анатомия и физиология мужской репродуктивной систем1
1. 7
Анатомия и физиология мужской репродуктивной системы.
По мнению многих исследователей мужская репродуктивная система устроена
значительно сложнее женской и является очень точным маркером любого
патологического процесса в организме.
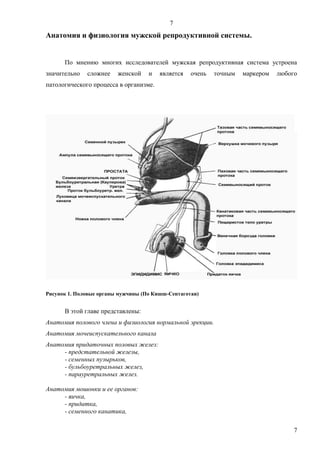
Рисунок 1. Половые органы мужчины (По Кишш-Сентаготаи)
В этой главе представлены:
Анатомия полового члена и физиология нормальной эрекции.
Анатомия мочеиспускательного канала
Анатомия придаточных половых желез:
- предстательной железы,
- семенных пузырьков,
- бульбоуретральных желез,
- парауретральных желез.
Анатомия мошонки и ее органов:
- яичка,
- придатка,
- семенного канатика,
7
2. 8
Анатомия семявыносящего протока.
Анатомия семявыбрасывающего протока.
Анатомия гидатиды яичка, придатка и протоков.
Герминативная и эндокринная функция яичек.
При написании использованы материалы из «Атласа анатомии человека» Р.Д.
Синельникова (1979); «Атласа операций на органах мочеполовой системы» Д. П.
Чухриенко и А.В. Люлько; (1972); материалы из «An Atlas of ERECTILE DYSFUNCTION»
Roger S. Kirby (2005).
Половой член (Penis).
Размеры полового члена очень вариабельны и колеблются от 5-7 см до 10-15 см и
более. В половом члене различают головку (glans penis), тело (corpus penis) и корень (radix
penis), которым прикрепляется связками спереди и ниже лобка к области симфиза.
III V
IV
II
B
Рисунок 2. Строение полового члена
Корень скрыт под мошонкой и фиксирован к костям и диафрагме таза, в результате
чего он является неподвижной, фиксированной частью (pars fixata), остальная часть
полового члена подвижна (pars mobilis). Половой член состоит из двух собственных
пещеристых тел (corpora spongiosum penis) и пещеристого тела мочеиспускательного
8
3. 9
канала (corpus саvernosum urethrae), которое утолщено на обоих концах и образует
спереди головку члена, а сзади — луковицу (bulbus penis).
VI
IV
III
A I
Рисунок 3. Строение полового члена
Кожа полового члена растяжима, легко подвижна. Кожа головки тонка, плотно
сращена с подлежащей белочной оболочкой пещеристого тела головки и поэтому
неподвижна. Кожа полового члена у основания головки собирается в виде складки,
образуя крайнюю плоть (praeputium). Между головкой и крайней плотью образуется
препуциальный мешок, открытый кпереди. На внутреннем листке крайней плоти
расположено большое количество сальных желез. Пещеристые тела полового члена
представляют собой вытянутое в виде цилиндра образование с заостренными концами.
Начинаются они ножками от внутренней поверхности нижних ветвей лобковой и
седалищной костей. У нижней части симфиза ножки срастаются с ним и продолжаются в
тело полового члена, располагаясь по тыльной его стороне. Пещеристые тела соединены
между собой так, что на тыльной и задней их поверхностях образуются углубления; в
первом расположены тыльные сосуды, во втором — пещеристое тело уретры. Пещеристое
тело уретры значительно длиннее и тоньше пещеристых тел полового члена. Внутри его
проходит часть мочеиспускательного канала.
9
4. 10
2
3
4
I
5
Рисунок 4. Поперечные срезы полового члена
Дистальный конец пещеристого тела уретры заканчивается головкой, в углубление
основания которой входят дистальные концы пещеристых тел полового члена. Задний
отдел головки выдается вперед и носит название венчика (corona glandis), позади которого
расположена шейка (collum glandis).
2
1
Рисунок 5. Сагитальный срез полового члена
10
5. 11
Каждое пещеристое тело окружено белочной оболочкой (tunica albuginea). Все три
пещеристых тела покрыты общей фасцией (fascia penis). Поверх фасции расположен слой
рыхлой подкожной клетчатки, а затем кожа.Пещеристые пространства наполнены кровью,
поступающей из аа. profundae penis.
В области симфиза половой член фиксирован двумя связками: поверхностной,
поддерживающей (lig. suspensorium penis s. superficiale) и глубокой, пращевидной (lig.
fundiforme penis). Пещеристые тела у корня полового члена прикрыты мышцами, которые
образуют с каждой стороны по седалищно-луковичному треугольнику. Внутренней
стороной треугольника является луковично-пещеристая мышца (m. bulbo-spongiosus),
которая охватывает нижне-боковую поверхность bulbus penis и срастается по средней
линии с такой же мышцей противоположной стороны. У места их сращения образуется
шов (raphe). Латеральную сторону образует седалищно-пещеристая мышца (m.
ischiocavernosus), а заднюю — поверхностная поперечная мышца промежности (m.
transversus perinei superficialis). Луковично-пещеристая и седалищно-пещеристая мышцы
принимают участие в эрекции.
1
2
3
Рисунок 6. Тазовое дно мужчины
Половой член богато васкуляризован. Артериальную кровь он получает из
внутренней срамной артерии, от которой отходят: артерия луковицы мочеиспускательного
канала (a. bulbi penis), артерия уретры (a. urethralis), глубокая артерия полового члена (a.
11
6. 12
profunda penis) и тыльная артерия полового члена (а. dorsalis penis). A. dorsalis penis
снабжает кровью кожу полового члена, фасции, кавернозные тела, головку и, частично,
кавернозное тело уретры, анастомозируя с a. urethralis. Аа. profunda penis проходят в
толщу пещеристых тел, образуя густую сеть анастомозов с артериями противоположной
стороны. Вены полового члена делятся на поверхностные и глубокие. Поверхностные
вены идут из корня полового члена кнаружи и впадают в большую бедренную вену,
глубокие анастомозируют с поверхностными на головке полового члена и вливаются во
внутреннюю срамную вену. Главной веной является v. dorsalis penis, которая несет кровь в
венозное сплетение мочевого пузыря.
Рисунок 7. Сосуды полового члена
Лимфатические сосуды разделяются на поверхностные и глубокие. Поверхностные
сосуды собирают лимфу из кожи и подкожной клетчатки полового члена, вливаясь в
срединные паховые лимфатические узлы; глубокие образуют в толще головки густую
сеть, от которой стволики идут в кольцевидный лимфатический ствол, откуда лимфа по
truncus dorsalis profundus впадает частично в nodi lymphatici inguinalis superiores et
inferiores, частично в nodi lymphatici iliaci interni.
Иннервация полового члена осуществляется п. dorsalis penis, который является
концевой ветвью срамного нерва (п. pudendus). Он ветвится в коже пещеристых тел и
головке. Nn. erigentes происходят из centrum erectionis, расположенного в боковых рогах
12
7. 13
спинного мозга на уровне I—III сегментов. Симпатические нервные волокна идут от
предстательного сплетения в составе пещеристых нервов (nn. cavernosis penis).
Рисунок 8. Артериальное кровоснабжение полового члена.
Рисунок 9. Венозный дренаж полового члена.
Рисунок 10 Поперечный разрез полового члена
Механизм эрекции управляется автономной нервной системой. Нервные
парасимпатические волокна от S2-4 ответственны за индуцирование эрекции, в то время
13
8. 14
как нервные симпатические волокна от T11-L2 регулируют механизмы эякуляции и
детумесценции. Объединяясь в тазовое сплетение, они образуют кавернозные нервы,
которые, проходя позади предстательной железы, формируют дорсальный нервный
пучок полового члена. Эти нервы и тазовое сплетение часто травмируются при
оперативных вмешательствах на органах таза, что является причиной ЭД в
послеоперационном периоде.
Периферические нервы содержат сенсорные и моторные элементы и формируют
рефлекторную дугу, замыкающуюся в спинном мозге, образуя спиномозговой центр
эрекции.
Достижение полной эрекции зависит от полноценного функционирования как
вегетативной (симпатической и парасимпатической), так и соматической нервной
систем. Таким образом половой член, как и большинство органов человеческого
организма, находится под двойным вегетативным контролем: симпатическая и
парасимпатическая система взаимно дополняют друг друга, действуя на половой член
поочередно.
Механизм эрекции.
Расслабленное состояние.
В расслабленном состоянии половой член небольших размеров, мягко-
эластичной консистенции, что обусловлено низким уровнем притока крови, но этого
уровня достаточно для питания кавернозных тел.
Активность симпатических нервов вызывает сужение артерий и поддерживает
сокращение гладкой мускулатуры пещеристых тел. Низкий кровоток через пещеристую
ткань в состоянии покоя скорее является результатом периферического сопротивления,
чем действия замыкательных механизмов. Существование шунтирующих механизмов
вероятно, но их роль туманна. Следует подчеркнуть, что мускулатура пещеристых тел в
покое активно сокращена и расслабляется во время эрекции.
Эрекция.
Во время эрекции половой член увеличивается в объеме и становится твердым.
Сосудистые изменения могут быть разделены на различные фазы. Различают 5 таких фаз.
1. Латентная фаза. Началу эрекции предшествует расслабление гладкой
мускулатуры артерий и пещеристых тел. Это ведет к падению сосудистого
сопротивления и быстрому притоку крови в пещеристые пространства. В течение
короткого времени возрастания интракорпорального давления нет или даже
наблюдается его небольшое снижение. Этот период изометрического заполнения
синусоидальных пространств связан с наивысшим за время эрекции возрастанием
14
9. 15
кровотока, который может превышать обычный, в состоянии покоя, более чем в
два раза. В этой фазе наблюдаются только незначительное дальнейшее удлинение и
увеличение полноты полового члена.
2. Фаза набухания. По мере продолжения притока крови интракавернозное
давление начинает превышать диастолическое, и с этого момента поступление
крови происходит только во время систолы. Для этой фазы характерны быстрое
удлинение и расширение полового члена до полного объема. Продолжительность
этой фазы зависит от возраста, и на нее влияет также интенсивность стимуляции.
3. Фаза полной эрекции. Продолжающийся приток крови и расширение
синусоидальных пространств придавливают подоболочечные венозные сплетения к
неподатливой белочной оболочке. Это уменьшает отток крови через эмиссарные
вены и еще больше повышает интракавернозное давление. Фаза полной эрекции
достигается при уравнивании интракавернозного и среднего систолического
давления. В этот период давление остается постоянным, свидетельствуя, что
артериальный приток уменьшился по сравнению с фазой набухания и равен
венозному оттоку.
4. Фаза ригидной эрекции. Твердость полового члена достигается сокращением
седалищно-пещеристых мышц, что поднимает интракавернозное давление
значительно выше систолического. На этой стадии артериальный приток в
кавернозной системе прекращается, в то же самое время идет дальнейшее
перекрытие венозных каналов, так что отток через них также приближается к нулю.
В результате на короткий период в несколько минут пещеристые тела становятся
функционально инертными пространствами, практически без притока и оттока
крови. Эта фаза наступает естественно во время полового сношения или
мастурбации, а ее продолжительность ограничена мышечной утомляемостью,
которая устраняет риск ишемии тканей.
5. Фаза детумесценции. Начинается с расслабления седалищно-пещеристых
мышц, однако детумесценция — активный процесс, обусловленный сокращением
кавернозной гладкой мускулатуры под воздействием симпатической иннервации.
Это сокращение выдавливает кровь из синусоидальных пространств и
сопровождается артериальной вазоконстрикцией для снижения притока крови до
уровня покоя. Половой член уменьшается, укорачивается и в итоге становится
вялым.
Как уже упоминалось выше, половой член иннервируют:
1) тазовые нервы (преимущественно парасимпатические);
15
10. 16
2) подчревные нервы (преимущественно симпатические) и
3) половой нерв (преимущественно соматический).
Тазовые и подчревные нервы, объединяясь, образуют тазовое сплетение и дают
начало кавернозным нервам, снабжающим эректильные ткани; половой нерв
иннервирует поперечнополосатую мускулатуру полового члена и воспринимает
чувствительные волокна дорсального нерва полового члена.
.
Рисунок 11. Веноокклюзивный механизм.
В эрекции участвуют три периферических нейрофизиологических механизма:
парасимпатический “сосудистый механизм”, симпатический “тормозящий механизм” и
16
11. 17
соматический “мышечный механизм”. Основная функция автономной нервной системы
(парасимпатического и симпатического отделов) — регуляция тонуса гладкой
мускулатуры артериол и трабекул полового члена. Соматический механизм придает
эрегированному половому члену ригидность. Под воздействием парасимпатических
нервных импульсов происходит расслабление гладкой мускулатуры кавернозных тел и
наступает расширение артериального русла полового члена. Это приводит к повышению
внутрикорпорального давления. Повышение объема крови в синусоидах приводит к
сдавлению венозного сплетения и нарушению венозного оттока. Этот сосудистый
механизм эрекции обеспечивает достижение максимальной ригидности полового члена.
Непосредственно перед оргазмом происходит сокращение седалищно-кавернозных
мышц, приводящее к дополнительной компрессии основания полового члена и
достигается полная эрекция.
Детумесценция.
Расслабление полового члена называется детумесценцией. Она обусловлена
влиянием нервной симпатической системы, которая оказывает сосудосуживающее
действие. Это происходит после оргазма и эякуляции, которые также регулируются
нервной симпатической системой.
Поступление крови в кавернозные тела и ее давление на белочную оболочку
приводит к окклюзии венозного оттока.
Мочеиспускательный канал (urethra masculina)
Мочеиспускательный канал служит для выведения мочи при мочеиспускании, а
также для выведения эякулята в процессе семяизвержения. Нарушение проведения мочи
по мочеиспускательному каналу чаще всего связано с сужением диаметра его отверстия
вследствие воспалительных процессов, травм, опухолевидных разрастаний и т. п.
Воспалительные процессы в предстательной железе и простатической части уретры могут
нарушать запирательную функцию сфинктера мочевого пузыря. При нарушении
последовательности мышечных сокращений при семяизвержении возможен заброс
эякулята в мочевой пузырь (ретроградная эякуляция). В процессе формирования
возможно частичное или полное отсутствие нижней стенки уретры с открытием
мочеиспускательного канала не на головке, а на дорсальной поверхности полового члена.
При недоразвитии верхней стенки возможно открытие уретры на вентральной стороне
полового члена.
Мочеиспускательный канал мужчины представляет собой состоящую из слизистой
и мышечной оболочек узкую трубку длиной 22—25 см, которая служит для выведения
17
12. 18
мочи и семени. Форму канала уретра принимает только во время акта мочеиспускания или
выбрасывания спермы, а в состоянии покоя продольные складки слизистой оболочки
непосредственно прилегают одна к другой. Начинается уретра у основания мочевого
пузыря внутренним отверстием (ostium urethrae internum) и заканчивается на верхушке
полового члена наружным отверстием (ostium urethrae externum). Анатомически она
подразделяется на простатическую часть (pars prostatica), перепончатую (pars
membranacea) и пещеристую (pars cavernosa). Из последней выделяют еще луковичную
часть (pars spongiosa), соответствующую bulbus urethrae, и часть, заключенную в головке
полового члена. С клинической точки зрения весь мочеиспускательный канал принято
разделять на два отдела: передний и задний. Под передним подразумевают пещеристую
часть канала, а под задним — все остальные его части до внутреннего отверстия.
На своем пути уретра образует два изгиба: верхний подлонный изгиб при переходе
перепончатой части уретры в пещеристую вогнутостью обращен кверху и кпереди,
нижний предлонный изгиб формируется при переходе фиксированной части уретры в
подвижную, обращен вогнутостью книзу и кзади. Если половой член приподнят к
передней брюшной стенке, то оба изгиба переходят в общий изгиб, вогнутостью
обращенный вперед и вверх. На всем своем протяжении мочеиспускательный канал имеет
неодинаковый диаметр просвета: чередуются узкие части с более широкими. Одно
сужение находится у внутреннего его отверстия, другое — при прохождении уретры через
мочеполовую диафрагму, третье — у наружного отверстия уретры. Расширений имеется
также три: в простатической части, в луковичной и в конце уретры, где находится
ладьевидная ямка (fossa navicularis). В среднем ширина мочеиспускательного канала равна
4—7 мм.
Простатическая часть уретры расположена вертикально между внутренним и
наружным сфинктерами мочевого пузыря. На всем протяжении она пронизывает
предстательную железу. Длина ее 3—4 см. Внутреннее отверстие уретры охвачено
кольцевидной мускулатурой. На задней стенке простатической части уретры
располагается гребешок мочеиспускательного канала (crista urethralis). В центре его
имеется возвышение — семенной бугорок (colliculus seminalis), на вершине которого
слепое углубление — зачаточная мужская матка (utriculus prostaticus). По бокам от него
находятся устья семявыбрасывающих протоков (ductus ejaculatorii) и протоки
предстательной железы (ductuli prostatici).
Перепончатая часть уретры начинается от конца простатической части и
простирается до места, где уретра входит в пещеристое тело. Это самая короткая и узкая
часть уретры, длина ее 1,5—2,5 см. К задней стенке перепончатой части уретры прилежат
18
13. 19
куперовы железы (glandulae bulbourethrales), выводные протоки которых открываются в
пещеристой части уретры. Перепончатая часть мочеиспускательного канала окружена m.
sphincter urethrae membranaceus, связанной с m. рrostaticus.
Рисунок 12. Нижние мочевыводящие пути
Пещеристая часть мочеиспускательного канала является самой длинной —
начинается от мочеполовой диафрагмы и простирается до наружного его отверстия.
Длина ее равна 12—15 см. У сухожильного центра промежности конец ее колбовидно
расширен (bulbus penis). В заднем отделе пещеристой части уретры открываются
выводные протоки куперовых желез. В передней части, в области головки имеется
ладьеобразная ямка (fossa navicularis). На передней ее стенке имеется складка слизистой
оболочки — заслонка ладьеобразной ямки (valvula fossae navicularis).
На слизистой оболочке уретры имеются углубления — ямки мочеиспускательного
канала (lacunae urethrales) и железы (glandulae urethrales). Мышечный слой
мочеиспускательного канала состоит из гладких и поперечнополосатых мышц.
Мочеиспускательный канал питается мелкими веточками от a. pudenda interna,
которые, анастомозируя между собой, образуют вокруг уретры, в основном в
19
14. 20
подслизистом слое, артериальную сеть. Кроме того, простатическая часть уретры
получает кровь и от a. rectalis media и a. vesicalis inferior, перепончатая — из a. rectalis
inferior и a. perinealis, луковичная — из a. bulbi penis, пещеристая — из a. urethralis. A.
dorsalis penis и а. profunda penis также снабжают своими веточками стенку
мочеиспускательного канала. Вены мочеиспускательного канала образуют сплетения, из
которых кровь поступает в v. dorsalis penis к v. profunda и в вены мочевого пузыря.
Рисунок 13. Анатомия нижних мочевыводящих путей
20
15. 21
Лимфа из пещеристой и перепончатой частей уретры оттекает в nodi lymphatici
inguinalis, а из простатической — в лимфатические сосуды предстательной железы.
Иннервация мочеиспускательного канала осуществляется nn. perineales и п. dorsalis
penis, частью из plexus prostaticus.
Придаточные мужские половые железы.
Предстательная железа (prostata).
Предстательная железа это непарный железисто-мышечный орган, охватывающий
мочеиспускательный канал непосредственно ниже мочевого пузыря и его сфинктеров и
расположенный между мочевым пузырем и мочеполовой диафрагмой. Предстательная
железа вырабатывает слабощелочной опалесцирующий секрет, составляющий 25-35 %
объема семенной жидкости при эякуляции. Секрет предстательной железы имеет сложный
биохимический состав и принимает участие в разжижении эякулята, оказывает буферное
и ферментативное действие на эякулят в целом, активизирует движение сперматозоидов,
оказывает антимикробное действие.
Предстательная железа выполняет также барьерную функцию, препятствуя
проникновению микроорганизмов из уретры в верхние мочевые пути. Выполнение этой
функции обеспечивается содержащимися в секрете железы цинк-пептидным комплексом,
лизоцимом, факторами клеточного иммунитета, IgА, IgG и IgМ. Со всех сторон железа
охватывает простатическую часть мочеиспускательного канала. Форма ее напоминает
каштан. Длина железы 4—4,5 см, ширина 2,5—3 см, толщина 1,4—2 см. Вес железы в
среднем равен 20 г. У мужчин пожилого возраста нередко развивается аденома
предстательной железы, в результате чего ее размеры и вес значительно увеличиваются.
В предстательной железе различают: основание (basis prostatae), обращенное
кверху, верхушку (apex prostatae), обращенную кверху, и, кроме того, переднюю, заднюю
и боковые поверхности. Передняя поверхность направлена к задней поверхности
лобкового сочленения и фиксирована к нему лобково-предстательной связкой (lig.
puboprostaticum). Здесь же к предстательной железе примыкает венозное сплетение (plexus
venosus vesicalis). Задняя поверхность ее прилежит к передней стенке прямой кишки
21
16. 22
1 2 3
B
Рисунок 14.Предстательная железа
. Между ними расположен тонкий слой клетчатки и пузырнопрямокишечная
фасция (septum recto vesicale). По средней линии задней поверхности предстательной
железы имеется бороздка, которая делит ее на правую и левую доли. Между ними
находится перешеек или средняя доля. Своим основанием железа тесно сращена с шейкой
мочевого пузыря. К заднему участку основания предстательной железы примыкают
нижние отделы семенных пузырьков и семявыносящих протоков. Последние вступают в
предстательную железу на ее основании позади уретры в углубление, получившее
название воронки предстательной железы (infundibulum prostatae). Предстательная железа
со всех сторон окружена соединительнотканной капсулой (capsula prostatae), которая
является производной прямокишечной перегородки и наиболее выражена позади, где она
отделяет железу от прямой кишки. Снизу в образовании капсулы принимает участие и
диафрагмальная фасция (fascia diaphragmatis pelvis superior).
22
17. 23
Рисунок 15. Синтопия предстательной железы
. Паренхима простаты состоит из альвеол, которые, группируясь, образуют 30—50
отдельных долек, окруженных фиброзно-мышечными перегородками. Каждая долька
переходит в проток (ductus prostaticus), который точечными отверстиями открывается в
простатическом отделе уретры на боковых поверхностях семенного бугорка. Так как
некоторые протоки объединяются, общее их количество меньше количества долек.
Последние концентрируются в основном в заднем и боковых отделах железы, кпереди от
уретры их почти не бывает.
23
18. 24
Рисунок 16. Поперечный срез предстательной железы
Здесь сильно развита гладкая мышечная ткань, от которой и отходят междолевые
перегородки, собираясь вокруг уретры. Вся эта мышечная ткань получила название
предстательной мышцы (substantia muscularis), которая образует внутренний
непроизвольный сфинктер пузыря.
Рисунок 17. Поперечный срез предстательной железы
24
19. 25
Кровоснабжение предстательной железы происходит из нижних пузырных артерий
(аа. vesicales inferiores), средних геморроидальных (аа. rectales medii), а также
предстательной артерии (a. prostatica), которая отходит от a. pudenda interim. В области
железы все эти артерии между собой широко анастомозируют.
Рисунок 18. Кровоснабжение предстательной железы
Рисунок 19. Схема лимфооттока от предстательной железы
25
20. 26
Вены предстательной железы довольно многочисленны, широко анастомозируют
между собой, образуя мощное венозное сплетение (plexus venosus prostaticus),
располагающееся над предстательной железой. Указанное сплетение принимает кровь из
предстательной железы, мочевого пузыря, семенных пузырьков и семявыносящих
протоков plexus hypogastricus.
Лимфоотток от предстательной железы осуществляется в трех направлениях: 1)
вдоль семявыносящих протоков к подвздошным лимфатическим узлам; 2) к подчревным
лимфатическим узлам; 3) к нижним поясничным лимфатическим узламИннервация
предстательной железы осуществляется волокнами plexus prostaticus, являющимися
непосредственным продолжением plexus hypogastricus inferior, и иннервируют
преимущественно мышечную часть предстательной железы. Секреторные волокна идут от
n. hypogastricus.
Рисунок 20. Иннервация предстательной железы
26
21. 27
Семенные пузырьки (vesiculi seminales).
Семенные пузырьки являются парным железистым органом, состоящим из двух
мешковидных образований, расположенных выше верхнего края предстательной железы.
Секрет семенных пузырьков состоит из желеподобной субстанции, составляющей 50-60 %
семенной жидкости и разжижающейся в течение нескольких минут после эякуляции.
Наиболее важная функция семенных пузырьков - секреция фруктозы, являющейся
источником энергии, метаболизма и поддержания подвижности сперматозоидов.
Рисунок 21. Семенные пузырьки
Семенные пузырьки способны к резорбции жидких компонентов, в них также
происходит лизис сперматозоидов макрофагами. Длина пузырька 6-8 см, ширина 1,5- 2
см, толщина 0,5-1 см. Различают переднюю и заднюю поверхности семенного пузырька.
Передней поверхностью пузырьки прилежат ко дну мочевого пузыря, задней - к передней
стенке ампулы прямой кишки. Между прямой кишкой и семенными пузырьками лежит
пузырно-прямокишечная перегородка. Семенные пузырьки состоят из тела (corpus
vesiculae seminalis) и шейки (collum vesiculae seminalis). Верхняя треть семенных
пузырьков покрыта брюшиной, остальная их часть лежит внебрюшинно. Шейка семенных
27
22. 28
пузырьков переходит в выводной проток (ductus excretorius), открывающийся в боковую
стенку ампулы семявыносящего протока. Над телом семенных пузырьков проходит
нижняя часть мочеточников. Вверху семенные пузырьки расходятся, а внизу почти
соприкасаются.
Рисунок 22. Иннервация семенных пузырьков
Кровоснабжение семенных пузырьков происходит из нижних пузырных артерий
(аа. vesicales inferiores) и средних геморроидальных артерий (аа. rectales mediae). Вены
семенных пузырьков образуют plexus venosus vesicalis et prostaticus, откуда отток крови
осуществляется по трем направлениям: кпереди в срамное сплетение (plexus pudendalis),
28
23. 29
кверху и в стороны в венозное сплетение мочевого пузыря (plexus vesicalis), кзади в
геморроидальное сплетение.
Лимфатические сосуды густо анастомозируют между собой, с лимфатическими
сосудами мочевого пузыря, предстательной железы и вливаются в подчревные
лимфатические сосуды. Иннервируются семенные пузырьки волокнами из подчревного
симпатического сплетения (plexus hypogastricus) и частично ветвями из п. pelvicus
Бульбоуретральные (Куперовы) железы (glandulae bulbourethrales)
Бульбоуретральные железы - парный железистый орган около 1 см в диаметре
шаровидной формы. Протоки от альвеол сливаются в более крупные и превращаются в
общий выводной проток длиной 3-6 см, оканчивающийся двумя щелевидными
отверстиями в луковичном отделе уретры. Секрет этих желез представляет собой
бесцветную прозрачную, лишенную запаха слизь со щелочной реакцией, выделяющуюся
в просвет уретры при половом возбуждении за счет сокращения мышц промежности. При
прохождении по уретре секрет нейтрализует кислую реакцию, остающейся в ней мочи и,
выделяясь из наружного отверстия мочеиспускательного канала, обеспечивает введение
полового члена во влагалище. В пожилом возрасте отмечается гипотрофия желез.
Парауретральные (Литтре) железы.
Парауретральные железы - мелкие гроздьевидные трубчато-альвеолярные железы
расположены в подслизистом слое уретры на всем ее протяжении, выделяют слизистый
секрет, количество которого увеличивается при половом возбуждении. Как и секрет
бульбоуретральных желез, он смачивает уретру, поддерживая в ней щелочную среду.
Рисунок 23. Строение парауретральных желез
29
24. 30
Мошонка (scrotum) и ее органы.
В связи с тем, что сперматогенез в норме протекает при температуре на 2-4 градуса
ниже температуры тела, мошонка выполняет две основные функции - создание
оптимального температурного режима для яичек и защита органов мошонки от
механической травматизации. Таким образом при сокращении объема мошонки
осуществляются ее терморегулирующая и защитная функции.
Рисунок 24. Органы мошонки
30
25. 31
Мошонка представляет собой кожно-мышечное мешковидное образование,
разделенное перегородкой (septum scroti) на две половины, каждая из которых является
вместилищем для яичка, придатка и мошоночного отдела семенного канатика.
Располагается она между корнем полового члена и передним отделом
промежности. Мошонка является редуцированной частью передней брюшной стенки;
слои ее — видоизмененное продолжение слоев передней брюшной стенки; среди них
различают: 1) кожу (derma); 2) мясистую оболочку (tunica dartos); 3) fascia spermatica
externa — продолжение поверхностной фасции живота; 4) fascia cremasterica; 5) m.
cremaster; 6) fascia spermatica interna — продолжение fascia transversalis; 7) tunica vaginalis
testis.
Кожа мошонки тонка, гладка, хорошо растяжима, собрана в складки, более темной
пигментации, чем кожа живота и бедер, покрыта редкими волосами. По средней линии ее
проходит шов (raphe scroti), который спереди переходит на кожу полового члена, сзади —
на кожу промежности. Подкожная клетчатка отсутствует. Непосредственно под кожей
лежит мясистая оболочка, представляющая собой слой гладких мышечных волокон,
имеющих примесь соединительных волокон. Эта оболочка на передней брюшной стенке
соответствует подкожной клетчатке; она плотно фиксирована к коже и представляет с ней
как бы один слой. С мошонки tunica dartos переходит в фасцию полового члена,
поверхностную фасцию живота, подкожную клетчатку промежности и внутреннюю
поверхность бедер. Из мясистой оболочки образуется перегородка мошонки (septum
scroti).
Под мясистой оболочкой лежит fascia spermatica externa, являющаяся
продолжением собственной фасции живота. Она представляет собой слой рыхлой
клетчатки, отделяющий мясистую оболочку от кремастера. Мышца, поднимающая яичко,
происходит из поперечной и косой мышц живота. Проходит она через наружное паховое
кольцо, затем, покрывая семенной канатик, вместе с ним опускается вниз к яичку и
обхватывает его. Под m. cremaster лежит fascia spermatica interna, являющаяся
продолжением поперечной фасции живота и представляющая собой
соединительнотканную пластинку с примесью волокнистой ткани. Внутренней
поверхностью она прилежит к tunica vaginalis testis. Последняя происходит из брюшины и
образует вокруг яичка серозный мешок, в котором различают два листка: пристеночный
(lamina parietalis), выстилающий изнутри общую влагалищную оболочку, и висцеральный
(lamina visceralis), покрывающий яичко и придаток. В нижней части мошонки имеется lig.
scrotale, остаток gubernaculum testis.
31
26. 32
Кровоснабжение кожи мошонки осуществляется из аа. scrotalis anteriores (от аа.
pudendae externae) и аа. scrotalis posteriores (от a. perinealis). Остальные слои получают
кровь из a. cremasterica. Вены мошонки образуют богатую сеть, из которой кровь оттекает
в v. v. pudendae extern., венозные сплетения семенных канатиков (pl. pampiniformes и vv.
rectales inferiores).
Лимфатические сосуды мошонки впадают в поверхностные паховые узлы.
Иннервация мошонки осуществляется из нескольких источников — срамного,
поясничного и крестцового сплетений. Спереди проходят передние нервы мошонки (nn.
scrotales anteriores) из ветвей n. ileoinguinalis, сзади — задние нервы мошонки (nn. scrotalis
posteriores) из ветвей п. pudendus. М. cremaster иннервируется ветвями n. genitofemoralis.
Яичко (testis).
Яичко — парный железистый орган мужчины. Длина его равна 4—5 см, ширина
2—2,5 см, толщина 2,5—3 см. Средний вес 15—25 г. Различают верхний конец яичка
(extremitas superior), обращенный кнаружи, и нижний (extremitas inferior), обращенный
внутрь; наружную (fades lateralis) и внутреннюю (fades medialis) поверхности; передний
(margo anterior) и задний (margo posterior) края.
Рисунок 25. Яичко
32
27. 33
У верхнего полюса и на заднем крае находится придаток яичка (epididymis). Яичко
имеет две оболочки: влагалищную (tunica vaginalis testis) и белочную (tunica albuginea).
Белочная оболочка — это плотная соединительнотканная пластинка, окутывающая
все яичко; на заднем крае она образует треугольную складку (гайморово тело),
вдающуюся в толщу яичка (mediastinum testis), от которого веерообразно расходятся
соединительнотканные перегородки (septula testis).
Рисунок 26. Оболочки яичка
Всю паренхиму яичка перегородки делят на 250-300 долек (lobuli testis), которые
состоят из 1—2 извитых семенных канальцев (tubuli seminiferi contorti) 0,2-0,3 мм
диаметром и длиной 30-45 см. На верхушке дольки канальцы, соединяясь, образуют узкие
прямые семенные трубочки (tubuli seminiferi recti), которые, прорастая гайморово тело,
образуют сеть семенной железы (rete testis). Здесь прямые канальцы соединяются в 10—15
более крупных выносящих канальцев (ductuli efferentes testis), которые прободают
белочную оболочку и образуют так называемый придаток яичка.
Местом образования сперматозоидов являются извитые канальцы, собственная
оболочка которых выстлана сустеноцитами (клетками Сертоли) и зародышевым
эпителием, из которого, собственно, и развиваются сперматозоиды. В межуточной ткани
между извитыми канальцами расположены соединительно-тканные элементы, сосуды,
нервы, а также интерстициальные эндокриноциты (гландулоциты, Клетки Лейдига),
33
28. 34
вырабатывающие мужские половые гормоны: андрогены (среди которых основным
является тестостерон) и эстрогены (главным образом - эстрадиол). Под влиянием
тестостерона происходит маскулинизация как всего организма, так и половых органов,
развитие их по мужскому типу.
Основной объем яичка составляет канальцевая система паренхимы, в которой и
осуществляется процесс сперматогенеза. На стенках канальцев недифференцированные
первичные половые клетки (сперматогонии) размножаются в течение 74 суток и, проходя
через ряд делений (сперматоциты 1-го порядка, сперматоциты 2-го порядка или
пресперматиды, сперматиды, обладающие уже гаплоидным набором хромосом),
превращаются в зрелые сперматозоиды. Сустеноциты, выстилающие мембрану семенных
канальцев, обеспечивают сперматогенные клетки продуктами своей секреторной
активности, выполняют фагоцитирующую функцию в отношении остатков процесса
сперматогенеза, синтезируют эстрогеноподобное вещество - ингибин - выделяют
андрогенсвязывающий протеин, способствующий переносу тестостерона и
дегидротестостерона в половые клетки, где они закрепляются в ядре. По мере созревания
клетки сперматогенного эпителия смещаются от базальной мембраны к просвету канальца
между цитоплазматическими отростками сустеноцитов, постепенно трансформируясь в
процессе деления в сперматозоиды.
В зародышевом эпителии яичек образуется фермент гиалуронидаза,
локализующийся, преимущественно, в головках сперматозоидов (который в дальнейшем
растворяет цервикальную слизь и разъединяет клетки лучистого венца яйцеклетки).
Другим продуктом секреции являются простагландины, обеспечивающие сокращение
гладкой мускулатуры женских половых путей и повышающие скорость прохождения
яйцеклетки навстречу сперматозоидам.
Базальная мембрана канальцев создает гематотестикулярный барьер,
предотвращающий распознавание иммунной системой наличия антигенов на поверхности
генеративных клеток и угнетения процесса сперматогенеза вследствие аутоиммунного
поражения этих клеток антителами. Кроме того, базальная мембрана снижает уровень
поражения сперматогенеза при воспалительных заболеваниях, интоксикациях и т. п.
Гормональная регуляция функции яичек осуществляется через соотношение
гормонов, вырабатываемых в гипоталамусе, гипофизе и яичках по принципам прямой и
обратной связи.
Кровоснабжение яичка и придатка осуществляется из a. testicularis и частично из a.
cremasterica, а семявыносящего протока— из a. deferentialis. Отток крови совершается
34
29. 35
через венозное сплетение plexus pampiniformis, которое входит в состав семенного
канатика и впадает в семенную вену.
Рисунок 27. Срез яичка
Лимфатические сосуды яичка разделяют на глубокие и поверхностные. Глубокая
лимфатическая сеть находится в соединительной ткани между извитыми канальцами,
дольками и в гайморовом теле, а поверхностные сосуды сосредоточены в белочной
оболочке и придатке яичка. Лимфатические капилляры поверхностной и глубокой сети
взаимно переплетаются с кровеносными сосудами. Лимфа оттекает по лимфатическим
сосудам семенного канатика в регионарные лимфатические узлы, расположенные на
аорте, нижней полой вене, передней поверхности подвздошной вены.
Иннервация яичка и придатка осуществляется за счет plexus testicularis et plexus
deferentialis. В составе сплетений проходит также значительное количество и
чувствительных волокон.
Придаток (epididymis).
Придаток— продолговатый орган, прилежащее к задней поверхности яичка. Длина
его 5—6 см, ширина 2—2,5 см, толщина 0,5—1 см. В его более крупную верхнюю часть
(головку придатка) из средостения яичка проникают 12-15 канальцев, в средней части
35
30. 36
(теле придатка) они начинают объединяться и из нижней части (хвоста придатка) выходит
один общий семявыносящий проток. В придатке яичка под действием андрогенов
происходит окончательное морфологическое, биохимическое и физиологическое
дозревание сперматозоидов (продолжающееся 14 дней). В придатке сперматозоиды
освобождаются от цитоплазматической капли, обволакиваются защитной белковой
оболочкой, приобретают отрицательный заряд насыщаются рядом веществ (жиры,
холестерин, фосфаты, корнитин и т. д.), происходит ряд ультраструктурных и
цитохимических преобразований акросомы. Слабое напряжение кислорода и отсутствие
фруктозы препятствуют активации сперматозоидов. Хвост придатка является их
хранилищем. При половом воздержании старые формы могут лизироваться
спермиофагами, предотвращая переполнение и разрушение канальцевой системы яичек
при обтурационной аспермии.
Рисунок 28. Придаток яичка
В придатке различают головку (caput), тело (corpus) и хвост (cauda), который
переходит в семявыносящий проток (ductus deferens).
Головка образована выносящими канальцами, представляющими собой
продолжение протоков галлеровой сети. Протоки соединяются и образуют один общий
проток (ductus epididymidis), который в конце хвоста переходит в семявыносящий проток
(ductus deferens).
36
31. 37
Головка придатка расположена на верхнем конце яичка и сращена с ним.
Поверхность придатка покрыта висцеральной пластинкой влагалищной оболочки, которая
переходит на него с яичка. К яичку придаток фиксирован в области головки и хвоста
связками: верхней (lig. epididymidis superior) и нижней (lig. epididymidis inferior). Между
телом яичка и придатком у места перехода влагалищной оболочки имеется углубление —
пазуха придатка (sinus epididymidis).
Рисунок 29. Придаток яичка
Семенной канатик (funiculus spermaticus).
Семенной канатик— парный орган, который представляет собой кругловатой
формы образование, начинающийся от придатка яичка и заканчивающийся у внутреннего
отверстия пахового канала, откуда отдельными своими составными элементами вступает
в полость таза. Длина его равна 15—20 см, ширина — 2—5 см. У плодов мужского пола
он образуется вследствие опускания яичек с их протоком, сосудами, нервами и
окружающими оболочками из полости живота в мошонку.
В состав семенного канатика входят: 1) cемявыносящий проток (ductus deferens); 2)
артерия семявыносящего протока (a. ductus deferentis); 3) внутренняя семенная артерия (a.
37
32. 38
testicularis); 4) гроздьевидное (лозовидное) сплетение (plexus pampiniformis); 5) vv.
testicularis et deferentialis; 6) vasa lymphatica; 7) нервное сплетение — plexus deferentialis et
plexus testicularis и 6—И отводящих лимфатических сосудов. Все эти элементы окружены
рыхлой клетчаткой, которая связывает их в один пучок. В толще ее заключены гладкие
мышечные лоскуты. В заднем отделе пучка лежит семявыносящий проток. Кпереди от
него расположена a. spermatica interna, сзади — a. deferentialis.
Семявыносящий проток (ductus deferens).
Семявыносящий проток является продолжением ductus epididymidis и соединяется
с верхушкой семенного пузырька, где переходит в ductus ejaculatorius. Это плотная, легко
ускользающая из-под пальцев мышечная трубка длиной 30— 40 см. Диаметр ее равен 2—
3 мм, просвет самого канала не превышает 0,5 мм.
Рисунок 30. Семявыносящий проток
Стенка состоит из 3 слоев: tunica adventitia, tunica muscularis et tunica mucosa.
Артериальная система семенного канатика слагается из трех источников:
внутренней семенной артерии (a. testicularis), артерии семявыносящего протока (a. ductus
deferentis), артерии канатика (r. genitalis) — ветви нижней надчревной артерии. Вены
яичка и придатка образуют обширное венозное сплетение, получившее название
гроздевидного (plexus pampiniformis). Вверху вены этого сплетения разделяются на
переднюю часть, идущую по ходу a. spermatica interna, и заднюю, расположенную по ходу
v. deferentialis. Постепенно сливаясь, они образуют семенную вену (v. spermatica), по
38
33. 39
которой кровь справа вливается в нижнюю полую, а слева — в левую почечную вену.
Лимфатические сосуды проходят между артериями и венами, анастомозируют между
собой и впадают в поясничные лимфатические узлы.
Иннервация яичка, его придатка и семенного канатика, как упоминалось выше,
осуществляется из нескольких источников. К ним следует отнести п. spermaticus externus,
являющийся ветвью п. genitofemoralis и plexus spermaticus, которые отходят от нижней
части почечного сплетения, и plexus deferentialis, отходящего от пузырного сплетения.
Парасимпатические волокна идут в составе симпатического сплетения, беря начало от
тазового сплетения.
Семявыбрасывающий проток (ductus ejaculatorius).
Семявыбрасывающий проток - парный трубчатый орган длиной 2-3 см,
образующийся при слиянии ампулы семявыносящего протока и семенных пузырьков и
открывающийся в области маточки семенного бугорка.
Гидатиды яичка, придатков, протоков (Жиральдов орган).
Это некрупные образования округлой или удлиненной формы на ножке или плотно
прикрепленные к яичку или придатку яичка, являющиеся остатками различных
образований процесса эмбриогенеза.
Герминативная и эндокринная функция яичек.
Сперматогенез является одним из наиболее динамичных процессов в организме
человека, связанных с клеточной регенерацией и дифференцировкой. Он протекает под
контролем специфических генов развивающихся гамет и регулируется совокупностью
гормонов, цитокинов и факторов роста. В сутки у взрослого мужчины в яичке образуется
около 100-200 млн спермиев, что соответствует продукции примерно 70-150 тыс. клеток в
минуту или 15 000 спермиев с каждым биением сердца. Вместе с тем в эякулят поступает
значительно меньшее количество спермиев, чем то, что начально образуется в семенных
канальцах яичка, вследствие их частичной гибели в самом яичке и в семявыносящих
путях. В последних, в особенности в придатке яичка, происходит дозревание и
накопление этих клеток.
39
34. 40
Процесс образования сперматозоидов начинается в подростковом возрасте и
протекает до глубокой старости. Процесс созревания одного сперматозоида занимает 72
дня.
Рисунок 31 Поперечный разрез канальца яичка (по Stieve)
Для этого необходимы следующие условия:
- извитой каналец, где происходит созревание сперматозоидов,
- участие клеток Сертоли,
- высокая местная концентрация гормона тестостерона,
- температура яичек на 2 градуса по Цельсию ниже, чем температура тела.
Канальцевый отдел. В канальцевом отделе, занимающем около 60-80-90% общего
объема яичек, происходит сперматогенез. В этом отделе присутствуют зародышевые
клетки и два разных вида соматических клеток – околоканальцевые клетки и клетки
Сертоли. Соединительнотканные перегородки делятяичко примерно на 250-300 долек,
каждая из которых содержит от 1 до 3 сильно извитых семенных канальцев. Всего в яичке
человека содержится около 600 семенных канальцев. Длина отдельных канальцев
колеблется от 30 до 80 см, а их общая протяженность в каждом яичке достигает 360 м.
Таким образом, у человека имеется 720 м семенного эпителия.
40
35. 41
Клетки Сертоли представляют собой постоянную популяцию неделящихся клеток
микроокружения. Они расположены на базальной мембране семенных канальцев и
соединены между собой плотными соединениями (tight junctions). Эти плотные
соединения вместе с тесно взаимосвязанными между собой мышечными клетками
перитубулярного пространства формируют гемато-тестикулярный барьер.
Функциональное значение гемато-тестикулярного барьера заключается в создании
уникального микроокружения, необходимого для обеспечения сперматогенеза, и
поддержании иммунологической привилегированности яичка. Такая изоляция очень
важна, так как образование сперматозоидов происходит с периода полового созревания,
что намного позже становления функции самораспознавания иммунной системы. Если бы
образование сперматозоидов не было иммунологически защищено, то
дифференцирующиеся клетки распознавались бы иммунной системой, как чужеродные, и
уничтожались бы.
Клетки Сертоли участвуют, как в обеспечении питания развивающихся клеток
сперматогенеза, так и в фагоцитозе погибших клеток. Сперматогонии и молодые
сперматоциты расположены в семенных канальцах ближе к базальной мембране и по мере
созревания перемещаются в более верхние слои, вглубь просвета канальцев.
Соматические клетки Сертоли расположены среди зародышевого эпителия. В зрелом
возрасте митозы в них отсутствуют. Эти клетки названы по имени итальянского ученого
Энрико Сертоли (1842-1910), впервые описавшего их в 1865 г.
Длинные цитоплазматические отростки и разветвления определили и второе их
название: "ветвистые клетки". Они покоятся на базальной мембране, выступают в просвет
семенных канальцев и могут считаться структурами, во всех отношениях
поддерживающими зародышевый эпителий. Все морфологические и физиологические
изменения зародышевой клетки в процессе ее дифференцировки и созревания вплоть до
образования зрелого сперматозоида происходит вдоль тела клетки Сертоли, тянущейся на
всю высоту зародышевого эпителия. Расположение и пространственная ориентация
сперматозоидов в ходе их дифференцировки поддерживаются особыми эктоплазмати
чески ми выростами. Клетки Сертоли занимают около 35-40% объема зародышевого
эпителия. Интактное яичко с полностью завершенным сперматогенезом содержит 800-
1200х106 клеток Сертоли. Эти клетки синтезируют и секретируют множество различных
факторов: белки, цитокины, ростовые факторы, опиоиды, стероиды, простагландины,
модуляторы клеточного деления и т.д. Принято считать, что клетки Сертоли организуют
процесс сперматогенеза пространственно и функционально. С другой стороны, последние
данные указывают на роль зародышевых клеток в регуляции функций клеток Сертоли.
41
36. 42
Клетки Сертоли выполняют и другую важную функцию: именно они определяют
окончательный объем яичек и продукцию спермы у взрослых. Каждая клетка Сертоли
морфологически и функционально связана с определенным числом сперматозоидов. Это
число зависит от вида животного. У человека на каждую клетку Сертоли приходится 10
зародышевых клеток или 1,5 сперматозоида. Это позволяет считать, что в пределах вида
большее количество клеток Сертоли обусловливает более высокую продукцию
сперматозоидов, если, конечно, все клетки Сертоли функционируют нормально. Какие же
факторы определяют степень пролиферации клеток Сертоли и тем самым объем яичек у
взрослых? У человека до 15-летнего возраста число этих клеток прогрессивно
увеличивается.
Андрогены или ФСГ увеличивают и количество клеток Сертоли, и экспрессию ими
маркеров клеточного деления. Появление гематотестикулярного барьера совпадает с
началом первого мейоза зародышевых клеток (прелептотена, зиготена) и с прекращением
пролиферации клеток Сертоли. Барьер делит семенной эпителий на две области,
совершенно разные как анатомически, так и функционально. Зародышевые клетки на
ранних стадиях развития располагаются в базальной области, а на более поздних –
прилежат к просвету канальцев. В ходе созревания они смещаются из базальной в
прелюминальную область, что сопровождается синхронным размыканием и повторным
смыканием плотных контактов выше и ниже мигрирующих зародышевых клеток.
Гематотестикулярный барьер выполняет две важные функции: физически
изолирует гаплодные и тем самым антигенные зародышевые клетки, препятствуя их
распознаванию иммунной системой, и создает особую среду для мейоза и созревания
сперматозоидов. В этот момент между клетками Сертоли образуются плотные контакты,
формирующие так называемый гематотестикулярный барьер. Производя и секретируя
канальцевую жидкость, клетки Сертоли создают и поддерживают в открытом состоянии
просвет канальцев. Туда попадает более 90% жидкости, секретируемой этими клетками.
Особые структурные элементы гематотестикулярного барьера препятствуют реабсорбции
этой жидкости, благодаря давлению которой просвет канальцев остается открытым.
Сперматозоиды перемещаются в канальцевой жидкости, состав которой отличается от
состава крови большей концентрацией ионов калия и меньшим содержанием ионов
натрия. Кроме того, в этой жидкости присутствуют ионы бикарбоната, магния и хлора,
инозитол, глюкоза, карнитин, глицерофосфорилхолин, аминокислоты и ряд белков.
Герминальные, или сперматогенные, клетки располагаются в семенных канальцах
строго определенным образом. Сперматогонии лежат непосредственно над базальной
мембраной, а в в направлении просвета канальцев последовательно располагаются
42
37. 43
первичные сперматоциты, вторичные сперматоциты и сперматиды. Выделяют 13
различных типов клеток герминогенного эпителия, представляющие разные стадии
созревания сперматозоидов. Сперматогенез начинается с деления стволовых клеток и
заканчивается образованием зрелых сперматозоидов. В семенном канальце различные
зародышевые клетки группируются в соответствии со стадиями своего созревания (стадии
сперматогенеза). Весь процесс сперматогенеза можно разделить на три фазы:
• митотическая пролиферация и дифференцировка диплоидных зародышевых клеток
(сперматогонии),
• мейотическое деление тетраплоидных зародышевых клеток (сперматоциты),
• трансформация гаплоидных зародышевых клеток (сперматид) а зрелые сперматозоиды
(спермиогенез).
Рисунок 32. Сперматогонии, сперматоциты и сперматиды в клетках Сертоли
Сперматогонии расположены в основании семенного эпителия и среди них
различают клетки А- и В-типа. Цитологические, а теперь уже и физиологические данные,
позволяют разделить клетки А-типа на два вида: Ат (темные) сперматогонии и Аб
(бледные) сперматогонии. Ат-клетки в нормальных условиях лишены пролиферативной
43
38. 44
активности и поэтому могут считаться стволовыми клетками сперматогенеза. Но при
резком уменьшении общего количества сперматогониев, например вследствие облучения
в Ат-клетках начинаются митозы. В отличие от них, Аб-сперматогонии делятся, и каждый
из них превращается в два В-сперматогония. Из В-сперматогониев, еще до начала мейоза,
прямо образуются сперматоциты прелептотены, в которых начинается синтез ДНК.
Материнские и дочерние клетки сохраняют между собой тесный контакт с помощью
межклеточных мостиков. Такой "клональный" способ развития зародышевых клеток, по-
видимому, является основой и в то же время необходимым условием координированного
созревания гамет в семенном эпителии.
Тетраплоидные зародышевые клетки, называемые сперматоцитами, проходят
различные фазы мейоза. Фаза пахитены характеризуется интенсивным синтезом РНК. В
результате мейотического деления образуются гаплоидные зародышевые клетки –
сперматиды. Мейоз – центральное событие гаметогенеза. В ходе этого процесса
рекомбинируется генетический материал, уменьшается число хромосом и завершается
развитие сперматид. В результате первого деления мейоза образуются вторичные
сперматоциты, содержащие двойной гаплоидный набор хромосом. При втором мейозе
сперматоциты делятся на гаплоидные сперматиды. Профаза первого мейоза длится 1-3
нед., тогда как другие его фазы и весь процесс второго мейоза продолжаются всего 1-2
дня.
Сперматиды, образующиеся при втором мейозе, представляют собой круглые
митотически неактивные клетки, которые в результате сложной трансформации в конце
концов превращаются в дифференцированные удлиненные сперматиды и сперматозоиды.
Эти процессы включают конденсацию и структурное оформление клеточного ядра,
появление жгутика и освобождение от большей части цитоплазмы. Весь процесс носит
название спермиогенеза и с точки зрения качества одинаков у всех видов. Спермиогенез
целесообразно разделить на 4 фазы: фазу Гольджи, фазу формирования головки,
акросомную фазу и фазу созревания.
Во время фазы Гольджи появляются акросомные пузырьки и возникает
краниокаудальная симметрия. В фазу формирования головки сперматида удлиняется и
развивается акросома, покрывающая половину-две трети клетки с головного конца. В
процессе оплодотворения акросома высвобождает ферменты, обеспечивающие
возможность проникновения сперматозоида в яйцеклетку.
В течение акросомной фазы клеточное ядро еще больше конденсируется, и клетка
продолжает удлиняться. При конденсации большинство гистонов теряется, и
транскрипция генов прекращается. Ядерный хроматин становится очень плотным.
44