1. See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/316842076
Pomicultură generală
Book · November 2011
CITATIONS
0
READS
6,940
2 authors, including:
Some of the authors of this publication are also working on these related projects:
AGRAL 146/2002 – Research concerning the achievement an efficient model of farm (orchards) for implementation in similar zone and the opportunity to attract the
young farmers in fruit growing View project
ADER 16.1.1. - Adaptation of innovative technologies applicable to local (cooperative) production, conditioning and storage centers for short and long-term fresh and
processed horticultural products View project
Dorin Ioan Sumedrea
Research Institute for Fruit Growing, Pitesti
41 PUBLICATIONS 22 CITATIONS
SEE PROFILE
All content following this page was uploaded by Dorin Ioan Sumedrea on 10 May 2017.
The user has requested enhancement of the downloaded file.
2.
3. Dorin Sumedrea Mihaela Sumedrea
POMICULTURĂ
GENERALĂ
Referenţi ştiinţifici: Prof. asoc. dr. Mihail IANCU, Membru titular ASAS
Prof. dr. Alexandru TEODORESCU
ISBN 978-973-1886-60-2
4. Dedic această carte mamei mele care trece în aceste momente
printr-o grea suferinţă. Mama mi-a oferit un dar de preţ – dorinţa de
a excela. Toată viaţa a muncit, luptându-se cu greutăţile, pentru a
crea condiţii mai bune pentru cei trei copii ai săi, ne-a învăţat să
reuşim prin muncă, corectitudine, ne-a învăţat să fim tari şi să ţinem
capul sus.
Piteşti, 11.12.2011
Dorin Ioan SUMEDREA
5. 3
C U P R I N S
CAPITOLUL 1. DEFINIŢIA, IMPORTANŢA, SITUAŢIA ACTUALĂ ŞI
TENDINŢELE DEZVOLTĂRII POMICULTURII ……………… 007
1.1. Definiţie ………………………………………………………………………. 007
1.2. Importanţa economică şi socială a pomiculturii ……………………………. 007
1.3. Situaţia actuală şi tendinţele dezvoltării pomiculturii ……………………… 010
CAPITOLUL 2. SPECIILE POMICOLE DE CLIMAT TEMPERAT.
TAXONOMIE, NOMENCLATURĂ, CLASIFICARE …………….. 015
2.1. Taxonomie …………………………………………………………………… 015
2.2. Clasificarea speciilor pomicole ……………………………………………… 015
2.2.1. Clasificarea după habitus …………………………………………… 015
2.2.2. Clasificarea pomicolă ……………………………………………….. 017
CAPITOLUL 3. MORFOLOGIA ŞI FIZIOLOGIA POMILOR ŞI ARBUŞTILOR
FRUCTIFERI ……………………………………………………….. 019
3.1. Organele hipogee …………………………………………………………….. 019
3.2. Organele epigee ..…………………………………………………………….. 024
3.3. Polaritatea şi dominanţa apicală …………………………………………….. 030
CAPITOLUL 4. CICLUL DE VIAŢĂ AL SPECIILOR POMICOLE ……………. 032
4.1. Perioadele de vârstă ………………………………………………………….. 032
4.2. Comportarea pomilor pe rădăcini proprii şi a celor altoiţi în ciclul de viaţă …. 035
4.3. Relaţiile dintre epibiont şi hipobiont ………………………………………… 037
4.3.1. Altoirea şi utilizarea ei în pomicultură ……………………………… 037
4.3.2. Influenţa reciprocă dintre epibiont şi hipobiont ……………………. 041
CAPITOLUL 5. CICLUL ANUAL AL SPECIILOR POMICOLE ………………. 044
5.1. Fenofazele iniţiale ale organelor vegetative şi de rod ………………………... 044
5.1.1. Fenofazele iniţiale ale organelor vegetative …………………………. 044
5.1.2. Fenofazele iniţiale ale organelor de rod ……………………………… 045
5.2. Starea de repaus ………………………………………………………………. 047
5.3. Fenofazele finale ale organelor vegetative şi de rod …………………………. 050
5.3.1. Fenofazele finale ale organelor vegetative …………………………... 050
6. 4
5.3.2. Fenofazele finale ale organelor de rod ……………………………… 052
CAPITOLUL 6. ALTERNANŢA DE RODIRE …………………………………….. 060
6.1. Cauze şi forme de manifestare .……………………………………………… 060
6.2. Măsuri de prevenire a alternanţei de rodire …………………………………. 061
CAPITOLUL 7. ECOLOGIA POMILOR ŞI ARBUŞTILOR FRUCTIFERI ……. 063
7.1. Particularităţile agroecosistemului pomicol ………………………………… 063
7.2. Lumina ca factor de vegetaţie ……………………………………………….. 064
7.3. Căldura ca factor de vegetaţie ………………………………………………… 067
7.4. Apa ca factor de vegetaţie …………………………………………………… 070
7.5. Aerul ca factor de vegetaţie ………………………………………………….. 071
7.6. Cerinţele speciilor pomicole faţă de sol ………………………………………. 072
7.7. Relieful şi distribuţia factorilor ecologici …………………………………… 075
CAPITOLUL 8. REGIUNILE POMICOLE ALE ROMÂNIEI …………………... 077
CAPITOLUL 9. PORODUCEREA MATERIALULUI SĂDITOR POMICOL ….. 080
9.1. Factorii în funcţie de care se amplasează pepiniera ………………………… 080
9.2. Sectoarele pepinierei …………………………………………………………. 080
9.3. Organizarea şi pregătirea terenului …………………………………………… 081
9.4. Producerea portaltoilor generativi …………………………………………... 082
9.5. Producerea portaltoilor vegetativi …………………………………………… 086
9.5.1. Tehnologia înmulţirii prin marcotaj ………………………………… 086
9.5.2. Tehnologia înmulţirii prin butaşi …………………………………… 089
9.5.3. Înmulţirea prin altoire ………………………………………………. 091
9.5.4. Înmulţirea prin drajoni ……………………………………………… 097
9.5.5. Înmulţirea prin micropropagare „in vitro” …………………………. 097
9.6. Tehnologia producerii pomilor altoiţi ……………………………………….. 098
9.7. Metode de scurtare a timpului pentru producerea materialului săditor pomicol 102
7. 5
CAPITOLUL 10. TEHNOLOGIA ÎNFIINŢĂRII PLANTAŢIILOR
POMICOLE ……………………………………………………….. 104
10.1. Sisteme de cultură a pomilor ………………………………………………. 104
10.2. Alegerea terenului …………………………………………………………... 113
10.3. Organizarea, amenajarea şi pregătirea terenului …………………………... 114
10.4. Alegerea şi amplasarea speciilor şi soiurilor ……………………………….. 116
10.5. Stabilirea distanţelor de plantare şi pichetarea terenului …………………... 117
10.6. Plantarea pomilor şi lucrări de îngrijire după plantare ……………………. 123
CAPITOLUL 11. TEHNOLOGIA ÎNTREŢINERII PLANTAŢIILOR
POMICOLE ……………………………………………………….. 126
11.1. Operaţiile tehnice folosite pentru dirijarea creşterii şi rodirii …………….. 126
11.1.1. Tăierile şi influenţa acestora asupra proceselor de creștere şi rodire 126
11.1.2. Clasificarea tăierilor ………………………………………………… 127
11.1.3. Schimbarea poziţiei ramurilor ………………………………………. 129
11.1.4 Operaţii tehnice secundare …………………………………………. 132
11.2. Forme de coroană …………………………………………………………… 133
11.2.1. Coroane cu volum mare …………………………………………… 135
11.2.2. Coroane cu volum mic ..…………………………………………… 141
11.3. Tăierile de formare a coroanelor …………………………………………… 155
11.4. Tăierile de producţie ……………………………………………………….. 156
11.4.1. Tăierile de întreţinere ……………………………………………… 157
11.4.2. Tăierile de fructificare ……………………………………………… 158
11.5. Tăierile de creştere a coroanelor …………………………………………… 159
11.6. Mecanizarea tăierii pomilor ………………………………………………… 161
11.7. Întreţinerea şi lucrărilor solului în plantaţiile pomicole …………………... 163
11.8. Fertilizarea plantaţiilor pomicole ………………………………………….. 168
11.8.1. Consumul şi cerinţele speciilor pomicole pentru elementele nutritive
în cursul ciclului anual ……………………………………………… 169
11.8.2. Criterii de stabilire a normelor de fertilizare ………………………. 170
11.8.3. Îngrăşăminte utilizate în pomicultură …………………………….. 172
11.8.4. Tehnica administrării îngrăşămintelor …………………………….. 175
11.9. Irigarea plantaţiilor pomicole ………………………………………………. 176
11.9.1. Consumul de apă al speciilor pomicole …………………………… 176
8. 6
11.9.2. Constantele hidrografice ale solului utilizate la stabilirea necesarului
de apă ……………………………………………………………….. 177
11.9.3. Stabilirea momentelor şi normelor optime pentru irigarea pomilor 178
11.9.4. Metode de irigare …………………………………………………... 179
11.10. Protejarea pomilor împotriva accidentelor climatice …………………….. 181
11.11. Îngrijirea recoltelor ………………………………………………………… 183
11.11.1. Reglarea încărcăturii de rod ………………………………………. 183
11.11.2. Prevenirea căderii fructelor înainte de recoltare ………………….. 185
11.11.3. Reglarea hormonală a maturării şi îmbunătăţirea calităţii fructelor 185
11.12. Recoltarea fructelor ……………………………………………………….. 186
11.12.1. Estimarea producţiei de fructe …………………………………… 186
11.12.2. Epoca optimă de recoltare a fructelor …………………………….. 187
11.12.3. Tehnica recoltării …………………………………………………. 188
BIBLIOGRAFIE SELECTIVĂ ………………………………………………………. 191
9. 7
Capitolul 1
DEFINIŢIA, IMPORTANŢA, SITUAŢIA ACTUALĂ ŞI
TENDINŢELE DEZVOLTĂRII POMICULTURII
1.1. DEFINIŢIE
Pomicultura este o ramură a ştiinţelor agricole, unul dintre principalele sectoare ale
producţiei horticole. Etimologic, această denumire derivă din latinescul pomus, -i = arbore
fructifer şi cultur, -ae = îngrijire, cultivare.
Pomicultura constituie un domeniu al “resurselor agricole” care prin interacţiunea
naturală (cu factorii de mediu) şi dirijată (cu factorii agrotehnici) are ca rezultat obţinerea
unor sortimente de produse agricole - fructele - vital necesare existenţei umane.
Ca ştiinţă, alături de întreaga ştiinţă agricolă a luat naştere după consolidarea ştiinţelor
fundamentale: fizica, chimia, botanica, fiziologia, biochimia, etc.
Într-o accepţiune modernă, pomicultura, ca resursă, reprezintă un subsistem conectat
printr-o multitudine de legături, directe şi indirecte, cu celelalte resurse agricole, împreună cu
care se constituie în sistemul agricol, dar şi cu alte sisteme (industrial, transporturi,
construcţii, servicii, etc.), precum şi cu mediul, în cadrul ecosistemului. Agroecosistemul
pomicol este în directă dependenţă de condiţiile ecologice locale: legile şi principiile generale
se aplică numai în raport de condiţiile locale concrete. Din acest motiv, pomicultura poate fi
definită ca fiind “Studiul particularităţilor biologice şi productive ale pomilor şi arbuştilor
fructiferi şi stabilirea tehnologiilor diferenţiate pe specii, soiuri şi zone ecologice pentru
obţinerea unor recolte mari de fructe cu valoare biologică superioară, în condiţii de eficienţă
economică ridicată (Popescu M. şi colab., 1992).
În învăţământ, ca disciplină didactică, pomicultura abordează aspectele ştiinţifice şi
practice legate de cultura speciilor fructifere. Aceasta este divizată în Pomicultura generală şi
Pomicultura specială (Pomologia), prima cuprinzând aspecte legate de biologia , ecologia şi
tehnologia comună tuturor speciilor fructifere; pomicultura specială aprofundează pe cea
generală, prin studierea particularităţilor de creştere şi fructificare ale diferitelor specii, grupe
de soiuri şi chiar soiuri, în vederea stabilirii tehnologiei diferenţiate a acestora.
1.2. IMPORTANŢA ECONOMICĂ ŞI SOCIALĂ A POMICULTURII
Principala raţiune a culturii pomilor o constituie folosirea în alimentaţie a fructelor, ca
un component esenţial pentru asigurarea funcţiilor metabolice echilibrate ale organismului
uman.
Importanţa economică şi socială a culturii pomilor poate lua în considerare
următoarele aspecte:
Fructele - alimente indispensabile. Cu puţine excepţii, datorită echilibrului şi
armoniei dintre diferitele lor elemente (forme, mărimi, culori arome, gusturi, componente
chimice, etc.), fructele reprezintă aproape singurul aliment gata pregătit în natură, în stare
finită, care se poate consuma proaspăt, fără nici un adaos sau proces de prelucrare. Orice
operaţiune de prelucrare scade valoarea alimentară specifică fructelor.
Valoarea alimentară a fructelor este dată de compoziţia chimică a acestora, în forme
uşor accesibile organismului uman, la care se adaugă diferiţi excitanţi olfactivi, vizuali şi
gustativi.
10. 8
Din punct de vedere chimic fructele conţin apă şi substanţă uscată (substanţe organice
şi minerale). Conţinutul fructelor proaspete în apă variază în funcţie de specie în limite largi:
73,5% la prune, 84,1% la mere, 91,4% la piersici, 93,0% la căpşun şi numai 4,0% la alune.
Zaharurile reprezintă principalul component al substanţei uscate (circa 90%). Acestea
sunt prezente, în principal, sub forme de monozaharide (glucoza, fructoza), dizaharide
(zaharoza) şi polizaharide (celuloza, amidonul, pectina). Conţinutul acestora în fructe variază
de la 3,40% la agrişe, până la 16,72% la mere. Zaharurile din fructe, mai ales cele simple, sunt
asimilate de organismul uman foarte uşor, intrând în circuitul sanguin, fără transformări
prealabile care să necesite consum energetic. Trebuie subliniată “sănătatea“ pentru organism a
zahărului din fructe în comparaţie cu cel obţinut prin prelucrare industrială, mai ales prin
dublă rafinare (Isac Il., şi colab., 2001).
Grăsimile şi proteinele se găsesc în cantităţi reduse în fructele cărnoase (mere, pere,
prune, caise, piersici, cireşe, vişine): 0,1 - 0,7% lipide (ceride şi cutină) şi 0,1 - 0,4% proteine.
Cantităţile reduse de proteine sunt compensate printr-o diversitate mare. La măr, de exemplu,
s-au identificat 26 aminoacizi, printre care şi cei esenţiali, pe care organismul uman nu-i poate
sintetiza. La fructele uscate (nuci, alune, migdale) conţinutul în lipide depăşeşte 50%, iar cel
în proteine variază în jur de 18-20% (Miliţiu I., 1992).
Celuloza, conţinută în proporţie de 0,8 - 1,0%, joacă un rol mecanic, în sensul că
stimulează peristaltismul intestinal şi favorizează digestia.
Valoarea nutritivă a fructelor este sporită de conţinutul lor în vitamine şi săruri
minerale, care sunt biocatalizatori ai proceselor vitale, lipsa lor din organism provocând grave
tulburări funcţionale ale metabolismului.
În ordinea descrescândă a cantităţii, fructele conţin următoarele vitamine: vitamina C,
care la majoritatea fructelor variază între 0,5 şi 40 mg., dar ajunge la 150- 400 mg. la 100 g
substanţă proaspătă în coacăzele negre; provitamina vitaminei A (caroten 0,02 - 0,09 mg la
100 g, şi poate ajunge la 4,8 mg în pulpa caiselor şi 1.8 mg. în cea a piersicilor); vitaminele B1
(alune, nuci, castane, prune), B2 (castane, nuci, caise, căpşuni); biotina (mere, pere); vitamina
PP (mere, cireşe, caise, piersici), etc.(Radu I. F., 1985). Dintre toate acestea, cea mai mare
importanţă îi revine vitaminei C. Datorită faptului că organismul uman nu poate sintetiza
această vitamină, omul este obligat să-şi procure zilnic cantităţile necesare, în principal prin
consumul de alimente vegetale. În timpul iernii o parte din fructe (mere, pere) devin, dacă nu
singura, în orice caz una din puţinele surse de vitamina C.
Substanţele minerale se găsesc sub formă de oxizi ai principalelor metale (K, Na, Ca,
Fe, etc) sau săruri ale acizilor fosforic, clorhidric, carbonic, etc. Conţinutul în săruri minerale
variază de la o specie la alta, în funcţie de soi, dar şi de condiţiile pedoclimatice, de
tehnologiile aplicate, etc. Deşi în cantităţi mici, substanţele minerale au un rol deosebit în
organismul uman influenţând secreţia diferitelor glande şi servesc ca substanţe tampon în
metabolismul intern, în special al sucului gastric (Isac Il., 2001).
Fructele - medicamente valoroase. Datorită proprietăţilor terapeutice fructele sunt
utilizate nu numai în alimentaţia omului sănătos ci şi în regimurile alimentare recomandate,
atât de medicina populară cât şi de cea ştiinţifică, multor categorii de bolnavi.
În special, datorită conţinutului în celuloză fructele ajută digestia; aciditatea lor
moderată provoacă o importantă secreţie de salivă, stimulează sucurile gastrice şi reglează
funcţiile intestinale. De asemenea, sunt renumite ca deconstipante merele, perele, caisele,
piersicile (Popescu M., şi colab., 1992).
Fructele au o acţiune benefică asupra diurezei, asupra eliminării acidului uric, motiv
pentru care sunt recomandate reumaticilor, obezilor, etc.
Fructele foarte bogate în pectină (merele, perele, gutuile) pot reduce colesterolul, fiind
recomandate în prevenirea arteriosclerozei şi a infarctului miocardic. Consumul de fructe este,
11. 9
de asemenea, unul din mijloacele de corectare a hipertensiunii arteriale (Popescu M., şi colab.,
1992).
Materii prime pentru industria prelucrătoare. Datorită caracterului sezonier al
producţiei de fructe, al dezechilibrului dintre consum şi producţie din unele perioade, cât şi
nevoii aportului de fructe în alimentaţie în cursul anului, industria prelucrătoare capătă o
importanţă deosebită. Sortimentele de produse în care se transformă fructele sunt tot mai
diversificate: sucuri naturale, concentrate, nectaruri, siropuri, băuturi răcoritoare, compoturi,
jeleuri, gemuri, marmelade, dulceţuri, bomboane, îngheţată, fructe deshidratate, congelate,
etc.
Pe plan mondial se manifestă tendinţa de sporire a proporţiei consumului de fructe sub
formă prelucrată în comparaţie cu cele proaspete. În SUA, în ultimele decenii ale secolului
trecut structura medie a consumului de fructe a fost de 40% proaspete şi 60% prelucrate (din
care 33% conservate, 22% congelate, 5% uscate).
În România, în aceiaşi perioadă, structura consumului de fructe a fost de 77% în stare
proaspătă şi 23% prelucrate (21% conservate, 0,5% congelate, 1,5% uscate). Dacă se ia în
calcul producţia de fructe care se prelucrează pentru export, inclusiv cea care se distilează,
rezultă că în activitatea de industrializare a fructelor poate fi luată în considerare o cantitate
anuală de 500-600 mii tone materii prime (Isac Il., 2001).
Exportul de fructe. În condiţii normale producţia de fructe din ţara noastră
depăşeşte nevoile de consum intern, existând posibilitatea exportului de fructe în stare
proaspătă (căpşuni, prune, mere, pere, etc.) sau prelucrată (căpşuni, cireşe, vişine, caise,
piersici, zmeură, coacăze, etc.). Însă, în anii 1992-1993, România exporta doar 16 mii tone
fructe (producţia medie de fructe în aceşti ani a fost de 1675 mii tone), în special prune pentru
consum în stare proaspătă şi mere pentru industrie (Papp J., 1996). Pe piaţa merelor de
consum, România precum şi ţările din Europa de Est, nu pot fi exportatori importanţi, datorită
sortimentului cultivat şi neîndeplinirii cerinţelor de calitate cerute pe piaţa occidentală.
Valorificarea mai eficientă a unor terenuri comparativ cu alte culturi.
Importanţa economică a pomiculturii este accentuată de posibilităţile pe care le au speciile
pomicole de a pune în valoare terenuri cu fertilitate scăzută: terenurile în pantă din zona
dealurilor, precum şi cele nisipoase din sudul şi nord-vestul ţării. Sistemul radicular dezvoltat
al speciilor pomicole valorifică şi astfel de terenuri, asigurând în condiţiile unor tehnologii
adecvate producţii eficiente. În plus, se mai adaugă şi suprafeţele mici de teren situate în
condiţii care nu permit cultura plantelor de câmp, cum sunt cele din jurul caselor.
Pretabilitatea valorificării economice şi estetice a terenurilor din grădinile de
lângă casă. Pomii şi arbuştii fructiferi sunt printre cele mai indicate plante pentru cultura pe
loturi mici: asigură o productivitate mare (4-5 kg fructe/m2
); pot asigura o gamă sortimentală
de fructe proaspete în gospodărie prin eşalonarea epocilor de coacere a speciilor şi soiurilor
cultivate; oferă o ocupaţie plăcută, folositoare şi reconfortantă pentru membrii familiei.
Aceste considerente explică de ce peste 87 milioane pomi (58% din patrimoniu) sunt cultivaţi
în gospodăriile populaţiei (Isac Il., 2001). De asemenea, prelucrarea fructelor în gospodărie,
sub diferite forme, a devenit o îndeletnicire de tradiţie, acoperindu-se în acest fel o parte
importantă din necesarul de consum.
Funcţia climatică şi de conservare a solului. Prezenţa plantaţiilor pomicole
contribuie activ la modificarea, de multe ori esenţială, a regimului şi calităţii climatului.
Plantaţiile pomicole asigură o compoziţie normală a atmosferei prin consumul de CO2 şi
eliberarea de oxigen, atenuează efectul unor agenţi poluanţi (praf, fum, etc.) şi extremele de
temperatură, măresc umiditatea relativă, micşorează viteza vânturilor, favorizează depunerea
zăpezii, etc. (Isac Il., 2001). În plus, plantaţiile pomicole au un important rol antierozional,
intervenind activ în conservarea solului prin protecţia împotriva eroziunii de suprafaţă şi
adâncime, prin evitarea şi împiedicarea alunecărilor de teren, prin fixarea nisipurilor, etc.
12. 10
Ocuparea populaţiei în activitatea pomicolă. Suprafaţa pe care o ocupă
plantaţiile de pomi şi arbuşti fructiferi, producţia de fructe de cca. un milion tone realizată
anual în ţara noastră, absorb o parte a populaţiei care lucrează fie direct la aplicarea
tehnologiilor în livezi, fie în activităţi conexe (industria prelucrătoare, comerţul cu fructe,
transporturi, construcţia de maşini şi unelte specifice, industria chimică, etc.).
1.3. SITUAŢIA ACTUALĂ ŞI TENDINŢELE DEZVOLTĂRII POMICULTURII
Analizate pe o perioadă lungă de timp, pe plan mondial, producţiile de fructe au
crescut continuu, la început pe măsura extinderii suprafeţelor cultivate, iar în ultimele decenii
datorită sporirii producţiei la unitatea de suprafaţă. De la 58.220 mii tone, cât era media anilor
1948-1952, producţia mondială a principalelor specii pomicole de climat temperat a ajuns la
70.935,6 mii tone, media anilor 1980-1989. În perioada anilor ’90 producţia totală de fructe
din zona temperată depăşeşte 100.000 mii tone, ajungând la sfârşitul primului deceniu al
mileniului 3 la peste 130.000 mii tone tabelul 1.1. Creşterea producţiei mondiale de fructe se
datorează în cea mai mare parte tendinţei permanente de creştere la specia măr.
Două treimi din producţia mondială de fructe este furnizată de 3 specii, şi anume:
mărul, bananierul şi portocalului. În plus, mărul este specia care furnizează peste 50% din
producţia de fructe de climat temperat (tabelul 1.1), fiind urmat de păr cu (17%), piersic şi
nectarin (15%), prun (8%), etc.
Având în vedere aceste considerente vom analiza în continuare producţia mondială de
mere şi structura acesteia pe continente şi respectiv ţări mari producătoare.
Tabelul 1.1
Evoluţia producţiei mondiale de fructe la principalele specii pomicole
de climat temperat
Specia
Media
1980-1989
Nivelul producţiei în anii (mii tone)
1997 1998 2001 2002 2008 2009
TOTAL 70935,6 100223,3 100147,9 97863,1 115712,3 133431,1 138867,1
Măr 39194,4 57692,3 56886,5 58673,1 57982,6 69304,4 71216,6
Păr 9418,0 14185,7 15132,6 17312,0 17198,1 21201,6 22460,5
Piersic şi nectarin 8012,5 10994,3 10996,5 1330,3 20000,0 18670,2 20319,3
Prun 6339,9 7808,2 7177,2 9183,3 9141,8 11163,8 10776,2
Căpşun 2060,0 2683,4 2701,5 3168,4 3099,1 3716,5 3728,1
Cais 2002,8 2346,2 2544,5 2590,6 2738,6 4093,7 4178,2
Cireş 1471,9 1616,0 1605,5 1926,8 1948,4 1801,3 2196,5
Nuc 845,3 1100,1 1122,8 1280,1 1267,9 161,5 830,0
Alun 507,3 607,8 771,5 660,8 590,7 1062,1 765,7
Castan 455,8 539,8 522,7 971,9 962,6 1330,4 1408,3
Gutui 321,6 329,6 330,9 385,8 400,3 467,9 503,3
Zmeur 306,1 319,9 355,7 380,0 382,2 457,7 484,4
Sursa: FAO STAT (http://apps.fao.org)
Din analiza producţiei mondiale de mere şi repartizarea ei pe continente se observă că
(tabelul 1.2), la nivelul anilor 1989-1991, din producţia totală de 40.118 mii tone, 32,5% se
realizează în Europa, urmata de Asia (29,8%), America de Nord (13,7%), URSS (13,5%),
America de Sud (5,9%), Africa (2,3%), Oceania (1,7%) şi Australia (0,6%).
Începând cu ultima parte a anilor ‘90 producţia mondială de mere prezintă o tendinţă
de creştere continuă (tabelul 1.2), ajungând 2009 la o producţie de 71216 mii tone, cu 77,5%
mai mare decât media anilor 1989-1991.
În Europa se observă o creştere a producţiei de mere, ajungând la 16.111 mii tone în
anul 2009, ceea ce reprezintă o creştere cu 23,7% faţă de media anilor 1989-1991.
13. 11
După 1990 Europa prezintă creşteri uşoare ale producţiei de mere, însă creşteri foarte
mari sunt înregistrate în Asia, care devine primul producător mondial. La nivelul anului 2009
ponderea producţiei de mere pe continente situează Asia pe primul loc cu 60,9%, urmată de
Europa cu 22,6%, America de Nord (6,9%), America de Sud (5,0%), Africa (2,9%), etc. Asia
cunoaşte o creştere spectaculoasă a producţiei de mere, care ajunge la 43.422 mii tone în
2009, de aproximativ 3,6 ori mai mare decât media anilor 1989-1991.
Tabelul 1.2
Producţia mondială de mere şi repartizarea ei pe continente
mii tone
1989-1991 1997 1998 2001 2002 2008 2009
Prod. mondială
din care:
40118 57692 56887 58673 58785 693047 71216
Europa
Asia
America de Nord
America de Sud
Africa
Australia şi Noua Zeelandă
13027
11956
5488
2381
907
263
18613
27669
5844
3779
1526
920
15836
29538
6172
3445
1465
832
16879
30695
5301
3974
1522
775
15822
31164
5030
4136
1546
812
15933
41711
4716
3563
2132
621
16111
43422
4928
3561
2064
642
Sursa: FAO STAT (http://apps.fao-.org)
Creşterea producţiei în Asia se datorează în primul rând Chinei (tabelul 1.3), ţară în
care producţia de mere a crescut de peste 7 ori, de la 4469 mii t media anilor 1989-1991, la
31.684 mii tone în 2009. Alţi mari producători ai acestui continent sunt Turcia, Iran (peste 2
mil. tone fiecare), şi India (cu peste 1 mil. tone).
În America de Nord, principalul producător - SUA are tendinţa de menţinere a
producţiei anuale de mere la aproximativ 4,5 milioane tone, iar America de Sud este
reprezentată de Argentina, Chile şi Brazilia (peste 1 milion tone în 2009).
În Europa, ţări mari producătoare sunt Polonia şi Italia (peste 2 milioane tone anual),
Franţa (cca 2 milioane tone anual), Rusia, Germania (fiecare cu peste 1 milion tone anual).
Tendinţa generală a ţărilor din Europa de Vest este de creştere uşoară şi chiar de stagnare a
producţiei de mere.
În România, după 1990 în urma aplicării legii fondului funciar este dificil de apreciat
suprafaţa şi producţia de mere obţinută de actualii proprietari. De asemenea, trebuie avute în
vedere pierderile mari datorate defrişărilor făcute de către noii proprietari, neexistând o
evidenţă precisă a acestui aspect.
Evoluţia suprafeţelor ocupate cu pomi se caracterizează printr-o reducere după 1938 şi
până în anii ‘70, datorită celui de al doilea război mondial şi colectivizării forţate din anii ‘60
(tabelul 1.4). În 1970 se atinge un maxim (328.4 ha), după care are loc o diminuare, ce se
datorează defrişării unor livezi îmbătrânite, precum şi a unor livezi mici situate pe terenurile
arabile din zonele de şes, supuse politicii agrare din acea perioadă (înfiinţarea livezilor se
făcea numai pe terenurile în pantă, slab fertile şi erodate).
Se apreciază că, după anul 2000, suprafeţele pomicole din România au scăzut sub 200
mii ha, fiind acceptată idea că producţia de fructe nu trebuie să crească pe seama sporirii
suprafeţelor, ci a randamentului la unitatea de suprafaţă (Glăman Gh., 1996) şi în condiţiile în
care România a negociat cu UE numai 180 mii ha. Livezile de măr reprezintă peste o treime
din suprafaţa totală ocupată cu livezi pe rod (tabelul 1.4).
În perioada 1991-2009 producţia totală de fructe obţinută în România se situează în
jurul valorii de 1,1-1,3 mil. tone, cu excepţia anului 1993 când se depăşeşte producţia de
2 mil. tone (Tabelul 1.5).
14. 12
Tabelul 1.3
Producţia de mere în principalele ţări cultivatoare din lume
mii tone
ŢARA 1989-1991 1997 1998 2001 2002 2008 2009
EUROPA
Franţa
Italia
Rusia
Polonia
Ucraina
Germania
Spania
România
Ungaria
Olanda
AMERICA DE NORD
S. U. A.
Canada
AMERICA DE SUD
Argentina
Chile
Brazilia
ASIA
China
India
Iran
Japonia
Turcia
AFRICA
Africa de Sud
Egipt
Maroc
2113
1935
-
1090
-
1957
661
628
921
369
4434
531
963
713
516
4469
1118
1378
953
1883
485
57
255
„
2473
1966
1500
2092
1898
1602
984
664
500
420
4682
504
1118
940
794
17228
1308
1998
993
2550
538
403
384
2910
2143
1330
1687
568
2296
736
364
482
518
5283
489
1034
1000
791
19491
1321
1944
879
2450
576
388
285
2397
2341
1643
2434
475
1929
962
507
605
480
4363
465
1428
1135
723
20023
1500
2353
930
2450
558
474
228
2500
2370
1800
1900
482
1600
712
500
470
500
4042
460
1428
1100
858
20508
1500
2353
912
2500
579
484
228
1940
2210
1467
2831
719
1047
661
459
567
375
4359
427
950
1280
1124
29851
1984
2719
911
2504
771
551
404
1954
2314
1596
2626
853
1071
595
517
575
407
4515
473
1027
1090
1223
31684
1795
2432
846
2782
550
702
400
Sursa: FAO STAT (http://apps.fao.org)
Tabelul 1.4
Evoluţia suprafeţelor ocupate cu pomi
ANUL
Suprafaţa-total livezi
pe rod/d.c.măr (mii ha)
ANUL
Suprafaţa-total livezi pe
rod/d.c.măr (mii ha)
1938
1950
1960
1970
1980
1990
1991
1999
2000
247,0
184,2
212,6
328,4
259,7
230,7/90,8
232,8/88,3
208,9/80,5
208,1/71,0
2001
2002
2003
2004
2005
2006
2007
2008
2009
210,1/73,0
189,4/72,4
199,6/71,6
199,8/73,4
200,7/81,7
178,7/59,3
167,6/59,0
166,8/54,7
159,6/52,4
Sursa: FAO STAT (http://apps.fao.org) şi M.A.A.P. - Date operative
Dacă în anii ’50 prunele deţineau peste 50% din producţia totală, tendinţa de creştere a
producţiei de mere începând cu anii ‘80 se menţine şi în ultimul deceniu al secolului XX, când
producţia de mere depăşeşte pe cea de prune. Datorită investiţiei mai mici la înfiinţare şi
costurilor uşor mai mici în exploatare înregistrate la specia prun, la sfârşitul anilor ‘90 s-au
înfiinţat mai multe livezi de prun decât de măr şi, ca urmare producţia de prune în anii 2008-
2009 depăşeşte uşor pe cea de mere.
15. 13
Tabelul 1.5
Producţia de fructe realizată în România în perioada 1991-2009
Specia 1991 1992 1993 1996 1997 2000 2001 2002 2008 2009
TOTAL PROD. 1.164,7 1.167,0 2.183,0 1.631,8 1.416,5 1093,7 1349,6 1260,4 1183,8 1323,3
Mere 504,9 541,1 1.097,2 659,7 664,1 490,3 507,0 500,0 459,0 517,5
Pere 58,1 63,1 108,5 74,2 69,9 70,6 71,6 65,0 52,6 66,1
Prune 418,9 346,7 703,7 663,0 491,6 345,0 557,2 500,0 475,3 533,7
Piersici 43,6 37,3 49,7 25,9 17,4 18,3 16,7 12,0 16,4 17,1
Caise 26,0 40,7 41,7 43,8 27,6 28,4 28,3 25,0 32,1 32,5
Cireşe + Vişine 59,6 73,0 106,4 89,3 73,8 73,7 91,1 85,0 67,7 67,9
Nuci 18,0 21,8 32,6 35,7 32,9 32,5 34,9 34,0 32,3 38,3
Alte specii 21,8 30,5 15,9 28,5 26,0 23,0 24,0 21,0 25,0 26,0
Arbuşti fructiferi 2,6 1,8 1,6 0,7 0,4 0,2 0,4 0,4 2,2 2,3
Căpşuni 13,8 12,8 7,3 11,6 13,5 11,7 18,4 18,0 21,2 21,9
Sursa: FAO STAT (http://apps.fao.org) şi M.A.A.P. - Date operative
Dacă analizăm producţia de mere, pe o perioadă mai lungă de timp (tabelul 1.6), se
observă o scădere foarte mare în 1950 când se obţine numai 13,5% din producţia anului 1938,
datorită războiului şi secetei. Producţia anului 1938 (566,5 mii tone) este depăşită după anul
1980, când se obţin în mod constant aproximativ 600 mii tone anual, cu excepţia anului 1986
(994,4 mii tone).
După 1990 producţia de mere oscilează între 300-600 mii tone cu excepţia anilor cu
producţii foarte mari: 1993 şi 2004 când s-au înregistrat producţii record de peste1 milion
tone sau 2003 cu peste 800 mii tone.
În ultima perioadă sporurile de producţie obţinute la nivel mondial nu se datorează
extinderii suprafeţelor, ci intensivizării culturii mărului. Tendinţa este de a defrişa livezile
extensive şi de a înfiinţa livezi intensive şi superintensive.
Intensivizarea trebuie văzută nu numai în ceea ce priveşte numărul de pomi la unitatea
de suprafaţă, ci în toate verigile tehnologice: scurtarea timpului de producere a materialului
săditor, utilizarea de combinaţii soi/portaltoi care se pretează la densităţi mari, amplasarea
livezilor în condiţii pedoclimatice care satisfac cerinţele, asigurarea la nivel optim a
îngrăşămintelor chimice şi a apei, adoptarea unor forme de coroană care să utilizeze cu
eficienţă maximă energia solară.
Tabelul 1.6
Dinamica producţiei de mere în România
mii tone
ANUL PRODUCŢIA ANUL PRODUCŢIA
1938
1950
1960
1970
1980
1985
1986
1987
1988
1989
1990
1991
1992
1993
1994
566,5
76,4
111,1
175,8
391,5
699,1
994,4
594,2
609,4
697,4
683,2
504,9
541,1
1097,2
363,0
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
2008
2009
457,2
659,7
664,0
610,0
316,1
365,0
507,0
491,5
811,1
1097,8
637,9
599,4
475,4
459,0
517,5
Sursa: FAO STAT (http://apps.fao.org) şi M.A.A.P. - Date operative
16. 14
În Europa de Est, producţia de mere a stagnat sau a avut creşteri uşoare, datorită
îmbătrânirii, neîngrijirii corespunzătoare a livezilor şi moderatei răspândiri a livezilor
intensive. Această tendinţă este pronunţată mai ales în statele fostei URSS, Ungaria, România
şi Bulgaria. Un salt cantitativ şi calitativ s-a înregistrat în Polonia, Republica Cehă, Slovacia
şi statele ce formau Iugoslavia. Guvernele ţărilor din Europa de Est nu-şi pot permite să
sprijine financiar modernizarea şi intensivizarea pomiculturii, iar dobânzile mari la credite
împiedica înfiinţarea de noi plantaţii pomicole.
În viitor mărul va rămâne specia fructiferă cu importanţă majoră în Europa de Est, care
va participa cu 50-80% din producţia totală de fructe. Mai ales, în anii ‘80 au fost înfiinţate
livezi intensive, însă se estimează că rata de intensivizare a plantaţiilor de măr din ţările est-
europene este de 20-40%. Este foarte greu de preconizat când ţările din această regiune vor
ajunge la nivelul de intensivizare al ţărilor din Europa de Vest (Papp J.,1996).
17. 15
Capitolul 2
SPECIILE POMICOLE DE CLIMAT TEMPERAT.
TAXONOMIE, NOMENCLATURĂ, CLASIFICARE
2.1. TAXONOMIE
Toate speciile fructifere de climat temperat aparţin încrengăturii Angiospermae. La
momentul apariţiei omului existau numeroase specii angiosperme spontane, care au constituit
una din principalele surse de hrană.
Speciile pomicole de climat temperat care intră în alimentaţia omului fac parte din 10
ordine de angiosperme: Rosales, Ericales, Eleagnales, Rhamnales, Cornales, Juglandales,
Fagales, Urticales, Rubiales, Actinidiales.
Ordinele Rosales şi Fagales sunt reprezentate prin două familii fiecare: Rosaceae şi
Saxifragaceae, respectiv Fagaceae şi Betulaceae, iar celelalte ordine participă cu câte o
singură familie.
Ţinând cont de producţia de fructe din zona de climat temperat, ponderea cea mai
mare o ocupă familia Rosaceae. Datorită numărului mare de specii, precum şi diversităţii lor,
rosaceele sunt grupate pe subfamilii. Trei din aceste subfamilii: Pomoideae, Prunoideae şi
Rosoideae furnizează 2/3 dintre principalele specii fructifere (11 din cele 17 specii mai
importante de climat temperat), în timp ce celelalte familii sunt reprezentate în livezile din
ţara noastră prin 1-4 specii (tabelul 2.1).
Speciile fructifere de climat temperat sunt cuprinse în 12 familii care cuprind şi specii
spontane, rezultate în urma evoluţiei vegetalelor, datorită îndelungatului proces de selecţie
naturală. Preponderentă este familia Rosaceae care cuprinde 12 genuri, alcătuite fiecare din
numeroase specii (tabelul 2.1).
Alegerea din flora spontană şi înmulţirea formelor cu fructe mai mari şi mai gustoase,
în acţiunea de selecţie empirică începută cu multe milenii în urmă a condus la obţinere
formelor domestice (cultivate), grupate în prezent sub denumirea de soiuri. După literatura de
specialitate, numărul soiurilor existente este extrem de mare. La măr - Malus domestica
Borkh, se cunosc peste 10000 de soiuri; părul - Pirus sativa Lam et. D.C. cuprinde peste 6000
soiuri; prunul - Prunus domestica L., peste 2000 soiuri, etc. Prin lucrările de selecţie şi
ameliorare efectuate în timp şi care continuă şi în prezent în toate ţările cultivatoare, numărul
soiurilor este mai mare şi continuă să crească (Miliţiu I, V. Cireaşă, 1992).
Pe lângă soiurile cultivate, mai sunt cuprinse şi specii spontane, utilizate în
pomicultură, fie ca portaltoi, fie ca genitori în activitatea de creare a noilor soiuri.
2.2. CLASIFICAREA SPECIILOR POMICOLE
2.2.1. Clasificarea după habitus
Ţinând cont de habitusul lor (aspectul general) speciile pomicole pot fi grupate în:
pomi propriu-zişi, arbustoizi, arbuşti, subarbuşti şi semierboase perene.
Pomii propriu-zişi. Sunt plante lemnoase de vigoare variabilă, cu un singur trunchi şi
coroană omogenă. Pomii pot atinge înălţimea de 5 - 20 m şi au o longevitate mare, de la 15
până la 100 de ani (chiar mai mult la castan). Se pot înmulţi prin seminţe (speciile sălbatice,
portaltoi franc), prin altoire şi mai rar prin drajoni. În această grupă sunt cuprinse următoarele
specii: mărul, părul, cireşul, vişinul, nucul, caisul, piersicul, prunul, castanul.
18. 16
Tabelul 2.1
Unităţile sistematice în care sunt cuprinse speciile pomicole
(după Miliţiu I, V. Cireaşă, 1992)
Familia Subfamilia Genul Specia Denumirea populară
0 1 2 3 4
Rosaceae
Pomoideae
Malus
- domestica Borkh.
- silvestris Mill.
- pumilla praecox Pall.
- pumilla paradisiaca Pall.
- baccata Borkh.
- prunifolia Borkh.
- măr cultivat
- măr pădureţ
- măr dusen
- măr paradis
- măr siberian
- măr chinezesc
Pirus
- sativa Lam. Et DC
- piraster L.
- eleagrifolia Pall.
- nivalis Jac.
- serotina Red.
- păr cultivat
- păr pădureţ
- păr sălcioară
- păr nins
- păr chinezesc
Cydonia
- oblonga Mill.
- japonica Pers.
- gutui cultivat
- gutui japonez
Sorbus - domestica L.
- aucuparia L.
- melanocarpa Rehd.
- scoruş
- scoruş de munte
- scoruş negru
Mespilus - germanica L. - moşmon
Prunoideae
Prunus
- domestica l.
- insititia jusse.
- cerasifera mhr.
- spinosa l.
- prun cultivat
- prun goldan
- corcoduş
- porumbar
Armeniaca
- vulgaris Lam
- sibirica L.
- mume Carr.
- mandchurica Skhvortz.
- cais
- cais siberian
- cais japonez
- cais Manciuria
Persica
- vulgaris Mill.
- davidiana Kaar.
- piersicul
- piersicul lui David
Amygdalus
- communis L.
- nana L.
- migdalul
- migdalul pitic
Cerasus
- avium L.
- vulgaris Mill.
- mahaleb Mill.
- fruticosa Pall.
- cireşul
- vişinul
- mahaleb
- vişin de stepă
Rosoideae
Fragaria
- vesca L.
- grandiflora Ehrh.
- virginiana Mill.
- chiloensis Duchean.
- fragul de pădure
- căpşun
- fragul de Virginia
- fragul de Chile
Rubus
- idaeus var. vulgatus L.
- idaeus var. strigosus Mill.
- caesius L.
- fruticosus L.
- zmeur roşu
- zmeur pufos american
- mur de câmp
- mur de pădure
Saxifragaceae Ribesoideae
Ribes
- vulgaris Lam.
- rubrum L.
- nigrum L.
- aureum Pursh.
- coacăz sălbatic
- coacăz roşu
- coacăz negru
- coacăz auriu
Grossularia - reclinata Mill. - agriş
Ericaceae Vaccinium
- vitis-idaea L.
- uliginosus L.
- myrtillus L
- corymbosum şi angustifolium
- afinul roşu
- afinul vânăt
- afinul negru
- afinul cu tufă înaltă
Eleagnaceae Hippohae - rhamnoides L. - cătina albă
Rhamnaceae Ziziphus - jujuba Mill. - gigif
Cornaceae Cornus - mas L. - cornul
Juglandaceae Juglans
- regia L.
- nigra L.
- nucul comun
- nucul negru
Fagaceae Castanea
- sativa Mill.
- dentata Borkh.
- pumilla Mill.
- castanul
- castan american
- castan pitic
Betulaceae Corylus
- avellana L.
- colurna L.
- maxima Mill.
- pontica Koh.
- alunul comun
- alunul turcesc
- funducul
- alunul pontic
Moraceae
Morus - alba L. - dudul
Ficus - carica L. - smochinul
Caprifoliaceae Sambucus - nigra L. - socul european
Actinidiaceae Actinidiae Actinidia
- chinensis Planch.
- deliciosa Achev.
- kiwi
- actinidia chinezească
19. 17
Arbustoizii. Sunt plante lemnoase de vigoare redusă, cu 2-4 tulpini de grosimi şi
înălţimi variabile (3-7 m), sub formă de tufă, care iau naştere din zona coletului, formând
coroane separate sau unice. Longevitatea este de cca. 20-30 ani şi se înmulţesc mai ales pe
cale vegetativă, prin drajoni, butaşi şi marcote, şi mai rar prin seminţe.
Principalele specii care fac parte din această grupă sunt: gutuiul, alunul, cornul,
moşmonul, vişinul arbustoid, mărul paradis, scoruşul, cătina albă.
Arbuştii. Sunt plante lemnoase de vigoare mică, care prezintă numeroase tulpini de
grosimi mici, slab ramificate, ce apar din zona coletului, formând tufe compacte, cu înălţimi
de 1-1,5 m. Durata vieţii este redusă (10-15 ani).
Având în vedere particularitatea de a emite uşor rădăcini adventive, se înmulţesc
vegetativ prin drajoni, butaşi şi marcote, dar se pot înmulţi şi prin seminţe. Dintre arbuştii
fructiferi amintim: coacăzul, agrişul, socul, scoruşul negru.
Semiarbuştii. Sunt plante semilemnoase, formate din tulpini care trăiesc numai 2 ani.
În primul an cresc tulpinile şi diferenţiază mugurii de rod, înalte de 1-3 m, iar în al 2-lea an
rodesc. După recoltarea fructelor tulpinile se usucă, însă plantele se regenerează prin drajoni,
ceea ce face ca durata de viaţă să ajungă la 10-20 ani. Această grupă este reprezentată prin:
zmeur şi mur.
Plantele fructifere semierboase. Sunt cele care fac trecerea dintre plantele
arboreşcente şi cele ierboase, fiind reprezentate prin căpşun şi frag. Căpşunul creşte sub formă
tufă cu înălţimea de 15-30 cm. Parte subterană este formată dintr-o tulpină scurtă (rizom) şi
rădăcini, iar partea aeriană este reprezentată de lăstari foarte scurţi (2-3 cm.), cu frunze lung
peţiolate, inflorescenţe şi stoloni (tulpini târâtoare). Frunzele se usucă anual, însă tulpinile
trăiesc 6-8 ani, exploatarea economică fiind de 3-4 ani.
2.2.2. Clasificarea pomicolă
În funcţie de particularităţile biologice şi tehnologice speciile pomicole de climat
temperat se împart în următoarele grupe:
• Pomacee. Cuprind speciile din familia Rosaceae, subfam. Pomoideae: măr, păr, gutui,
moşmon, scoruş, etc., şi au următoarele caracteristici:
sunt bine adaptate climatului temperat;
datorită cerinţelor mari faţă de umiditate şi fertilitatea solului, reuşesc mai ales în zona
dealurilor din ţara noastră; soiurile timpurii au cerinţe mai scăzute faţă de apă şi pot fi
cultivate şi în zone mai secetoase;
majoritatea speciilor din această grupă sunt rezistente la ger; repausul obligatoriu este
lung şi înfloresc târziu primăvara, astfel încât, de regulă, scapă de brumele târzii de
primăvară;
durata de viaţă în cultură este de 50-70 ani în livezile clasice şi de 15-20 de ani în
livezile intensive şi superintensive, unde datorită utilizării unor combinaţii soi-portaltoi
specifice, cu intrare rapidă pe rod, dau recolte economice încă din anul 2-3 de la plantare (mai
ales la măr);
mugurii floriferi sunt micşti, situaţi terminal, iar lateral şi spre baza lor există rezerve
de muguri vegetativi, ceea ce permite regenerarea ramurilor îmbătrânite şi epuizate;
fructele sunt “false”, la formarea lor participă nu numai ovarul, ci şi receptaculul florii,
şi poartă denumirea de poamă;
degarnisirea ramurilor se face într-un ritm lent, suportă tăierile şi îşi vindecă rănile
relativ uşor;
majoritatea soiurilor sunt autosterile;
20. 18
datorită existenţei unei game foarte largi de portaltoi (mai ales la măr), permit
cultivarea în diferite sisteme de cultură, de la clasic până la intensiv şi superintensiv.
• Drupacee. Această grupă include: prunul, caisul, piersicul, cireşul, vişinul, migdalul, care
aparţin familiei Rosaceae, subfamiliei Prunoideae şi se caracterizează prin:
sunt specii mai puţin rezistente la ger decât pomaceele; repausul este mai scurt,
înfloresc primăvara devreme motiv pentru care brumele şi îngheţurile târzii de primăvară pot
produce frecvent pagube (degerarea florilor sau a fructelor proaspăt legate). În consecinţă
cultura drupaceelor este cantonată în zonele mai calde ale ţării, pe coline şi la câmpie, cu
excepţia unor soiuri de prun , cireş şi vişin care merg în zona dealurilor;
sunt mai pretenţioase faţă de căldură, au cerinţe reduse faţă de umiditate şi sunt relativ
rezistente la secetă (cais, migdal);
durata medie de viaţă a pomilor este mai scurtă la cais şi piersic (15-20 de ani) şi mai
lungă la prun şi cireş (până la 40-50 de ani);
au o creştere viguroasă şi rapidă, formează mulţi lăstari anticipaţi (mai ales caisul şi
piersicul);
se degarnisesc foarte repede deoarece, de regulă, ramurile de rod la aceste specii au
mugurii floriferi amplasaţi lateral, iar cei vegetativi sunt situaţi terminal, creşterea
deplasându-se către exteriorul coroanei;
soiurile de drupacee sunt numeroase, dar mai puţine comparativ cu pomaceele, şi
multe dintre acestea sunt autofertile;
numărul portaltoilor folosiţi este mai redus decât la pomacee.
• Nuciferele. Cuprind speciile nuc, castan, alun şi prezintă următoarele caracteristici:
reprezintă speciile pomicole cu fructe uscate (alună şi castană), cu excepţia nucului
(drupă falsă);
în general, sunt mai puţin rezistente la geruri, pretenţioase faţă de căldură (castanul),
unele şi faţă de lumină (nucul), motiv pentru care sunt răspândite în zone cu climă mai blândă
(mai ales castanul); alunul este mai puţin pretenţios;
datorită polenizării anemofile şi a înmulţirii prin seminţe există numeroase biotipuri şi
populaţii locale (mai ales la nuc şi castan); se practică şi înmulţirea prin altoire (mai ales la
nuc) şi prin drajoni, butaşi, marcotaj (la alun).
• Baciferele:
cuprind specii din familii diferite:
- Saxifragaceae - coacăzul şi agrişul;
- Rosaceae - zmeurul, murul şi căpşunul;
- Ericaceae - afinul;
din punct de vedere morfologic fructele acestor specii sunt foarte diferite: bacă la
coacăz şi afin, pseudobacă la agriş, zmeurul şi murul au fructul polidrupă, iar căpşunul
poliachenă;
toate aceste specii au fructul foarte perisabil;
unele specii sunt lemnoase (coacăz, agriş), altele semilemnoase (zmeur şi mur), iar
căpşunul face trecerea între plantele lemnoase şi cele ierboase;
au talie mică şi formează tufe dese;
trăiesc 10-15 ani, cu excepţia căpşunului (6-8 ani);
se înmulţesc uşor pe cale vegetativă, prin drajoni, stoloni sau marcote.
• Grupa speciilor subtropicale. Aceasta grupă include specii din climatul subtropical
umed, sensibile la ger, puţin răspândite la noi. Unele se cultivă în câmp (smochinul, kiwi,
kaki), iar altele în apartamente (lămâiul, mandarinul, portocalul, grapefruitul).
21. 19
Capitolul 3
MORFOLOGIA ŞI FIZIOLOGIA POMILOR ŞI
ARBUŞTILOR FRUCTIFERI
În general, speciile pomicole sunt plante complexe obţinute prin altoirea a două sau
chiar trei porţiuni de plante diferite, care convieţuiesc formând o asociaţie, o simbioză. În
cadrul acestor asociaţii unul dintre parteneri - hipobiontul (portaltoiul) absoarbe din sol apa şi
substanţele nutritive şi le transmite epibiontului (altoiul) care, la rândul lui, prin fotosinteză, le
transformă în substanţe elaborate necesare pentru creşterea ambilor parteneri, precum şi
pentru obţinerea recoltei.
Simbionţii îndeplinesc o serie de funcţii comune (depozitarea, respiraţia, transpiraţia,
etc.), dar şi funcţii specifice fiecărui partener, cum ar fi absorbţia apei şi substanţelor nutritive
pentru rădăcină şi fotosinteza pentru epibiont.
Între partea hipogee şi cea epigee a pomilor există un echilibru, caracteristic pentru
fiecare specie şi asociaţie soi-portaltoi. Modificările produse asupra uneia dintre din cele 2
categorii de organe produce schimbări şi la celelalte. Astfel, tăierea unei părţi din ramurile
coroanei are ca efect realizarea unui volum al sistemului radicular mai redus decât la pomii
cărora nu li s-au aplicat tăieri. De asemenea, reducerea sistemului radicular, prin executarea
lucrărilor de întreţinere a solului, conduce la creşteri mai reduse ale pomului (Miliţiu I.,
1992).
În afară de indivizii altoiţi, mai există şi plante pomicole pe rădăcini proprii obţinute
pe cale generativă (prin seminţe) sau prin înmulţire vegetativă (marcote, butaşi, drajoni, etc.).
3.1. ORGANELE HIPOGEE
Speciile pomicole sunt alcătuite, ca toate plantele, din rădăcină (partea subterană) şi
tulpină (partea aeriană), însă principala deosebire constă în faptul că majoritatea pomilor se
înmulţesc prin altoire şi, în consecinţă, rădăcina aparţine altui soi sau cel mai des altei specii.
Organele vegetative situate sub nivelul solului alcătuiesc rădăcina (partea hipogee).
Clasificarea rădăcinilor
La pomii fructiferi, rădăcinile sunt clasificate după următoarele criterii:
A– După origine:
a) Rădăcini embrionare – se formează în urma germinării seminţei, din radicula
embrionului. Aceste rădăcini se întâlnesc la pomii obţinuţi din seminţe (nuc, tipuri locale de
prun, etc.) şi la portaltoii franc de : măr, păr, prun, piersic, cireş, vişin, mahaleb, zarzăr şi
corcoduş.
b) Rădăcini adventive – se formează pe nodurile şi internodurile butaşului, marcotei
sau drajonului.
B– După funcţiile îndeplinite:
a) Rădăcini de schelet şi semischelet, lungi de 2-3 m – 7-8 m şi cu grosimi de 2-3 mm
până la 20-30 cm. Scheletul şi semischeletul rădăcinii este alcătuit din rădăcina principală sau
pivotul şi rădăcinile secundare, de ordinul I şi II. Rădăcinile de schelet şi semischelet cresc
vertical, oblic şi orizontal şi îndeplinesc funcţii de fixare a pomului în sol, de conducere a
sevei şi de depozitare a substanţelor de rezervă.
b) Rădăcinile de garnisire (fibroase) sunt rădăcini de ordin superior (începând cu
ordinul IV) cu lungimea şi grosimea de câţiva mm; la această grupă se încadrează următoarele
tipuri de rădăcini:
b1) Rădăcini axiale (de creştere) au structură primară, se formează primăvara în
vârful rădăcinilor de schelet ale pomilor tineri, sunt albe, transparente, lungi de 10 – 25 mm.
22. 20
b2) Rădăcinile active sau absorbante sunt albe translucid, lungi de 0,1 – 4 mm şi
groase de 0,3 – 1 mm şi cu o durată de viaţă scurtă. Au ca funcţie principală absorbţia apei şi
a substanţelor nutritive din sol.
b3) Rădăcinile intermediare sunt albe-violacee, situate între rădăcinile axiale şi
conducătoare şi între absorbante şi conducătoare. Ele se transformă în rădăcini conducătoare
(dacă provin din rădăcini axiale), sau se usucă şi pier (dacă provin din rădăcini active).
b4) Rădăcini conducătoare, în devenire – cu grosimi de 2,5-3 mm situate în
vârful rădăcinilor conducătoare – propriu-zise.
C– După durata de viaţă:
a) Rădăcini permanente, care se dezvoltă în sol simultan cu ramurile principale ale
coroanei şi formează structura de bază a sistemului radicular (rădăcinile de schelet şi de
semischelet). Ele au o durată lungă de viaţă, echivalentă cu vârsta componentelor coroanei, cu
care se află într-o conexiune permanentă.
b) Rădăcini nepermanente (fibroase) sunt reprezentate prin: rădăcini axiale, active,
intermediare şi conducătoare. Ele asigură extensia sistemului radicular, absorbţia şi
transportul elementelor nutritive spre şi de la rădăcinile permanente.
D. – După unghiul pe care îl formează rădăcinile de schelet cu verticala (unghiul
geotropic) rădăcinile pot fi:
a) Cu creştere verticală (rădăcini pivotante)– unghiul geotropic este cuprins între 0o
şi
30o
(nuc, cireş, păr);
b) Cu creştere oblică – unghiul geotropic este cuprins între 30-60o
(cais, vişin, prun,
portaltoii generativi ai mărului);
c) Cu creştere orizontală – unghiul geotropic este cuprins între 60-90o
, deci rădăcinile
cresc aproape paralel cu orizontala solului. Aceste rădăcini sunt numite şi rădăcini trasante.
Sunt întâlnite la gutui, portaltoii de măr de vigoare slabă.
Creşterea în lungime a rădăcinilor se datorează prezenţei meristemului apical, iar
formarea rădăcinilor laterale se realizează în urma diferenţierii celulelor periciclului, care se
divid formând un con de creştere, care pătrunde printre celule, până la suprafaţa rădăcinii
(Burzo I. şi colab, 2005).
Creştere a rădăcinilor este influenţată de specie, soi, însuşirile fizico-chimice ale
solului, agrotehnica din livadă, etc.
Pomii şi arbuştii fructiferi care se înmulţesc prin sămânţă, au o rădăcină principală
(pivot), care provine din radicula embrionului. Plantele înmulţire prin butaşi, au rădăcini
adventive situate mai ales la nivelul nodurilor şi cu tendinţă de creştere orizontală. Gradul de
ramificare a rădăcinii este dependent de condiţiile existente în sol şi de gradul de fertilitate al
acestuia, o bună ramificare realizând-se în solurile uşoare, profunde şi bine aerate. Lungimea
rădăcinilor este însă mai mare în solurile sărace, comparativ ce cele fertile.
Durata de viaţă a perilor absorbanţi variază de la câteva săptămâni, la câteva luni.
Black citat de Burzo I. şi colab, 2005, a constatat că mai puţin de 60% din rădăcinile de cireş
supravieţuiesc mai mult de 14 zile. Rădăcinile absorbante reprezintă cea mai mare parte a
sistemului radicular. Astfel, Negrilă A., 1982, arată că, numărul rădăcinilor active reprezintă
circa 90% din numărul total al rădăcinilor, iar după Baldini E., 1986, partea absorbantă a
rădăcinii pomilor reprezintă 80-85% din sistemul radicular.
Distribuţia rădăcinilor în sol este importantă pentru efectuarea corectă a lucrărilor de
întreţinere şi fertilizare. Adâncimea la care se găsesc majoritatea rădăcinilor este, în general,
de 20-80 cm., iar pe solurile înierbate, acestea se situează ceva mai sus (10-40 cm.).
Adâncimea maximă la care pot ajunge unele rădăcini cu poziţie verticală, depinde de specie şi
portaltoi, putând fi de până la 8-10 m, în special pe solurile mai uscate şi nisipoase. Însuşirile
fizice şi chimice ale solului, fertilitatea acestuia, precum şi portaltoiul, influenţează distribuţia
23. 21
sistemului radicular; solurile uşoare nisipoase determină o mai mare creştere şi răspândire a
rădăcinilor , în timp ce în solurile fertile sistemul radicular este mai puţin extins.
Extinderea în plan orizontal a rădăcinilor este influenţată de combinaţia soi-portaltoi,
proprietăţile fizico-chimice şi modul de întreţinere a solului. În general, majoritatea
rădăcinilor active se găsesc sub proiecţia coanei (0,6-1,8 m de la trunchi şi până la periferia
coroanei). Aceasta este zona în care trebuie să se administreze îngrăşămintele chimice şi
organice pentru a putea fi valorificate rapid şi eficient.
Longevitatea şi ritmul anual de creştere al rădăcinilor. În decursul vieţii rădăcinile
unui pom se modifică foarte mult: an de an apar rădăcini noi, care se îndreaptă către zonele
unde există îngrăşăminte şi apă, iar altele mor datorită excesului accidental de apă, secetei
prelungite sau sunt tăiate cu ocazia lucrărilor solului.
Sezonul de activitate al rădăcinilor este mult mai lung decât al organelor epigee.
Rădăcinile îşi încep activitatea mult mai devreme primăvara când în sol se înregistrează cca.
20
C şi o încheie mult mai târziu toamna. Există păreri că, de exemplu la măr, creşterea
rădăcinilor are loc tot anul. Această creştere îndelungată a rădăcinilor are, în general, 1 - 3
valuri. Astfel, creşterea rădăcinilor atinge un maximum înainte de creşterea intensă a
lăstarilor, un al doilea maxim înregistrându-se după ce lăstarii şi-au oprit creşterea. La pomii
tineri raza volumului de sol ocupat de rădăcini creşte, în medie, cu 0,5 m anual (Miliţiu I.,
1992).
Temperaturile sub 00
C şi mai mari de 300
C reduc ritmul de creştere a rădăcinilor.
Acest proces este iniţiat la temperatura de 10
C la căpşun, 4-60
C la măr, 6-70
C la păr şi 120
C la
cais şi piersic.
Durata şi ritmul de creştere anul al rădăcinilor sunt influenţate de foarte mulţi factori,
în primul rând de către portaltoi, condiţiile de mediu (mai ales solul) şi tehnologiile aplicate
(mai ales fertilizarea, irigarea, tăierile). Astfel, pe solurile pietroase sistemul radicular al
pomilor depăşeşte uneori de 3-4 ori proiecţia coroanei, iar pe un sol lutos, de peste 5 ori. La
portaltoii generativi, sistemul radicular este de 4-6 ori mai mare decât la cei vegetativi
(Cepoiu N., 2001).
Tăierile severe ale coroanei, stimulează creşterea părţii aeriene în detrimentul
rădăcinilor a căror creştere este inhibată. Fenomenul este determinat de corelaţia trofică
pozitivă care există între rădăcinile şi organele aeriene ale plantelor pomicole.
Încărcătura mare de fructe determină transportul prioritar al asimilatelor spre fructe şi
seminţe, în detrimentul creşterii rădăcinilor. Prioritatea în transportul substanţelor organice se
face în următoarea ordine: fructe, muguri, rădăcini, organe de rezervă rădăcinii (Burzo I. şi
colab, 2005).
Valoarea optimă a pH-ului solului, pentru creşterea sistemului radicular la speciile
perene, variază între 5,7 şi 8,3, cu excepţia afinului care preferă solurile mai acide, cu pH-ul
de 4,5-5,5 şi a migdalului care preferă pH-ul alacalin.
Înierbarea permite creşterea rădăcinilor pomilor în stratul superficial al solului, dar
apare concurenţa între rădăcinile acestora şi ale ierburilor, în ceea ce priveşte apa şi
substanţele minerale, crescând sensibilitatea la stresul hidric în cazul livezilor cu intervalul
între rânduri înierbat.
În mod evident, irigarea stimulează creşterea rădăcinilor. Astfel, numeroşi autori
consideră că, numărul rădăcinilor care se formează în solul irigat este de 5-7 ori mai mare,
comparativ cu martorul neirigat în funcţie de specie, soi şi portaltoi. Excesul de umiditate are
efect negativ, determinând asfixia rădăcinilor. Potenţialul hidric al solului trebui să varieze
între -0,01 şi - 0,5 MPa (Chiţu E., 2002) pentru a putea asigura o bună aprovizionare cu apă a
plantelor.
24. 22
Seceta are un efect stresant asupra rădăcinii pomilor şi a arbuştilor fructiferi,
determinând inhibarea creşterii rădăcinilor, moartea perilor absorbanţi şi sinteza acidului
abscizic, care transportat în frunze, determină închiderea stomatelor.
Nutriţia minerală reprezintă un factor important ce influenţează creşterea rădăcinii
pomilor şi a arbuştilor fructiferi. Nutriţia corespunzătoare cu azot favorizează creşterea în
lungime a rădăcinilor, iar fosforul stimulează ramificarea şi creşterea masei acestora
(Kolesnikov, citat de Burzo I. şi colab, 2005).
Funcţiile organelor hipogee:
Ancorarea în sol. Depinde în primul rând de particularităţile genetice ale portaltoiului,
dar şi de textura şi adâncimea solului. Portaltoii viguroşi au o mai bună ancorare în sol decât
cei de vigoare redusă care necesită sistem de susţinere.
Absorbţia apei şi a substanţelor nutritive. Odată cu apa sunt absorbiţi din sol ionii
elementelor nutritive, dar şi substanţe organice cu moleculă mică existente în soluţia solului.
Absorbţia se efectuează îndeosebi prin intermediul perişorilor absorbanţi. În literatura de
specialitate exista păreri conform cărora, apa poate pătrunde în rădăcini şi prin intermediul
scoarţei, care este mult mai permeabilă pentru apă decât coaja tulpinii (Miliţiu I., 1992).
Transportul apei şi substanţelor nutritive. Transportul substanţelor minerale
absorbite de către frunze se face prin xilem, care este continuu de la vârful rădăcinii până la
frunze. Paralel cu acest curent ascendent are loc transportul sevei elaborate, de la frunze către
vârfurile de creştere ale rădăcinii sau către locurile de depozitare, prin intermediul floemului.
Respiraţia. La majoritatea speciilor pomicole se manifestă nevoi mari faţă de oxigenul
din sol (se consideră condiţii optime când O2 reprezintă 10% din aerul din sol). Rădăcinile
tinere sunt mai pretenţioase faţă de O2 decât cele în vârstă, iar rădăcinile absorbante sunt
foarte sensibile la lipsa de O2.
Depozitarea. Rădăcinile reprezintă un important organ de depozitare a substanţelor de
rezervă. Cele cu structura primară depozitează substanţe nutritive sub formă de amidon în
cortex, iar la cele care apar îngroşări secundare depozitarea se face în celulele parenchimatice
şi de sclerenchim ale xilemului şi floemului (Miliţiu I., 1992).
Substanţele depozitate în rădăcini pot fi mobilizate şi utilizate în diferite momente:
creşterea rădăcinilor primăvara înainte de intrarea în vegetaţie, vindecarea rănilor apărute cu
ocazia plantărilor sau arăturilor, etc.
Sinteza primară a unor compuşi organici. La nivelul rădăcinilor (mai ales a celor
absorbante) s-a constatat transformarea azotului anorganic absorbit din sol în substanţe
organice (amide, aminoacizi), utilizând energia rezultată din descompunerea hidraţilor de
carbon.
Sinteza substanţelor bioactive. Biosinteza şi transportul hormonilor vegetali
(auxinele, giberinele, citokininele, acidul abscizic şi etilenul) reprezintă una dintre cele mai
importante funcţii ale sistemului radicular.
Creşterea rădăcinilor este influenţată de prezenţa substanţelor hormonale. Rădăcinile
sunt sediul principal al biosintezei acidului abscisic, al citochininelor şi al etilenei, alături de
care se pot biosintetiza şi gibereline. Auxina provine însă din organele aeriene ale plantelor.
Acidul abscisic are un efect de inhibare a creşterii rădăcinilor, iar etilena stimulează
procesul de îngroşare a pereţilor celulari. Rolul giberelinelor, citochininelor şi a auxinei în
procesul de creştere a sistemului radicular, nu este bine precizat. Se consideră că,
citochininele stimulează diviziunea celulelor din meristemul apical al rădăcinilor, dar în
cantitate mai mare acest hormon vegetal poate inhiba acest proces. Auxina are un rol
stimulator asupra procesului de rizogeneză (Burzo I. şi colab, 2005).
Auxina (acidul indolilacetic). Sinteza auxinei se pare a fi mai degrabă în ţesuturile mai
mature ale rădăcinilor decât în vârfuri. Aceasta este transportată acropetal, de la baza
rădăcinilor către vârful lor, prin intermediul sistemului vascular sau prin celule vii ale
25. 23
cilindrului central (Miliţiu I., 1992). În afară de rădăcină, auxina naturală (acid 3-
indolilacetic) mai este produsă în regiunile subapicale ale lăstarilor cu creştere activă, în
frunzele tinere, în embrionii în curs de dezvoltare (Westwood M.N., 1978, Dorobanţu N.,
1976, Milică C. I. şi colab., 1977).
Auxina controlează formarea şi creşterea rădăcinilor, fiind inactivă în concentraţii
foarte mici şi inhibitoare a acestor procese când se acumulează în cantităţi foarte mari
(Negrilă A., 1982).
În practica pomicolă auxinele sintetice AIB (acid indolil-3-butiric), ANA (acidul
alfanaftilacetic), acidul 2,4 D se utilizează pentru a stimula înrădăcinarea butaşilor.
Giberelina. Vârful rădăcinilor este locul de conversie a giberelinelor sintetizate în
frunzele tinere către o formă mai activă. Nu se cunoaşte complet rolul giberelinei în creşterea
rădăcinilor, însă se pare că acestea stimulează creşterea, mai exact elongaţia celulară şi
iniţierea rădăcinilor laterale (Negrilă A., 1982, Miliţiu I., 1992).
În general, giberelina stimulează creşterea lăstarilor şi în consecinţă a suprafeţei
foliare, formarea anticipaţilor, formarea partenocarpică a fructelor, întârzie colorarea şi
maturarea fructelor (Negrilă A., 1982).
Citokinina. Este produsă în meristemul apical al rădăcinii. Rolul acesteia depinde de
interacţiunea cu alţi hormoni, în special cei care influenţează diviziunea celulară (Miliţiu I.,
1992).
Se pare că, citokinina inhibă creşterea rădăcinilor, având un rol important în formarea
mugurilor adventivi de pe rădăcini şi tulpini (Negrilă A., 1982). Transportul citokininelor
sintetizate în rădăcină se face ascendent prin xilem şi stimulează sinteza proteinelor şi a
enzimelor din frunze, formarea lăstarilor laterali, ieşirea mugurilor din repaus, legarea
fructelor şi previne îmbătrânirea florilor, frunzelor şi fructelor. În seva xilemică a pomilor
deplasarea citokininelor este maximă primăvara, aproximativ în faza înfloririi depline, nivelul
este foarte scăzut spre sfârşitul verii şi rămâne minim pe întreaga perioadă a iernii. Această
schema sezonieră concordă cu rolul citokininelor în legarea fructelor, creşterea frunzelor şi
controlul îmbătrânirii (Westwood M.N., 1978).
Acidul abscizic. Este un inhibitor natural de creştere care provoacă repausul mugurilor
şi seminţelor, inhibă creşterea rădăcinilor şi lăstarilor, absorbţia şi acumularea ionilor de P şi
K, stimulează îmbătrânirea şi căderea frunzelor şi fructelor (Negrilă A., 1982, Miliţiu I.,
1992).
Locul primar de sinteză al acidului abscizic este în frunzele mature când zilele devin
din ce în ce mai scurte, la sfârşitul verii, dar şi rădăcinile generează acest hormon, mai ales în
condiţii de stres fizic şi hidric (Westwood M.N., 1978).
Etilenul. Nu este în sens strict un hormon vegetal, dar formarea lui în pom are aspecte
specifice de reglare, ca: scoaterea din repaus a mugurilor şi seminţelor, accelerează coacerea
fructelor şi formarea culorii, stimulează inducţia florală, facilitează desprinderea fructelor şi
frunzelor (Negrilă A., 1982, Miliţiu I., 1992).
Rădăcinile sunt puternic inhibate în ceea ce priveşte creşterea în lungime de către
etilen. Efectul acestuia este legat de raportul auxină - etilen; în mod normal conţinutul de
etilen este sub pragul de inhibare a creşterii, însă prezenţa auxinei la un nivel ridicat provoacă
creşterea etilenului peste pragul de inhibare a creşterii.
Etilenul este generat de pom mai ales în condiţii de stres fizic, îndeosebi în zonele
rănite. S-a constatat că etilenul stimulează mai degrabă extinderea laterală a rădăcinilor decât
iniţierea lor, datorită unei auxine indusă de etilen (Miliţiu I., 1992).
Secreţia (excreţie sau desorbţie) constă în eliminarea din rădăcini în mediul exterior a
unor substanţe care au rol în solubilizarea sărurilor complexe din sol, dezvoltarea rizosferei;
iar altele au efect toxic, conduc la aleopatie.
26. 24
În imediata vecinătate a rădăcinilor, în zona perişorilor absorbanţi (denumită
rizosferă), se creează condiţii pentru dezvoltarea unui număr mare de microorganisme:
bacterii, ascomicete, ciuperci.
Exudatele rădăcinii favorizează relaţii de simbioză între flora solului şi rădăcini, cu
efecte benefice asupra participanţilor la asociaţie. Microorganismele oferă pomilor vitamine,
hormoni de creştere şi factori de rezistenţă la paraziţi şi atacă mineralele insolubile,
descompun humusul, sintetizează proteinele din azotul atmosferic.
La pomi se formează, în general, micoriză Prezenţa micorizei are ca efect sporirea
absorbţiei substanţelor nutritive, mai ales în solurile sărace în P, facilitează pătrunderea apei
în rădăcini şi sporeşte rezistenţa acestora la atacul microorganismelor dăunătoare.
Aleopatia este interacţiunea negativă dintre anumite substanţe lăsate de plantă în sol şi
creşterea rădăcinilor unei plante învecinate din aceiaşi specie sau din specii diferite. Prezenţa
în sol a substanţelor alopatice contribuie la apariţia fenomenului de “oboseală a solului”, care
se înregistrează în cazul replantării cu pomi a unor plantaţii recent defrişate.
Substanţele alelopatice au o compoziţie variată şi sunt reprezentate de: juglonă, acid β-
cetoadipic, taninuri, fenoli sau acizi fenolici, alcaloizi, purine, cumarine, lactone, terpene,
steroizi etc. Aceste substanţe ajung în soluţia solului si sub formă de secreţii ale rădăcinii, prin
biodegradarea frunzelor căzute pe suprafaţa solului, sau sunt eliminate sub forma de compuşi
volatili. Unele substanţe alelopatice acţionează direct, sub forma în care au fost eliminate, în
timp ce altele sunt transformate de bacterii sau ciuperci din sol în noi compuşi inhibitori de
creştere (ex. acidul β-cetoadipic). Efectele acţiunii substanţelor allelopatice asupra
metabolismului plantelor sunt foarte diverse. Unele substanţe ca: acizii cinamic, benzoic şi
cumarinele sunt inhibitori de creştere, în timp ce altele inhibă respiraţia, fosforilările
oxidative, fotosinteza, activitatea unor enzime, sinteza proteinelor, etc. (Burzo I. şi colab,
2005).
Este bine cunoscut cazul piersicului care elimină substanţe alelopatice şi, ca urmare
împiedică replantarea piersicului pe aceiaşi parcelă. Nucul produce juglona şi compuşi ce
conţin iod, care au un rol alelopatic, împiedicând creşterea unor plante în apropierea acestora.
Înmulţirea vegetativă La unele specii: prun, vişin, cătină, zmeur, mur, rădăcinile mai
au şi funcţia de înmulţire vegetativă. Pe rădăcinile acestor specii, se formează muguri
adventivi, care apoi evoluează în drajoni, iar odată cu detaşarea de planta mamă devin indivizi
noi.
3.2. ORGANELE EPIGEE
Partea aeriană a pomilor şi arbuştilor fructiferi este cunoscută sub numele de tulpină
(partea epigee).
Tulpina (fig. 3.1) reprezintă totalitatea ramificaţiilor unui pom cuprinse între colet şi
mugurii terminali ai ramurilor anuale periferice.
Creşterea în lungime a ramurilor se realizează prin diviziunea celulelor din meristemul
central al mugurilor terminali, iar prin diviziunea meristemelor apicale ale mugurilor axilari se
formează ramificaţiile laterale.
Creşterea în grosime a trunchiului şi a ramurilor se realizează prin diviziunea celulelor
cambiale, în urma căruia are loc şi procesul de formare al floemului şi xilemului. Diametrul
vaselor conducătoare variază în funcţie de condiţiile pedoclimatice ale fiecărui an, iar acestea
determină formarea inelelor anuale, care corespund cu vârsta pomului. La creşterea în
grosime participă şi feloderma, care acoperă la exterior tulpina.
Creşterea organelor aeriene ale plantelor este influenţată de substanţele hormonale.
Citochininele stimulează diviziunea celulelor din meristeme, iar auxina stimulează extensia
27. 25
celulară. Acidul abscizic are un efect contrar inhibând procesul de creştere (Burzo I. şi colab,
2005).
La pomii propriu-zişi, tulpina este formată dintr-un trunchi (de 30-100 cm) şi o
coroană, caracteristică speciei şi soiului. La arbustoizi, se întâlnesc 3-5 tulpini, cu sau fără
trunchi, care de regulă se dezvoltă asimetric şi dau un contur coroanei destul de neregulat. La
arbuşti, tulpina este o tufă deasă, cu înălţimea de 1-2 m. (Cepoiu, N., Monica Murvai, 1992).
Trunchiul – este partea cea mai de jos a tulpinii, încadrată între colet şi prima ramură
din coroană.
Coroana – este definită structural de axul coroanei (exceptând coroanele lipsite de ax),
ramurile de schelet şi semischelet şi ramurile de garnisire (vegetative şi de rod).
Axul coroanei se află situat în prelungirea trunchiului şi este delimitat la bază de
trunchi, iar la vârf de o creştere anuală numită săgeată.
Ramurile de schelet sunt cele mai viguroase ramuri din coroană. Ele sunt flexibile (la
pomii tineri) sau rigide (la pomii maturi) şi îndeplinesc funcţiile de susţinere a ramurilor de
semischelet şi de rod, a frunzelor şi fructelor, de conducere a sevei şi depozitare a substanţelor
de rezervă.
Scheletul pomului (fig. 3.1) este alcătuit din ramuri de ordinul I (la pomii de vigoare
mică), ordinele I şi II (la pomii de vigoare mijlocie) şi ordinele I, II şi III (la pomii de vigoare
mare).
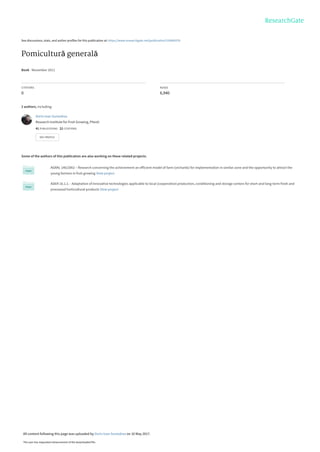
Fig. 3.1. Elementele componente ale tulpinii
Ramurile de semischelet sunt formaţiuni lemnoase multianuale (3-6 ani), garnisite cu
ramuri vegetative şi de rod. Se formează pe axul pomului şi pe ramurile de ordinele I, II şi III.
Ramurile de rod sunt formaţiuni lemnoase anuale, de dimensiuni mici, laterale sau
terminale, de vigoare slabă, fără sau cu suport mai vechi de lemn, purtătoare de muguri
floriferi, micşti sau numai vegetativi, în devenire floriferi.
La speciile sâmburoase ramurile de rod diferenţiază muguri de rod în timpul formării
lor, iar la seminţoase după 2-3 ani de evoluţie (cu mici excepţii).
Comparativ cu ramurile vegetative, cele de rod sunt mai scurte, mai groase şi cu
ţesuturile sfărâmicioase datorită conţinutului mai ridicat în substanţe minerale şi amidon.
Durata lor de viaţă este mai scurtă (1-2 ani) sau mai lungă (10-15 ani), în funcţie de
particularităţile biologice ale speciei şi de agrotehnica aplicată.
Pe ramurile principale şi ramificaţiile secundare se întâlnesc ramuri anuale, care după
poziţia pe care o ocupă pe schelet pot fi: terminale, laterale, concurente, lacome şi anticipate
(fig. 3.2).
28. 26
Fig. 3.2. Clasificarea ramurilor anuale după poziţie (după Cepoiu N., 2001)
1 – terminale; 2 – laterale; 3 – anticipate; 4 – concurente; 5 - lacome
Ramurile terminale (de prelungire) se formează dintr-un mugure terminal şi asigură în
continuare alungirea ramurilor de schelet.
Ramurile laterale se formează din mugurii axilari, descresc ca vigoare de la vârf către
baza ramurii mamă, şi îndeplinesc funcţii de creştere şi de rodire.
Ramura concurentă este prima ramură laterală subterminală, care tinde să concureze în
creştere ramura de prelungire.
Ramura anticipată este apropiată ca vârstă de ramura anuală-mamă din care s-a format
prin evoluţia anticipată a unui mugure axilar.
Ramura lacomă se formează din rezerva de muguri dorminzi în urma tăierilor de
reducţie şi regenerare, sau uscarea centripetă a scheletului.
Unghiul de inserţie (sau unghiul de ramificare) este unghiul format în plan vertical de
ramura principală (de ordinul I) cu axul pomului. În condiţii de dirijare valoarea unghiului de
inserţie se modifică în raport cu forma coroanei, de la 30o
(vas, ypsillon) şi 90o
(la palmetele
cu braţele orizontale).
Pentru majoritatea formelor de coroană, valoarea optimă a unghiului de inserţie, este
cuprinsă între 45 – 55o
.
Unghiul de divergenţă (unghiul de deschidere) este unghiul format în plan orizontal de
două ramuri prinse succesiv pe ax.
Distanţa dintre etaje reprezintă distanţa pe ax, între ultima şarpantă a etajului inferior
şi prima şarpantă a etajului imediat superior. Valoarea acestor distanţe se stabileşte în funcţie
de vigoarea pomului şi sistemul de coroană (40 – 120 cm).
În condiţiile conducerii dirijate a pomilor, unele elemente componente ale coroanelor
naturale, sunt cunoscute sub alte denumiri tehnice. Astfel ramurile de ordinul I devin braţe sau
şarpante, cele de ordinul II subşarpante, iar unghiul de inserţie, se transformă în unghi de
ramificare.
Mugurii - sunt organe de creştere şi rodire cu structura anatomo-morfologică diferită:
Mugurii vegetativi (de creştere) sunt mai mici, au vârful ascuţit şi baza lărgită şi o
structură alcătuită dintr-un con de creştere, primordii de frunze şi muguri. Din ei se formează
întotdeauna lăstari sau rozete de frunze (fig. 3.3).
Mugurii de rod sunt mai mari (cu mici excepţii) mai bombaţi şi cu vârful rotunjit.
Structura internă este alcătuită din primordiile florale (fig. 3.4).
După organele pe care le generează, mugurii sunt: vegetativi, floriferi şi micşti.
29. 27
Fig. 3.3. Mugure vegetativ (după Cepoiu N., 2001)
1 – aspect exterior; 2 – structura internă
Fig. 3.4. Mugure de rod (după Cepoiu N., 2001)
1 – aspect exterior; 2 – structura internă
Mugurii floriferi pot evolua într-o floare, la piersic, cais şi migdal, o inflorescenţă cu
2-5 flori la prun, cireş şi vişin (fig. 3.5).
Mugurii micşti evoluează într-o rozetă de frunze şi o inflorescenţă cu 5-7 flori (la măr,
păr), într-un lăstar şi o floare (la gutui).
După poziţia lor pe ramură, mugurii vegetativi au fost numiţi: terminali, axilari,
stipelari, dorminzi.
Mugurii terminali sunt situaţi în vârful ramurilor anuale şi îndeplinesc funcţia de
creştere şi prelungire (muguri vegetativi), sau de rodire (mugurii micşti).
Mugurii axilari (laterali) se formează în axila frunzei şi descresc ca mărime de la vârf
către baza ramurii.
Aşezarea mugurilor axilari, pe ramură este solitară la măr, păr, cireş, vişin, etc., în
grupuri de câte 2-3 la prun şi piersic, sau 3-8 la cais (fig. 3.6). În cadrul grupului mugurele
central este vegetativ, iar cei laterali, suplimentari, floriferi.
Mugurii suplimentari sunt cunoscuţi sub numele de stipelari şi au o aşezare colaterală
la piersic şi cais, şi serială la nuc (fig. 3.6).
Mugurii dorminzi sunt muguri axilari, incomplet dezvoltaţi, situaţi la inserţia
ramurilor şi constituie rezerva de muguri a pomului. Din ei se formează ramurile lacome.
Frunza. La noduri lăstarii poartă frunze. Acestea au rolul de a sintetiza hrana necesară
pentru creşterea organelor vegetative, cât şi pentru formarea frunzelor. Forma limbului
frunzelor, mărimea lui, lungimea peţiolului etc. constituie caractere care permit identificarea
speciilor şi soiurilor.
Important pentru pomicultor este faptul că, prin lucrările de îngrijire, întregul frunziş
al pomilor trebuie păstrat sănătos, neatacat de boli şi dăunători, prin aplicarea la timp a
tratamentelor fitosanitare, astfel încât să poată sintetiza cu întreaga sa capacitate.
30. 28
a b
Fig. 3.5 Evoluţia mugurilor de rod la măr (a), păr (b), cireş (c) şi piersic (d)
c d
Fig. 3.6. Modul de inserţie a mugurilor laterali pe ramură (după Cepoiu N., 2001)
1 – solitari; 2 – în grup – colaterali; 3 – seriali
31. 29
Frunzele speciilor pomicole cultivate, de diferite forme şi tipuri, sunt prezentate în
figura 3.7.
Fig. 3.7. Forme de frunze
1 – lanceolată; 2 – ovală; 3 – eliptică; 4 – obovată; 5 – subrotundă; 6 – simplă cu lobul întreg;
7 – lobată; 8 – palmat compusă; 9, 10 – imparipenat compuse
Floarea este organul de înmulţire dar şi cel care asigură recolta. Florile se formează în
mugurii de rod (floriferi sau micşti). La rozacee, care constituie majoritatea speciilor
pomicole, florile sunt hermafrodite, au celulele sexuale mascule şi femele normal dezvoltate
în aceiaşi floare (măr, păr, prun, cireş, vişin, cais, piersic, etc). La unele dintre speciile
pomicole (nuc, alun, castan), florile sunt unisexuate, prevăzute cu androceu sau gineceu, dar
ambele pe aceiaşi plantă (monoice). Există specii şi unisexuat dioice, la care fiecare individ
are numai un tip de floare, existând plante mascule şi plante femele (kiwi).
La majoritatea speciilor pomicole: măr, păr, prun, cireş, vişin, nuc, alun şi castan
florile sunt grupate în inflorescenţe. La măr şi păr inflorescenţa este corimb; la cireş şi vişin,
corimb umbeliform; la căpşun, cimă bipară; la coacăz, racem; la nuc şi castan, ament. Flori
solitare apar la gutui, moşmon, cais, piersic, porumbar şi corcoduş.
Fructul. După “legare” (fecundare), ovulele se transformă în seminţe, iar ovarul,
uneori împreună cu alte părţi ale florii, se transformă în fruct.
Majoritatea speciilor pomicole formează fructe cărnoase, iar un mic număr dintre ele
(alun, castan) formează fructe uscate.
Fructele cărnoase ale speciilor pomicole din ţara noastră sunt:
- poama este un fruct simplu, la alcătuirea căruia participă, alături de ovar şi
receptaculul îngroşat, motiv pentru care este numit “fruct fals”. Poama este caracteristică
pentru măr, păr şi gutui. Partea comestibilă, cărnoasă, este formată din receptacul şi partea
exterioară a ovarului. Endocarpul cartilaginos formează lojile seminale (fig. 3.8);
- drupa este un fruct simplu, la alcătuirea căruia participă numai ovarul. Partea
cărnoasă, comestibilă a drupei este alcătuită de mezocarp. Endocarpul lignificat formează
sâmburele, care închide sămânţa. Drupa este caracteristică pentru prun, cais, piersic, cireş,
vişin, corcoduş (fig. 3.8).
Fructele nucului şi ale migdalului sunt pseudodrupe, la maturitatea cărora mezocarpul
(necomestibil) se usucă, crapă si eliberează endocarpul, iar partea comestibilă este, de fapt,
sămânţa.
32. 30
La zmeur, ceea ce se consumă este o polidrupă, o asociaţie de drupeole.
- baciforme: baca este un fruct simplu, întâlnit la coacăz, afin şi pseudobacă
(concreşterea axului florii cu ovarul) la agriş (fig. 3.8). Seminţele rezultate din ovulele
fecundate sunt înglobate în pulpă.
Receptaculul îngroşat, numit impropriu fruct, este caracteristic pentru căpşun. Fructele
propriu-zise sunt mici achene (poliachene), prinse la exteriorul receptaculului.
Fructele uscate sunt reprezentate, în pomicultură, de alună şi castană. Acestea sunt
achene cu pericarpul lemnos.
Sămânţa – este organul generativ al pomului, care rezultă din dezvoltare ovulei
fecundate. Sămânţa este alcătuită dintr-un înveliş exterior numit tegument, endosperm şi
embrion, care la rândului este format din cotiledoane, tulpiniţă şi rădăcinuţă).
Fig. 3.8. Tipuri de fructe
a – poamă; b – drupă; c – polidrupă; d – pseudodrupă (migdal); e – achenă (alun);
f – poliachenă (căpşun); g – bacă (coacăz); h – pseudobacă (agriş); i – hesperidă (lămâi);
j – siconă (smochin)
Seminţele speciilor pomicole pot fi utilizate în alimentaţie (nucile, alunele, castanele,
migdalele), pentru producerea portaltoilor generativi şi pentru ameliorarea speciilor pomicole,
prin hibridări controlate.
3.3. POLARITATEA ŞI DOMINANŢA APICALĂ
Polaritatea. Reprezintă proprietatea celulelor, ţesuturilor, organelor sau a plantelor
întregi de a avea doi poli morfofiziologic determinaţi: polul apical - emite tulpină (lăstari) şi
polul bazal - emite rădăcină.
Acumularea auxinei la polul bazal ca urmare a circulaţiei sale bazipetale, stimulează
formarea rădăcinilor. Astfel, în cazul butaşilor, se consideră că polaritatea este determinată de
circulaţia bazipetală a auxinei, care este cauzată de amplasarea transportorilor specifici în
partea bazală a acestora. Ca urmare, în zona apicală unde se biosintetizează acest hormon se
formează întotdeauna organe aeriene, în timp ce la partea bazală, spre care circulă auxina, se
33. 31
formează rădăcini. Dacă se inversează poziţia butaşilor, locul de formare a rădăcinilor şi
respectiv a organelor aeriene nu se modifică (Burzo I. şi colab., 1999).
Rizogeneza poate fi stimulată prin introducerea butaşilor în auxină soluţie sau sub
formă de pulbere. Rezultate bune s-au obţinut şi în cazul tratării butaşilor cu auxine sintetice:
acid beta-indolil butiric, acid 2,4 – diclorfenoxiacetic (2,4D) şi cu acid alfa-naftilacetic
(NAA).
Polaritatea la plantele pomicole se manifestă numai la organele axiale (lăstari,
rădăcini), nu şi la frunze.
La organele axiale pe lângă polaritatea manifestată de-a lungul axei lor se distinge şi o
polaritate transversală. Un butaş aşezat culcat pe substrat nutritiv emite lăstari din mugurii
situaţi pe partea superioară şi rădăcini din ţesuturile situate pe partea inferioară.
Dominanţa apicală. Reprezintă o corelaţie de creştere prin care mugurele terminal
poate inhiba creşterea mugurilor laterali (corelaţia de creştere = interacţiuni fiziologice între
organe şi ţesuturi, care conduc la creşterea armonioasă a plantelor). Astfel, în cazul lăstarilor,
mugurii axilari formaţi la subsoara frunzelor nu pornesc în creştere decât dacă se ciupeşte
vârful lor, iar la reluarea creşterii, primăvara, mugurii axilari formează lăstari cu atât mai mici
cu cât sunt mai îndepărtaţi de mugurele terminal, din care rezultă cel mai puternic lăstar.
Pentru explicarea mecanismului dominanţei apicale au fost elaborate mai multe
ipoteze (Burzo I. şi colab., 1999):
* Ipoteza carenţei trofice susţine ca mugurii axilari nu se dezvoltă deoarece mugurele apical
este aprovizionat în mod preferenţial cu substanţe nutritive.
* Ipoteza inhibiţiei directe sau a controlului hormonal. Aceasta presupune participarea
tuturor hormonilor, dar rolul lor nu a fost bine precizat. Thimann, (1984), arată că,
auxinele biosintetizate în mugurii apicali sunt transportate bazipetal şi concentraţia mărită
din apropierea mugurilor laterali are efect inhibitor asupra creşterii acestora. Acidul
abscizic are efect sinergic cu al auxinei, în timp ce giberelinele şi citokininele au un efect
antagonist.
* Ipoteza inhibiţiei indirecte A fost elaborată de Snow (1924), care a dovedit că, dominanţa
apicală este determinată de prezenţa unui inhibitor specific, iar auxina anulează efectul
acestuia.
* Ipoteza diversiunii nutritive, a lui Went (1936), îmbină ipoteza carenţei trofice cu cea a
controlului hormonal. Conform acestei ipoteze auxina endogenă generează un curent de
substanţe nutritive către mugurii apicali, în detrimentul celor laterali.
Dominanţa apicală se manifestă şi în cazul rădăcinilor, apexul rădăcinii principale
având un efect inhibitor asupra creşterii rădăcinilor laterale.
Observaţiile referitoare la aspectul rădăcinii pomilor fructiferi, scot în evidenţă faptul
că rădăcinile situate în apropierea tulpinii, sunt mai lungi comparativ cu cele din zona apicală.
Aceasta se datorează procesului de dominanţă apicală, care se manifestă şi la nivelul
rădăcinilor. Auxina sintetizată la nivelul mugurilor apicali ai organelor aeriene, este
transportată bazipetal şi cantităţile mici ajunse la nivelul rădăcinilor stimulează procesul de
creştere al acestora. În acelaşi timp, citochininele şi acidul abscizic sintetizate în apexul
rădăcinii au un efect inhibitor asupra creşterii rădăcinilor secundare. Rolul celorlalţi hormoni
în dominanţa apicală a rădăcinii, nu este bine precizat (Burzo I. şi colab., 2005). Unii autori,
Kolek şi Kozinka, (1992), apreciază că în acest caz dominanţa apicală este determinată de
citokinine, iar auxinele au un efect inhibitor asupra acestui proces.
Polaritatea şi dominanţa apicală sunt procese fiziologice importante şi cunoaşterea
acestora stă la baza fundamentării şi aplicării corecte a verigilor tehnologice legate de
controlul creşterii şi fructificării speciilor pomicole prin tăieri sau prin tratamente hormonale.
34. 32
Capitolul 4
CICLUL DE VIAŢĂ AL SPECIILOR POMICOLE
4.1. PERIOADELE DE VÂRSTĂ
Ciclul de viaţă reprezintă totalitatea schimbărilor morfologice şi fiziologice parcurse
de pomi şi arbuştii fructiferi de la formarea zigotului şi embrionului până la moartea acestora.
În primele etape ale ciclului de viaţă, la speciile pomicole, ca la toate organismele
tinere, caracterele şi însuşirile sunt insuficient consolidate şi, în consecinţă manifestă
plasticitate şi posibilităţi de a asimila condiţii noi. Organismele mature (bătrâne) sunt lipsite
de aceste proprietăţi. Procesul de îmbătrânire la speciile pomicole este discontinuu, fiind
întrerupt de perioade cu o mărire temporară a potenţialului de creştere şi fructificare, dar fără
să atingă potenţialul iniţial.
În urma diviziunii celulare din vârfurile meristematice are loc o întinerire parţială
deoarece celulele nou apărute sunt mai tinere decât cele învecinate. Ţesuturile meristematice
existente în diferite părţi ale plantei posedă însuşiri caracteristice tinereţii (plasticitate,
asimilare de noi condiţii), iar ţesuturile definitive învecinate sunt lipsite de aceste
particularităţi. Astfel, pe orice lăstar în creştere, celulele noi se adaugă la vârf, fiind situate pe
celule şi ţesuturi definitive, mai bătrâne ca vârstă, lipsite de plasticitate. Spre deosebire de
acestea, celulele tinere asimilează condiţii noi, evoluează, ating un grad de dezvoltare mai
mare.
Rezultatul acestor procese este existenţa unei diferenţieri calitative (denumită frecvent
şi diferenţiere stadială) de-a lungul tulpinii sau al unei ramuri: la bază ţesuturile sunt mai puţin
evoluate, mai tinere stadial, deşi ca vârstă calendaristică sunt mai bătrâne, iar lăstarii mai
tineri au acumulat mai multe condiţii noi, sunt mai evoluaţi, mai bătrâni stadial.
În coroana fiecărui pom există ramuri de toate vârstele, de la cele de un an până la cele
mai bătrâne (trunchi, baza şarpantelor), dar care sunt mai puţin evoluate (tinere stadial).
Astfel, de exemplu un lăstar apărut la coletul pomului are cu totul alte însuşiri faţă de un lăstar
format în aceiaşi perioadă de vegetaţie în partea superioară a coroanei. Aceiaşi situaţie este în
cazul comparării unui lăstar lacom format dintr-un mugure dormind şi un lăstar de aceiaşi
vârstă din partea superioară a coroanei (Miliţiu I., 1992).
Indiferent de durata ciclului de viaţă (10-15 ani la arbuşti, până la peste 100 de ani la
nuc şi castan), ţinând cont de modificările morfologice şi fiziologice ce au loc pe parcursul
acesteia, au fost delimitate 6 perioade de vârstă, a căror cunoaştere este necesară pentru
fundamentarea şi stabilirea diferitelor verigi tehnologice.
1. Perioada embrionară. Începe cu fecundarea ovulului şi durează până la
germinarea seminţei, exteriorizată prin alungirea radicelei.
Datorită faptului că, majoritatea speciilor pomicole sunt adaptate la polenizarea
încrucişată, noul individ rezultat este un hibrid. În consecinţă, plasticitatea şi capacitatea
acestuia de a asimila condiţii noi, de a fi influenţat de condiţiile de mediu sunt mai mari decât
în cazul plantelor autogame. Astfel, se explică faptul că, din seminţele soiurilor cultivate (cele
cu polenizare încrucişată), cel mai adesea se obţin puieţi cu însuşiri inferioare faţă de ambele
soiuri care au participat la formarea noului embrion. Seminţele fiind obţinute prin polenizare
încrucişată dau naştere la pomi hibrizi, nepăstrând caracterele plantelor mamă.
De asemenea, în cazul speciilor pomicole noul embrion este influenţat şi de portaltoiul
pe care acesta se formează. Ca urmare, pentru a evita influenţa portaltoiului este de preferat ca
în lucrările de hibridare artificială să se utilizeze pomi crescuţi pe rădăcini proprii (Miliţiu I.,
1992).
35. 33
După fecundare are loc formarea elementelor constitutive ale seminţei: embrionul şi
cotiledoanele. Completa formare a acestora marchează maturitatea morfologică a seminţei,
care are loc pe planta mamă; procesul este însoţit de o deshidratare accentuată, ajungându-se
în final la cca. 10% apă. În urma acestui proces metabolismul seminţei se reduce foarte mult,
iar în această situaţie sămânţa poate suporta condiţii nefavorabile (temperaturi scăzute,
uscăciune).
Chiar dacă sunt mature morfologic, seminţele pomilor nu pot germina în condiţii
favorabile de umiditate şi temperatură, deoarece nu au ajuns la maturitatea fiziologică (sunt în
repaus).
Maturitatea fiziologică (postmaturarea) nu presupune modificări morfologice, ea se
realizează în condiţii de umiditate şi temperatură bine definite. Pentru realizarea
postmaturării, în general, seminţele pomilor au nevoie de temperaturi de 1-4o
C, de umiditate
de cca. 30% şi de aerisire.
La seminţele căzute pe sol împreună cu fructele, în mod natural, postmaturarea se
desfăşoară începând din toamnă când temperatura scade sub 4o
C şi există umiditate suficientă.
Procesul se întrerupe când temperatura scade sub 0o
C şi reîncepe primăvara când se
înregistrează condiţiile necesare.
În tehnologia pomicolă, condiţiile necesare postmaturării se realizează prin procesul
denumit stratificare. Durata postmaturaţiei este o caracteristică de specie: la cele cu coacere
timpurie (cireş, vişin) este mai lungă decât la cele cu coacere târzie (măr, păr).
Perioada embrionară se încheie cu procesul complex de germinare a seminţelor.
În pomicultură, perioada embrionară este întâlnită numai în tehnologia de producere a
portaltoilor generativi, în activitatea de producere a soiurilor noi şi în cazul unor soiuri de nuc
şi castan.
2. Perioada juvenilă (de tinereţe sau de creştere). Această perioadă începe cu
germinarea seminţelor, concretizată prin alungirea rădăcinii (la înmulţirea prin seminţe), sau
cu pornirea în vegetaţie a altoiului (la înmulţirea prin altoire) şi se încheie cu apariţia primelor
flori şi fructe. În plantaţiile superintensive de măr durează 1-2 ani, la piersic 2-3 ani, la prun şi
cais 3-4 ani şi ajunge la 8-12 ani la nucul nealtoit şi castanul din seminţe (Drăgănescu E.,
1998).
În această perioadă, an de an se formează noi organe care se adaugă celor precedente,
fenomen denumit înnoire progresivă.
Perioada juvenilă se caracterizează prin creşterea puternică atât a sistemului radicular
cât şi a părţii aeriene a pomilor. În perioada de tinereţe pomii au creşteri viguroase, care pot
depăşi 1-1,5 m, vegetaţia se prelungeşte până toamna târziu, ramurile nu se pot căli suficient
şi, astfel rezistenţa lor la gerurile mari de iarnă este mai redusă. Pentru a nu apărea astfel de
accidente trebuie să se urmărească evitarea excesului de azot şi de apă în a 2-a jumătate a
verii.
Polaritatea şi dominanţa apicală se manifestă puternic, apare frecvent etajarea naturală,
lăstarii au tendinţa de creştere verticală, coroanele fiind strânse, dese. Către sfârşitul acestei
perioade se formează ramuri de rod “în devenire”.
Lucrările care se execută în această perioadă sunt: tăierile de formare a coroanelor,
fertilizarea, irigarea, tratamentele fitosanitare, instalarea sistemelor de susţinere (unde este
cazul).
Formarea coroanelor (cea mai mare parte a scheletului), conform parametrilor tipului
de coroană ales, trebuie să aibă ca obiectiv scurtarea perioadei de tinereţe şi grăbirea intrării
pe rod. În acest sens este de dorit să se reducă tăierile la maximum posibil şi să se utilizeze cât
mai mult înclinarea şi arcuirea ramurilor.
3. Începutul rodirii. Începe de la apariţia primelor fructe şi durează până la obţinerea
recoltelor maxime, susţinute an de an. Durata acestei perioade depinde de specie, soi, portaltoi