony fishes.
5. Every hagfish has a tube (esophageocutaneous duct) aft of the gill pouches
on the left side only.
The gill in gnathostomes are carried on branchial arches.
In shark and relatives (Elasmobranch) the gill are in chambers resembling
those of the pouch gill of lampreys and hagfish.
The interbranchial septa that separate the chambers in Chondrichthyes which
are variously reduced in Osteichthyes where gills of all arches lie in a more or
less common chamber.
The septa are thus either complete (Chondrichthyes) or variously partial
(Osteichthyes) and each typically bears a hemibranch on each side.
Gill filaments in Chondrichthyes are ridge-like folds on the interbranchial
septa (Giving name Elasmobranch meaning Plate gill).
7. Fig. Generalized schematics of gills and associated pouches/arches in hagfishes
(A and E), lampreys (B and F), elasmobranchs (C and G), and teleosts (D and H). All
schematics are oriented anterior to posterior, with oral opening at top.
8. Gills in Lampreys and Hagfish
The seven paired gill sacs or branchial pouches
The gill sacs open toward the lumen of the alimentary canal.
Each is divided from the next by a thin diaphragm
9. Gills in Shark and Rays
Five, more rarely six or seven external gill slits exist ventrally in a
series on each side in rays and their relatives but laterally in sharks.
An anterior addition to this series of openings in both shark and rays
is the spiracle, corresponding to a vestigial gill slit.
Internal to the spiracle, gill lamellae may persist as a hyoidean
pseudobranch.
In shark, respiratory water typically enters through the mouth, in the
bottom dwelling rays the spiracle admits most of water that flows
subsequently over the gills.
Well developed septa with cartilaginous supports and individual gill-
arch.
Both primary and secondary gill filament are present in each
hemibranch. The primary end s of primary gill filaments are detached
from the septum.
10.
11.
12.
13.
14.
15. • Gills are the main
site of gas exchange
in almost all fishes.
• The gills consist of bony or
stiffened arches (cartilage) that
anchor pairs of gill filaments.
18. OMNIVOROUS
In the omnivorous fishes
like Puntius sarana,gill
rakers are short and
stumpy.
HERBIVOROUS:
Like Labeo rohita,Cirrhina
marigala,gill rakers form a broad
sieve like structure across the gill
slits for filtering the water in order
to retain the food in the bucco-
pharynx
19. Carnivorous Fishes : Gill rakers are normally long, hard and
teeth like forming rasping organs as in Wallago attu, Mystus seenghala,
Channa striatus.
20. Gills in Bony fishes
Bony fishes have a single external branchial aperture on each side of the head, beneath
a gill-covering operculam.
Loss of spiracle, reduction of gill slits to four, and deep indentation between the two
hemibranch of each holobrance form a transition from lower (Chondrostei) to higher
(Teleostei) rayfin bony fishes.
The forked efferent branchial artery of each aortic arch of sharks (Chondrosteans) is
replaced by a single efferent vessel in teleost.
In all bony fishes muscles move the bases of the hemibranchs so that the pressure in
and the flow of water from, the oral cavity is regulated.
Strong adductions of the two hemibranchs of a holobranch toward one another occur
in “coughing”.
Coughing is a violent sweeping of water over the gill lamellae to free them from
accumulated detritus.
21. Pseudobranch:
Although lacking of spiracle, rayfin bony fishs retain a hyoidean pseudobranch that is
free in some fishes but skin-covered in others.
This portion of gill region is the first to develop in the embryo and probably has an
early respiratory function.
In adult bony fishes, pseudobranch receives oxygenated blood through a secondarily
established blood supply from the aorta and related blood vessel and also has direct
connect to the choroid gland of the eye.
Pseudobranch is absent in the eels and their relatives, elephant fishes, the cat fishes,
featherbacks and other specialized group in which choroid gland missing.
22. Each filament of a pseudobranch has a thin cartilaginous supporting rod and
blood vessels in its core, around which are arranged layers of pseudobranch
cells and sinusoidal blood spaces.
The large granular pseudobranch cells which dominate the epithelium, are
covered at the outer edge by squamous pavement epithelial cells.
The blood space is supported by pilaster cells, whose thin extensions form
the endothelial lining of blood space.
Within the pseudobranch cells are large numbers of mitochondria and tubular
membrane arrays in close proximity to them. These tubules are extensions of
the plasma membrane and display secretory activity.
This tubules may the site of production of carbonic anhydrase, which
facilitate the formation of carbonic acid from CO2 and water.
23. The gill comprises of gill rakers, gill arch, gill filaments (Primary gill lamellae and
lamellae) (Fig. 5.3a & b).
Each gill filament bears many subdivisions or lamellae that are the main seat of gas
exchange.
In some fishes, (e.g., eel) acidophilic secretory cells occur at the bases of the
lamellae.
The free edge of the lamellae are very thin, covered with epithelium and contain a
vast network of capillaries supported by pilaster cells.
A complete gill is known as holobranch. It consists of a bony or cartilaginous arches.
The anterior and posterior part of each gill arch possesses plate-like gill filaments.
Each holobranch consists of an anterior (oral) and a posterior (aboral) hemi-branch.
24.
25. In teleost fishes, five pairs of branchial arches are present of which first four bear gill
lamellae (Fig. 5.4a, b) but the fifth is devoid of gill lamellae and transformed into the
pharyngeal bone for mastication of food. It does not play any role in respiration.
The gill arch is an important unit and bears primary (gill filament) and secondary
lamellae.
The branchial arch typically consists of paired pharyngobranchials, epibranchials,
ceratobranchials, hypo-branchials and a median unpaired basibranchial.
The epibranchial and the ceratobranchial elements of each branchial arch bears two
rows of gill filaments of the two hemibranchs of the holobranch, which are the seat of
gaseous exchange. It encloses afferent and efferent branchial arteries and veins (Fig.
5.4a and Fig. 5.6a, b, c).
It is also provided by nerves. The branches of 9th (glossopharyngeal) cranial nerve
innervate the first gill, while II, III, & IV arches are supplied by the branches of vagus
(10th cranial nerve). It also contains abductor and adductor muscles. Inside it contains
gill rakers, taste buds, mucous gland cells and sensory papillae.
26. Gill Raker:
It occurs in two rows on the inner margin of each gill arch. Each gill arch is short
stumpy structure supported by bony elements (Fig. 5.3a & b). The gill arch projects
across the pharyngeal opening. They are modified in relation to food and feeding
habits.
The mucous cells of the epithelium help to remove sediments from the covering
epithelium in order to enable the taste buds to function effectively and to sense the
chemical nature of food passing through the gill sieve.
Gill Filaments (Primary Gill Lmellae):
Each hemi-branch consists of both primary and secondary lamellae (Fig. 5.5)
The primary gill filaments remain separated from the branchial septum at their
distal end making two hemi-branch in opposition which direct the water flow
between the gill filaments.
Amongst dual breathers the heterogeneity in the gill system is more pronounced
particularly in the swamp eel, Monopterus, Amphipnous cuchia and climbing perch,
Anabas testudineus.
29. Lamellae (Secondary Lamella):
The each gill filament is made up of secondary gill lamellae which are actual seat of
exchange of gases.
They are generally semicircular and lined up along both sides of the gill filaments. The
lamellae frequency is directly proportional to the dimension and resistance of the gill sieve.
The secondary lamellae are having two sheets of epithelium which are separated by space
and through these spaces blood circulates.
The epithelial sheets are separated by a series of pillar cells. Each cell consists of central
body and is provided with extensions at each end (Fig. 5.6d).
30.
31. Branchial Glands:
These are specialized cells of the epithelium.
They are glandular in nature and perform different functions in normal and
experimental conditions.
The most common specialized branchial glands are the mucous glands and
acidophilic granular cells (chloride cells).
32. Mucous Glands:
These gland cells are unicellular. They may be oval or pear shaped with a neck
through which they open outside the epithelium. The nucleus lies at the bottom of
the cells. They are typical goblet cells. They are present throughout the epithelium,
i.e., gill arch, gill filament and secondary lamellae.
They secrete mucus which is glycoprotein, both acidic and neutral. According to Ojha
and Mishra (1987), acidic and neutral glycoproteins are secreted by same cells
suggesting that they undergo transformation from each other.
Protective
2. Friction reducing
3. Anti-pathogen
4. Help in ionic exchange
5. Help in gas and water exchange.
33. These cells contain granules, which take acidic stain, hence called acidophlic. They
are provided with a large number of mitochondria. These cells are also known as
ioncytes because they help in the ionic regulation in the euryhaline and
stenohaline fishes.
Chloride Cells:
Taste Buds:
They are flask shaped multicellular structure and distributed in the epithelium of
the gill arch region. There are two distinct type of cells, sensory and supporting, and
can be distinguished under phase contrast microscope. They help the fish to sense
the nature of food particles contained in the water flowing through the gill during
ventilation.
34. Respiratory Mechanism:
The continuous undulational flow of water over gill surface is accomplished
by respiratory pump.
It is now unanimously accepted that the respiratory pumps of a teleost consist
of buccal cavity and two opercular cavities, caused by movements of bone of
arches and operculi, resulting in the pumping action of the system.
In the beginning, the water enters into the mouth by expansion of buccal
cavity.
The water is then accelerated over the gill by the simultaneous contraction of
buccal cavity and cavity contracts, expelling the water out through the
opercular opening, the cycle begins again
38. Inspiration:
•Muscular contraction exerts in the floor of the pharynx and buccal
cavity resulting in buccal cavity expanding. The buccal cavity pressure
decreases. Mouth valve opens leading to opening of the mouth and
water with dissolved oxygen enters from the outside because of
reduced pressure.
* At the same time the operculum bulges outwards leading to the
opercular cavity expanding. The opercular cavity acquires a negative
pressure i.e. pressure reduces therefore opercular valve closes.
The expansion of the opercular cavity leads to pressure falling below
that of the buccal cavity which has began to contract as a result water
is sucked into the opercular cavity from buccal cavity and flows over
the gills where gaseous exchange takes place.
39. •Muscular contraction leads to floor of pharynx raising and buccal
cavity continuing to contract. The buccal cavity acquires positive
pressure i.e. pressure increases, mouth valve closes and water in
then forced from buccal cavity to opercular cavity.
•There is inward movement of the operculum. The opercular cavity
contracts and acquires a positive pressure, opercular valve opens and
water containing carbon dioxideis expelled.
The pressure in the buccal cavty is higher than that in the opercular
cavity. Also a continous stream of water is maintained over the gills
throughout most of the breathing cycle by the combined action of the
buccal cavity as a suction pump.
Apart from the time when both the mouth and opercular valves are
open, the pressure in the buccal cavity hence contionous flow of water
from the buccal cavity to opercular cavity.
40.
41.
42. • Fish employ the countercurrent system to extract O2 from
the water.
• This system moves water flowing across the gills, in an
opposite direction to the blood flow creating the maximum
efficiency of gas exchange.
43. • Blood flow through
lamellae is from posterior
to anterior
(back to front).
• Water flow over lamellae is
from anterior to posterior
(front to back).
• Counter-current allows for
diffusion from high O2 in
water to low O2 in blood
across entire length of
lamella.
Countercurrant
Close-up!
44. When the blood and water flows in the same direction, the co-current system, it will
initially diffuses large amounts of oxygen but the efficiency reduces when the fluids start to
reach equilibrium.
In the counter-current system, equilibrium is never reached! Result: Oxygen flow is
always directed into the gills.
% O2 in water
% O2 in gills
% O2 in water
% O2 in gills
45. Ram ventilation is a simpler process in which a fish swims forward with
its mouth open, taking in water that passes over the gills.
The drawback to ram ventilation is that the fish has to swim
continuously to be able to continue breathing.
A lot of sharks breathe this way, as do some large bony fish swimmers
like tunas and billfishes.
Many fishes, including sharks like the sand tiger shark, can switch
between buccal pumping and ram ventilation as the situation and their
swimming speed dictates.
Some sharks and bony fishes, on the other hand, can’t accomplish
buccal pumping at all and have to swim continuously to maintain the
water/blood gas exchange. This is obligate ram ventilation.
The bony fishes include the aforementioned tunas and billfishes.
Obligate ram-ventilating sharks include great whites, makos, salmon
sharks and whale sharks.
Ram ventilation
46. Respiratory structure other than gill
1. Skin:
In some fishes gas exchange takes place across the skin.
Diffusion through the skin is known to play an important role in the
respiration of larval fish-the larvae of Monopterus albus
The skin of adult fish especially those without scale or embedded
with small scales can function as respiratory surface- eels
(Anguilla).
2. Arborescent organ:
Clarias batrachus, the walking catfish has accessory respiratory
organ which consists of the supra-branchial chamber, the two
beautiful rosettes or air trees, the fans and the respiratory
membrane are called arborescent organ.
47. 3. Labyrinth organ:
Anabas, Trichogaster and Channa spp. possess a labyrinth suprabranchial
organ that functions as an air breathing organ.
The organ is the forms of highly vascularised plates which are derived from
branchial tissue.
Air taken in through the mouth passes through the labyrinth where oxygen
is absorbed into the blood.
4. Air sac:
In Heteropneustes fossilis air breathing organs are in the form of a pair, sac
like structure that extends backwards from the region of the gills up to the
middle of the caudal region.
They are thin-walled, long, tubular structure with highly vascular walls and
lie embedded between the myotomes of the body.
The opening of the sac is guarded by valve like stucture and the gill
lamellae on the arches are reduced.
The air sac receive blood from a thick branch of the 4th afferent branchial
vessel of its won side and are drained by means of of the 4th efferent
branchial vessel of the corresponding side.
48. 5. Gut:
In some species, parts of the gastrointestinal tract, notably the stomach or the
small intestine, have modified to serve a role in respiration.
These parts of the gut become highly vascularised which facilitated gas exchange
between the blood and swallowed air.
e.g. members of the families of armoured catfish, Callichthydae and loricaridae.
6. Gas bladder:
The gas bladder is a long cylindrical bag like structure situated within the body
cavity.
In some species, it functions as an accessory respiratory organ. In these species
bladder wall is highly vascularised.
e.g. Giant redfish: Arapaima gigas; Bowfin, Amia calva; Gars Lepisoteus etc.
7. True lung:
In lungfishes (Dipnoi) the gas bladder has evolved structurally to resemble a
lung.
The Australian lung fish Neoceratodus has one lung that lies dorsal to the gut.
In the African (Protopterus) and South American lung fish (Lepidosiren) the lung
is paired but ventral to the gut. Both are obligate air breather.
Like the lung of higher vertebrates, the lung of these fishes are well sacculated
and heavily vascularised.
49.
50. Gas bladder
The gas bladder, less precisely designated as the swim bladder or the air bladder is
characteristics of true fishes and reaches its fullest development as a hydrostatic
organ among spiny rayed fishes.
The gas bladder functions also
as an accessory breathing organ
As a sound producer
As a resonator in sound perception.
In certain deep sea mouth fish relatives the bladder may also serve as a fat-storage
organ of possible buoyant function.
In its weight regulating (hydrostatic) function, gas secretion is accomplished
through a special structure in the bladder wall, the gas secreting complex with its
rete mirabile or wondernet of circulatory vessels and a gas gland.
51. Structure and location
Swim bladders can be paired or unpaired gas-filled sacs.
The pneumatic duct usually connects to the oesophagus ventrally in dipnoans
and chondrostean while dorsally in garfishes, the bowfins, and modern bony
fish.
Sometimes this pneumatic duct is linked with the pharynx or stomach.
Swim bladders are present above the coelomic lining of the peritoneal cavity,
below the dorsal aorta and vertebral column, and lies close to the kidney.
The walls are made up of elastic tissues and smooth muscles.
52. On the basis of the connection of the pneumatic duct with the gut, fish are
divided into two groups:
Physostomous fishes:
These types of fishes have open pneumatic duct i.e., linked with the gut. E.g.
dipnoans, chondrostean, holostean and few teleosts.
Physoclistous fishes:
These types of fishes have closed pneumatic duct i.e., it is not linked or any
connection with gut. e.g., cat fish, carp, eels, herring and salmon.
Gases in swim bladder:
Different fishes contain different gases in their swim bladder. Some fishes have
almost 99% pure nitrogen, while some fishes have 87% oxygen. The swim
bladder of all fishes contains at least traces of four gases i.e., nitrogen, oxygen,
argon, and carbon dioxide.
53.
54. Basic Structure of Swim-Bladder:
The swim-bladder in fishes varies greatly in structure, size and shape:
1. It is essentially a trough sac-like structure with an overlying capillary
network.
2. Beneath the capillary system the wall of the anterior part of swim-bladder
consists of the following layers outside to inside (Fig. 2.38A1).
(a) Tunica externa made up of dense collagenous fibrous material.
(b) Sub-mucosa, consisting of loose connective tissue.
(c) Muscularis mucosa, consisting of a thick layer of smooth muscle fibres.
(d) Lamina propria, formed of thin-layer of connective tissue and
(e) Innermost layer of epithelial cells.
3. In the posterior chamber of swim bladder, outside the layer of muscularis
mucosa there is a glandular layer. This layer is richly supplied with blood
capillaries from rete mirabile (Fig. 2.38A2).
4. The swim bladder opens into the oesophagus by a ductus pneumaticus, which is
short and wide in lower teleosts (Chondrostie and Holostei), while in others it is
longer and narrower. The gas secreted by the swim- bladder is mostly oxygen.
Nitrogen and little quantity of carbon dioxide are also present.
55.
56. Functions of Swim bladder
Hydrostatic organ:
It is primarily a hydrostatic organ and helps to keep the weight of the body
equal to the volume of the water the fish displaces.
It also serves to equilibrate the body in relation to the surrounding medium by
increasing or decreasing the volume of gas content.
In the physostomous fishes the expulsion of the gas from the swim-bladder
occurs through the ductus pneumaticus, but in the physoclistous fishes-where the
ductus pneumaticus is absent -the superfluous gas is removed by diffusion.
Adjustable float:
The swim-bladder also acts as an adjustable float to enable the fishes to swim
at any depth with the least effort.
When a fish likes to sink, the specific gravity of the body is increased.
When it ascends the swim-bladder is distended and the specific gravity is
diminished.
By such adjustment, a fish can maintain an equilibrium at any level.
57. Maintain proper centre of gravity:
The swim bladder helps to maintain the proper centre of gravity by shifting
the contained gas from one part of it to the other and thus facilitates in
exhibiting a variety of movement.
Respiration:
The respiratory function of the swim-bladder is quite significant.
In many fishes living in water in which oxygen content is considerably low,
the oxygen produced in the bladder may serve as a source of oxygen.
In few fishes, specially in the dipnoans, the swim-bladder becomes modified
into the ‘lung’. The ‘lung’ is capable of taking atmospheric air.
Resonator:
The swim bladder is regarded to act as a resonator. It intensifies the
vibrations of sound and transmits these to the ear through the Weberian
ossicles.