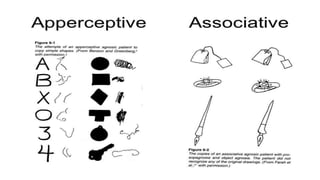
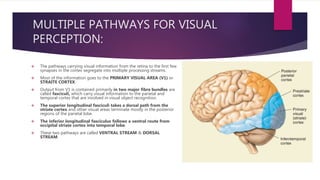
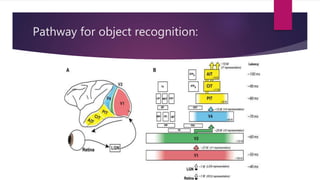
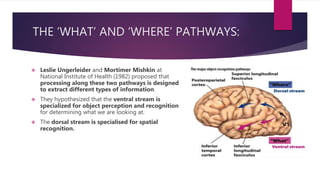
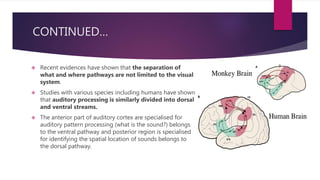
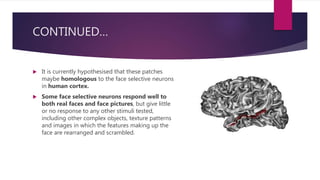
This document discusses visual cognition and object recognition. It begins by outlining the ability of the visual system to perceive and recognize objects despite changes in size, position, color, texture or motion. Object recognition involves hierarchical representations that combine simple features into more complex shapes and objects. The document then divides visual processing into low, intermediate and high level components involving extracting physical properties, detecting global properties, and recognizing and classifying objects. It provides the example of visual agnosia, an impairment in object recognition, and discusses the famous case of patient G.S. who could describe shapes but not recognize objects visually. The document outlines the pathways for visual processing in primates, including the ventral and dorsal streams, and their roles in object ("