Peptidomics Methods and Protocols 1st Edition Mikhail Soloviev (Auth.)
1.
Visit https://ebookultra.com todownload the full version and
explore more ebooks
Peptidomics Methods and Protocols 1st Edition
Mikhail Soloviev (Auth.)
_____ Click the link below to download _____
https://ebookultra.com/download/peptidomics-methods-
and-protocols-1st-edition-mikhail-soloviev-auth/
Explore and download more ebooks at ebookultra.com
2.
Here are somesuggested products you might be interested in.
Click the link to download
Polyadenylation Methods and Protocols 1st Edition Joanna
Rorbach
https://ebookultra.com/download/polyadenylation-methods-and-
protocols-1st-edition-joanna-rorbach/
Microsatellites Methods and Protocols 1st Edition
Panagiotis Madesis
https://ebookultra.com/download/microsatellites-methods-and-
protocols-1st-edition-panagiotis-madesis/
Biofuels Methods and Protocols 1st Edition Shahabaddine
Sokhansanj
https://ebookultra.com/download/biofuels-methods-and-protocols-1st-
edition-shahabaddine-sokhansanj/
Liposome Methods and Protocols 1st Edition Subroto
Chatterjee
https://ebookultra.com/download/liposome-methods-and-protocols-1st-
edition-subroto-chatterjee/
3.
Apoptosis Methods andProtocols 1st Edition Demetrius
Matassov
https://ebookultra.com/download/apoptosis-methods-and-protocols-1st-
edition-demetrius-matassov/
Xenotransplantation Methods and Protocols 1st Edition
Veronica Tisato
https://ebookultra.com/download/xenotransplantation-methods-and-
protocols-1st-edition-veronica-tisato/
Protoplast Technology Methods and Protocols 1st Edition
Kan Wang
https://ebookultra.com/download/protoplast-technology-methods-and-
protocols-1st-edition-kan-wang/
Chromatin Remodeling Methods and Protocols 1st Edition
Junbiao Dai
https://ebookultra.com/download/chromatin-remodeling-methods-and-
protocols-1st-edition-junbiao-dai/
Nanoimaging Methods and Protocols 1st Edition Alioscka A.
Sousa
https://ebookultra.com/download/nanoimaging-methods-and-protocols-1st-
edition-alioscka-a-sousa/
METHODS IN MOLECULARBIOLOGY
TM
Series Editor
John M. Walker
School of Life Sciences
University of Hertfordshire
Hatfield, Hertfordshire, AL10 9AB, UK
For other titles published in this series, go to
www.springer.com/series/7651
Preface
Despite being knownand studied for years, peptides have never before attracted enough
attention to necessitate the invention of the term “Peptidomics” in order to specify the
study of the complement of peptides from a cell, organelle, tissue or organism. This vol-
ume presents a comprehensive range of analytical techniques for analysis of the peptide
contents of complex biological samples. The emphasis is often on higher throughput
techniques, suitable for the analysis of large numbers of peptides typically present in the
peptidomes or other complex biological samples. A wide range of methods is presented,
covering all stages of peptidomic research including, where applicable, organism handling,
tissue and organ dissection, cellular and subcellular fractionation, peptide extraction, frac-
tionation and purification, structural characterisation, molecular cloning and sequence
analysis. In addition to this, a selection of methods suitable for quantification, display,
immunochemical and functional analysis of peptides and proteins are presented. The
methods and techniques covered in this volume encompass a number of species rang-
ing from bacteria to man and include model organisms such as Caenorhabditis elegans,
Drosophila melanogaster and Mus musculus. Strong emphasis is placed on data analysis,
including mass spectra interpretation and in silico peptide prediction algorithms. Where
relevant, the peptidomic approaches are compared to the proteomic methods. Here is a
snapshot of the practical information, peptidomic methods and other related protocols
included in this volume:
Target organisms and samples covered: Bacteria (Chapter 2), hydra (Chapter 21), nema-
tode (Chapter 3), mollusc (Chapter 4), crab (Chapter 5), spider venoms (Chapters 6
and 7), insects (Chapters 8, 9, 10, 11, 25), amphibians (Chapters 12, 13, 14), rodents
(Chapters 15, 16, 17, 18), samples of human origin (Chapters 19, 20, 22, 23) and plants
(Chapter 26).
Peptide extraction and Liquid chromatography fractionation methods (mostly size
exclusion, ion exchange, reverse-phase modes or their combinations) can be found in
Chapters 2, 3, 4, 5, 6, 7, 8, 12, 14, 15, 16, 17, 18, 19, 20, 21, 22). These include OFF-
line and ON-line techniques. The former are often used with MALDI-MS detection (e.g.
Chapters 3, 4, 5, 6, 7, 8, 12, 15, 16, 19) whilst the latter more generally with single or
multidimensional hyphenated LCN
-MSN
techniques (e.g. Chapters 2, 3, 15, 17, 18, 21).
Other separation and fractionation methods covered include microdialysis of live ani-
mals (Chapter 5), SDS-PAGE (Chapters 6, 18), magnetic bead based purification (Chap-
ter 20) and solid-phase extraction (Chapters 2, 6, 12, 19, 22).
Affinity peptide detection including anti-peptide antibody development and characteri-
sation, Affinity peptidomics, ELISA and microarray affinity assays are covered in Chapters
22, 23 and 24.
Mass spectrometry techniques include MALDI-TOF MS (e.g. Chapters 3, 6, 7, 8, 12,
16, 19, 20), MALDI-TOF with PSD (Chapter 8), MALDI-TOF MS/MS (e.g. Chapters
4, 6, 15, 21); ESI-MS/MS techniques (Chapters 3, 6, 16, 17, 18) or high-resolution
FTMS (Chapter 2). Direct MALDI-MS peptide profiling from cells and tissues is described
in Chapters 9, 10 and 11.
v
11.
vi Preface
The descriptionof functional assays can be found in Chapters 7, 14 and 21. Of par-
ticular interest in this respect is Chapter 21, where functional activity of the peptides is
assessed through the analysis of mRNA transcription levels changes in response to the
peptide application. That chapter contains a selection of protocols for peptide extraction,
fractionation and functional testing using a combination of molecular biology techniques,
cellular and morphological assays.
Molecular cloning of peptide cDNAs and the associated techniques are described in
Chapters 13 and 14.
Issues related to peptide sequence analysis are addressed in many chapters dealing with
MS spectra interpretation, but of special interest in this respect are Chapters 25 and 26,
dealing with in silico peptide prediction techniques and Chapter 20 which includes a section
on bioinformatics analysis of peptide expression profiling data. Differential peptide expression
issues are also covered in Chapter 2.
Peptidomics is 10-years old. My congratulations go to all scientists who have created
and developed the science of Peptidomics through their research and especially those who
found time to contribute their invaluable know-how in the form of methods and protocols
for inclusion in this volume. Peptidomics: Methods and Protocols is designed to complement
previously published titles in the Methods in Molecular BiologyTM
series, which focused on
protein analysis. This volume will help the beginner to become familiar with this fascinating
field of research and will provide scientists at all levels of expertise with easy-to-follow
practical advice needed to set up and carry out analysis of the peptide contents of complex
biological samples.
Royal Holloway University of London Mikhail Soloviev
December 2009
12.
Contents
Preface . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . v
Contributors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . xi
SECTION I INTRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
1. Peptidomics: Divide et Impera . . . . . . . . . . . . . . . . . . . . . . . . . . 3
Mikhail Soloviev
SECTION II FROM BACTERIA TO MEN . . . . . . . . . . . . . . . . . . . . . . . . 11
2. Performing Comparative Peptidomics Analyses
of Salmonella from Different Growth Conditions . . . . . . . . . . . . . . . . . 13
Joshua N. Adkins, Heather Mottaz, Thomas O. Metz, Charles Ansong,
Nathan P. Manes, Richard D. Smith, and Fred Heffron
3. Approaches to Identify Endogenous Peptides in the Soil Nematode
Caenorhabditis elegans . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29
Steven J. Husson, Elke Clynen, Kurt Boonen, Tom Janssen,
Marleen Lindemans, Geert Baggerman, and Liliane Schoofs
4. Mass Spectrometric Analysis of Molluscan Neuropeptides . . . . . . . . . . . . 49
Ka Wan Li and August B. Smit
5. Monitoring Neuropeptides In Vivo via Microdialysis
and Mass Spectrometry . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 57
Heidi L. Behrens and Lingjun Li
6. Protocols for Peptidomic Analysis of Spider Venoms . . . . . . . . . . . . . . . 75
Liang Songping
7. Purification and Characterization of Biologically Active Peptides
from Spider Venoms . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87
Alexander A. Vassilevski, Sergey A. Kozlov, Tsezi A. Egorov,
and Eugene V. Grishin
8. MALDI-TOF Mass Spectrometry Approaches
to the Characterisation of Insect Neuropeptides . . . . . . . . . . . . . . . . . 101
Robert J. Weaver and Neil Audsley
9. Direct MALDI-TOF Mass Spectrometric Peptide Profiling
of Neuroendocrine Tissue of Drosophila . . . . . . . . . . . . . . . . . . . . . 117
Christian Wegener, Susanne Neupert, and Reinhard Predel
10. Direct Peptide Profiling of Brain Tissue by MALDI-TOF
Mass Spectrometry . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 129
Joachim Schachtner, Christian Wegener, Susanne Neupert,
and Reinhard Predel
vii
13.
viii Contents
11. PeptidomicAnalysis of Single Identified Neurons . . . . . . . . . . . . . . . . . 137
Susanne Neupert and Reinhard Predel
12. Identification and Analysis of Bioactive Peptides
in Amphibian Skin Secretions . . . . . . . . . . . . . . . . . . . . . . . . . . . 145
J. Michael Conlon and Jérôme Leprince
13. An Efficient Protocol for DNA Amplification of Multiple Amphibian
Skin Antimicrobial Peptide cDNAs . . . . . . . . . . . . . . . . . . . . . . . . 159
Shawichi Iwamuro and Tetsuya Kobayashi
14. Combined Peptidomics and Genomics Approach to the Isolation
of Amphibian Antimicrobial Peptides . . . . . . . . . . . . . . . . . . . . . . . 177
Ren Lai
15. Identification and Relative Quantification of Neuropeptides
from the Endocrine Tissues . . . . . . . . . . . . . . . . . . . . . . . . . . . . 191
Kurt Boonen, Steven J. Husson, Bart Landuyt, Geert Baggerman,
Eisuke Hayakawa, Walter H.M.L. Luyten, and Liliane Schoofs
16. Peptidome Analysis of Mouse Liver Tissue by Size Exclusion
Chromatography Prefractionation . . . . . . . . . . . . . . . . . . . . . . . . . 207
Lianghai Hu, Mingliang Ye, and Hanfa Zou
17. Rat Brain Neuropeptidomics: Tissue Collection, Protease Inhibition,
Neuropeptide Extraction, and Mass Spectrometric Analysis . . . . . . . . . . . . 217
Robert M. Sturm, James A. Dowell, and Lingjun Li
18. Quantitative Neuroproteomics of the Synapse . . . . . . . . . . . . . . . . . . 227
Dinah Lee Ramos-Ortolaza, Ittai Bushlin, Noura Abul-Husn,
Suresh P. Annangudi, Jonathan Sweedler, and Lakshmi A. Devi
19. Peptidomics Analysis of Lymphoblastoid Cell Lines . . . . . . . . . . . . . . . 247
Anne Fogli and Philippe Bulet
20. Peptidomics: Identification of Pathogenic and Marker Peptides . . . . . . . . . . 259
Yang Xiang, Manae S. Kurokawa, Mie Kanke, Yukiko Takakuwa,
and Tomohiro Kato
SECTION III TOOLS AND APPROACHES . . . . . . . . . . . . . . . . . . . . . . . . 273
21. Peptidomic Approaches to the Identification and Characterization
of Functional Peptides in Hydra . . . . . . . . . . . . . . . . . . . . . . . . . 275
Toshio Takahashi and Toshitaka Fujisawa
22. Immunochemical Methods for the Peptidomic Analysis
of Tachykinin Peptides and Their Precursors . . . . . . . . . . . . . . . . . . . 293
Nigel M. Page and Nicola J. Weston-Bell
23. Affinity Peptidomics: Peptide Selection and Affinity Capture
on Hydrogels and Microarrays . . . . . . . . . . . . . . . . . . . . . . . . . . 313
Fan Zhang, Anna Dulneva, Julian Bailes, and Mikhail Soloviev
14.
Contents ix
24. InSitu Biosynthesis of Peptide Arrays . . . . . . . . . . . . . . . . . . . . . . . 345
Mingyue He and Oda Stoevesandt
25. Bioinformatic Approaches to the Identification of Novel Neuropeptide
Precursors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 357
Elke Clynen, Feng Liu, Steven J. Husson, Bart Landuyt,
Eisuke Hayakawa, Geert Baggerman, Geert Wets, and Liliane Schoofs
26. Bioinformatic Identification of Plant Peptides . . . . . . . . . . . . . . . . . . . 375
Kevin A. Lease and John C. Walker
Index . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 385
15.
Contributors
NOURA ABUL-HUSN •Department of Pharmacology and Systems Therapeutics, Mount
Sinai School of Medicine, New York, NY, USA
JOSHUA N. ADKINS • Fundamental and Computational Sciences Directorate, Pacific
Northwest National Laboratory, Richland, WA, USA
SURESH P. ANNAGUDI • Department of Chemistry and the Beckman Institute,
University of Illinois at Urbana-Champaign, Urbana, IL, USA
CHARLES ANSONG • Fundamental and Computational Sciences Directorate, Pacific
Northwest National Laboratory, Richland, WA, USA
NEIL AUDSLEY • The Food and Environment Research Agency, Sand Hutton, York, UK
GEERT BAGGERMAN • ProMeta, Interfacultary Center for Proteomics and
Metabolomics, K.U. Leuven, Leuven, Belgium
JULIAN BAILES • School of Biological Sciences, Royal Holloway University of London,
Egham, Surrey, UK
HEIDI L. BEHRENS • Department of Chemistry, University of Wisconsin-Madison,
Madison, WI, USA
KURT BOONEN • Functional Genomics and Proteomics Research Unit, Department of
Biology, K.U. Leuven, Leuven, Belgium
PHILIPPE BULET • TIMC-IMAG, UMR 5525, Domaine de Chosal, Archamps, France
ITTAI BUSHLIN • Department of Pharmacology and Systems Therapeutics, Mount Sinai
School of Medicine, New York, NY, USA
ELKE CLYNEN • Functional Genomics and Proteomics, Department of Biology, K.U.
Leuven, Leuven, Belgium
J. MICHAEL CONLON • Department of Biochemistry, Faculty of Medicine and Health
Sciences, United Arab Emirates University, Al-Ain, UAE
LAKSHMI A. DEVI • Department of Pharmacology and Systems Therapeutics, Mount
Sinai School of Medicine, New York, NY, USA
JAMES A. DOWELL • Department of Chemistry, School of Pharmacy, University of
Wisconsin-Madison, Madison, WI, USA
ANNA DULNEVA • School of Biological Sciences, Royal Holloway University of London,
Egham, Surrey, UK
TSEZI A. EGOROV • Shemyakin-Ovchinnikov Institute of Bioorganic Chemistry, Russian
Academy of Sciences, Moscow, Russia
xi
16.
xii Contributors
ANNE FOGLI• GreD UMR INSERM 931 CNRS 6142, Faculté de Médecine,
Clermont-Ferrand, France
TOSHITAKA FUJISAWA • Institute of Zoology, University of Heidelberg, Heidelberg,
Germany
EUGENE V. GRISHIN • Shemyakin-Ovchinnikov Institute of Bioorganic Chemistry,
Russian Academy of Sciences, Moscow, Russia
EISUKE HAYAKAWA • Functional Genomics and Proteomics Research Unit, Department
of Biology, K.U. Leuven, Leuven, Belgium
MINGYUE HE • The Babraham Institute, Cambridge, UK
FRED HEFFRON • Department of Molecular Microbiology and Immunology, Oregon
Health and Sciences University, Portland, OR, USA
LIANGHAI HU • Key Laboratory of Separation Science for Analytical Chemistry,
National Chromatographic R&A Centre, Dalian Institute of Chemical Physics, Chinese
Academy of Sciences, Dalian, China
STEVEN J. HUSSON • Functional Genomics and Proteomics, Department of Biology,
K.U. Leuven, Leuven, Belgium
SHAWICHI IWAMURO • Department of Biology, Faculty of Science, Toho University,
Funabashi, Chiba, Japan
TOM JANSSEN • Functional Genomics and Proteomics, Department of Biology, K.U.
Leuven, Leuven, Belgium
MIE KANKE • Clinical Proteomics and Molecular Medicine, St. Marianna University
Graduate School of Medicine, Kawasaki, Japan
TOMOHIRO KATO • Clinical Proteomics and Molecular Medicine, St. Marianna
University Graduate School of Medicine, Kawasaki, Japan
TETSUYA KOBAYASHI • Department of Regulation Biology, Faculty of Science, Saitama
University, Saitama, Japan
SERGEY A. KOZLOV • Shemyakin-Ovchinnikov Institute of Bioorganic Chemistry,
Russian Academy of Sciences, Moscow, Russia
MANAE S. KUROKAWA • Clinical Proteomics and Molecular Medicine, St. Marianna
University Graduate School of Medicine, Kawasaki, Japan
REN LAI • Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming,
Yunnan, China
BART LANDUYT • Functional Genomics and Proteomics Research Unit, Department of
Biology, K.U. Leuven, Leuven, Belgium
KEVIN A. LEASE • Division of Biological Sciences and Bond Life Sciences Centre,
University of Missouri, Columbia, MO, USA
JÉRÔME LEPRINCE • European Institute for Peptide Research (IFRMP 23), INSERM
U-413, UA CNRS, University of Rouen, Mont-Saint-Aignan, France
17.
Contributors xiii
KA WANLI • Department of Molecular and Cellular Neurobiology, Centre for
Neurogenomics and Cognitive Research, Faculty of Earth and Life Sciences, VU
University Amsterdam, Amsterdam, The Netherlands
LINGJUN LI • Department of Chemistry, School of Pharmacy, University of
Wisconsin-Madison, Madison, WI, USA
MARLEEN LINDEMANS • Functional Genomics and Proteomics, Department of Biology,
K.U. Leuven, Leuven, Belgium
FENG LIU • Data Analysis and Modeling Group, Transportation Research Institute,
Hasselt University, Diepenbeek, Belgium
WALTER H.M.L. LUYTEN • Department Woman and Child, Faculty of Medicine, K.U.
Leuven, Leuven, Belgium
NATHAN P. MANES • Fundamental and Computational Sciences Directorate, Pacific
Northwest National Laboratory, Richland, WA, USA
THOMAS O. METZ • Fundamental and Computational Sciences Directorate, Pacific
Northwest National Laboratory, Richland, WA, USA
HEATHER MOTTAZ • Fundamental and Computational Sciences Directorate, Pacific
Northwest National Laboratory, Richland, WA, USA
SUSANNE NEUPERT • Institute of General Zoology and Animal Physiology,
Friedrich-Schiller-University, Jena, Germany
NIGEL M. PAGE • School of Life Sciences, Kingston University London,
Kingston-upon-Thames, Surrey, UK
REINHARD PREDEL • Institute of General Zoology and Animal Physiology,
Friedrich-Schiller-University, Jena, Germany
DINAH LEE RAMOS-ORTOLAZA • Department of Pharmacology and Systems
Therapeutics, Mount Sinai School of Medicine, New York, NY, USA
JOACHIM SCHACHTNER • Department of Biology, Animal Physiology,
Philipps-University, Marburg, Germany
LILIANE SCHOOFS • Functional Genomics and Proteomics Research Unit, Department
of Biology, K.U. Leuven, Leuven, Belgium
AUGUST B. SMIT • Department of Molecular and Cellular Neurobiology, Centre for
Neurogenomics and Cognitive Research, Faculty of Earth and Life Sciences, VU
University Amsterdam, Amsterdam, The Netherlands
RICHARD D. SMITH • Fundamental and Computational Sciences Directorate, Pacific
Northwest National Laboratory, Richland, WA, USA
MIKHAIL SOLOVIEV • School of Biological Sciences, Royal Holloway University of
London, Egham, Surrey, UK
LIANG SONGPING • College of Life Sciences, Hunan Normal University, Changsha,
China
ODA STOEVESANDT • Babraham Bioscience Technologies, Cambridge, UK
18.
xiv Contributors
ROBERT M.STURM • Department of Chemistry, School of Pharmacy, University of
Wisconsin-Madison, Madison, WI, USA
JONATHAN SWEEDLER • Department of Chemistry and the Beckman Institute,
University of Illinois at Urbana-Champaign, Urbana, IL, USA
TOSHIO TAKAHASHI • Suntory Institute for Bioorganic Research, Osaka, Japan
YUKIKO TAKAKUWA • Clinical Proteomics and Molecular Medicine, St. Marianna
University Graduate School of Medicine, Kawasaki, Japan
ALEXANDER A. VASSILEVSKI • Shemyakin-Ovchinnikov Institute of Bioorganic
Chemistry, Russian Academy of Sciences, Moscow, Russia
JOHN C. WALKER • Division of Biological Sciences and Bond Life Sciences Center,
University of Missouri, Columbia, MO, USA
ROBERT J. WEAVER • The Food and Environment Research Agency, Sand Hutton, York,
UK
CHRISTIAN WEGENER • Emmy Noether Neuropeptide Group, Animal Physiology,
Philipps-University, Marburg, Germany
NICOLA J. WESTON-BELL • Genetic Vaccine Group, Cancer Sciences Division,
Southampton General Hospital, University of Southampton School of Medicine,
Southampton, Hampshire, UK
GEERT WETS • Data Analysis and Modeling Group, Transportation Research Institute,
Hasselt University, Diepenbeek, Belgium
YANG XIANG • Clinical Proteomics and Molecular Medicine, St. Marianna University
Graduate School of Medicine, Kawasaki, Japan
MINGLIANG YE • Key Laboratory of Separation Science for Analytical Chemistry,
National Chromatographic R&A Centre, Dalian Institute of Chemical Physics, Chinese
Academy of Sciences, Dalian, China
FAN ZHANG • School of Biological Sciences, Royal Holloway University of London, Egham,
Surrey, UK
HANFA ZOU • Key Laboratory of Separation Science for Analytical Chemistry, National
Chromatographic R&A Centre, Dalian Institute of Chemical Physics, Chinese Academy
of Sciences, Dalian, China
4 Soloviev
techniques, resultedin dramatic improvements in the sensitivity
and high throughput of protein and peptide analyses (2) and gen-
erated unprecedented growth in the number of relevant publica-
tions (Fig. 1.1).
0.0%
0.1%
0.2%
0.3%
0.4%
0.5%
0.6%
0.7%
0.8%
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
2007
2008
0.000%
0.001%
0.002%
0.003%
0.004%
0.005%
0.006%
0.007%
0.008%
Peptidomics
Proteomics
Fig. 1.1. Peptidomics publications since 1999. The bars represent the number of publications in PubMed containing
“peptidomics OR peptidomic OR peptidome” normalised to the total number of publications added to PubMed each year.
Proteomics publications in PubMed (found similarly) follow the same trend (solid line). Vertical axes show normalised
data (in %) for proteomics papers (left) and peptidomics papers (right). Total number of publications on 1 January 2009
was 28,273 (proteomics) and 246 (peptidomics).
2. Peptidomics
of Naturally
Occurring
Peptides and
Peptide Pools
Similarly to “proteomics”, the term “peptidomics” can be defined
as the systematic analysis of the peptide content within an organ-
ism, tissue or cell (3) in order to determine peptides’ identity,
quantity, structure and function. Such interpretation made its
public debut at the 2nd International Seminar on the Enabling
Role of MS in 1999 (4), before finally appearing in press in 2001
in research papers by Peter D.E.M. Verhaert et al. (5), Peter
Schulz-Knappe et al. (6) and Elke Clynen et al. (7). The disci-
pline of peptidomics focuses on peptides that often display bio-
logical activity such as hormones, cytokines, toxins, neuropeptides
and alike, which are generated from larger precursors, as well as
biomarker-type peptides that may not have any bioactivity but are
indicative of a particular pathology, for example the up/down-
regulation of many serum peptides that result from proteolysis.
22.
Peptidomics: Divide etImpera 5
Peptidomics is in its infancy relative to other “omics” (28,273
papers on PubMed for a “proteomics” search compared to just
246 for “peptidomics” as of January 1st, 2009) but is expanding
rapidly (Fig. 1.1).
3. Peptidomics
Approach to
Proteomics
In parallel, and independently of the peptidomics definition given
in (5–7), another meaning was introduced by Barry et al. (8)
in relation to the analysis of peptide pools (of biological fluids,
tissues or cells) obtained by means of proteolytic digestion of
these samples and in particular using affinity-based analysis (hence
“Affinity peptidomics”) e.g. in the form of protein arrays (9, 10).
Since biologically occurring peptides (whether biologically active
or not) are strictly speaking also the products of proteolysis (e.g.
insulin pre-pro-insulin, or biologically active peptides obtained
through “non-specific” proteolysis of e.g. haemoglobin), both
definitions of “peptidomics” are therefore very similar in that they
refer to the analysis of partially or fully proteolytically digested
proteins, i.e. peptides. And finally, to acknowledge everyone
involved in the birth of the “peptidomics” as a separate field of
chemical biology, we should mention a Germany-based company
“BioVisioN AG” which filed a trademark “peptidomics” in 1999
to cover “Chemicals used in science, in particular for analysis,
other than for medical or veterinary purposes”; “Medical, veteri-
nary and pharmaceutical products” as well as the “Scientific and
industrial research; conducting medical and non-medical analyses;
services in the field of diagnostics; development of pharmaceutical
active substances; purchasing, licensing and exploitation of intel-
lectual property” (11).
4. Peptidomics:
The
Methodologies
In the past, the majority of separation techniques used in pro-
tein and peptide analysis relied on their physical properties, such
as protein or peptide size, shape, polarity, pI, the distribution of
ionisable, polar and non-polar groups on the molecule surface,
and their affinity towards specific or non-specific affinity capture
reagents. Modern separation techniques rely on a combination of
isoelectric focusing, electrophoretic separation and a great vari-
ety of liquid chromatography techniques, often linked together
to yield two- or three-dimensional separation approaches, and
23.
6 Soloviev
frequently backedup by serious automation. Highly parallel anal-
ysis is often attempted through miniaturisation (12) and the use
of chip-based techniques (13–16) or the Agilent 2100 Bioan-
alyzer (www.chem.agilent.com). The inherent heterogeneity of
the proteins’ and to a lesser degree peptides’ physical properties
which underlies all of the above separation options is, at the same
time, the inherent problem of any highly parallel protein analy-
sis. A single universal system suitable for extraction and separa-
tion (let alone functional analysis) of all classes of proteins is yet
to be reported. Unlike proteins, the peptides are often less het-
erogeneous in their physico-chemical properties and therefore the
complete peptidomic analysis of samples, tissues and in some cases
whole organisms is more straightforward than proteomic analysis.
In addition to physical methods of analysis and separation,
chemical biology offers a number of other approaches, which rely
directly or indirectly on chemical modifications and separation
principles based on chemical properties of proteins and peptides.
Chemical modification of the side chains of proteins and peptides
was first reported many decades ago (see 17 for a review) and
has been used widely since for protein modifications, labelling
and cross-linking, but not so widely for protein separations – the
latter because of the issues related to the availability and surface
exposure of the reactive groups. Unlike proteins, peptides offer
a unique chance to apply chemical selection techniques because
of the lack of complex secondary structure and virtually com-
plete exposure to solvent of all of the reactive groups. A number
of reports utilising chemical biology approach to peptide sepa-
ration and analysis have been published more recently. In most
cases these describe various group-specific labelling procedures,
often linked to peptide quantification (18, 19) as well as chemical
depletion approaches (17, 20). Among the other “omics” tech-
nologies and approaches, “peptidomics” is the most comparable
to “Proteomics” and although the terms are not synonymous, the
underlying techniques and approaches are almost identical. For
example, MS, a cornerstone of modern proteomics, in most cases
actually analyses peptides (obtained through proteolytic digestion
of proteins) or their fragments (obtained through e.g. CID), not
proteins. The difference between the terms “peptidomics” and
the “proteomics” is therefore blurred, especially if “methods” are
being considered.
5. Peptidomics:
The Targets
Target-wise, the peptidomics research is often focused, although
not always, on studying peptides formed in vivo by proteolysis of
specialised or non-specialised precursor proteins (often bioactive
24.
Peptidomics: Divide etImpera 7
peptides), rather than “artificially” or in vitro-produced peptides.
The range of biological activities displayed by naturally occur-
ring peptides is truly remarkable; it ranges from toxins that can
paralyse or kill to peptides that have the ability to heal. The ven-
oms of arthropods such as spiders and scorpions, as well as other
species such as cone snails, comprise a vast number of neuromod-
ulatory peptides that are capable of serious harm, but also serve
as a highly useful point to discover new drugs such as painkillers
(21). The identification and functional characterisation of pep-
tides from all species including humans is crucial in the discovery
of novel biomarkers and drug targets, and may yield novel ther-
apeutic agents such as peptide-based vaccine Glatiramer acetate
(GA) (Copaxone) used for the treatment of relapsing and remit-
ting cases of multiple sclerosis (22). The suitability of peptides
as biomarkers stems from the fact that they are present in all
body fluids, cells and tissues (23), and many approaches focus
on identifying them from such samples (24, 25). Peptides also
play crucial roles in innate and adaptive immune responses by
forming complexes with MHC-I, MHC-II and T cells where they
stimulate defensive immune responses (26–28). The importance
of peptides in cell-to-cell communication underpins the impor-
tance of peptidomics in understanding multiple pathologies that
result from these communication processes going wrong. The
peptide content of biological fluids, such as urine for example,
can be used to produce a complete peptidomic fingerprint of
an individual’s health (29). The evolutionary evidence to sup-
port the importance of peptides in such widespread biological
roles is evident when one examines the conservation of peptide
families across species. The Tachykinin peptides for example, the
largest known neuropeptide family, are found in vertebrates, pro-
tochordates and invertebrates (30). On the other end of the scale,
even primitive microorganisms rely on peptide signalling, such as
for example bacterial quorum sensing (31, 32) and yeast mating
factors (33).
The following chapters provide a comprehensive guide to
peptidomics methods and applications, spanning a range of
species from bacteria to man and covering a wide range of relevant
methods from basic biochemistry techniques to in silico tools and
protocols.
References
1. Chervet, J.P., Ursem, M., and Salzmann,
J.B. (1996) Instrumental requirements for
nanoscale liquid chromatography. Anal.
Chem. 68, 1507–1512.
2. Quadroni, M. and James, P. (1999) Pro-
teomics and automation. Electrophoresis 20,
664–677.
3. Schrader, M. and Schulz-Knappe, P.
(2001) Peptidomics technologies for
human body fluids. Trends Biotechnol. 19,
S55–S60.
4. Verhaert, P., Vandesande, F., and De Loof,
A. (1999) Automated analysis of the pep-
tidome. No longer science fiction. In: 2nd
25.
8 Soloviev
International Seminaron the Enabling Role
of MS in Manchester.
5. Verhaert, P., Uttenweiler-Joseph, S., de Vries,
M., Loboda, A., Ens, W., and Standing,
K.G. (2001) Matrix-assisted laser desorp-
tion/ionization quadrupole time-of-flight
mass spectrometry: an elegant tool for pep-
tidomics. Proteomics 1, 118–131.
6. Schulz-Knappe, P., Zucht, H.D., Heine, G.,
Jürgens, M., Hess, R., and Schrader, M.
(2001) Peptidomics: the comprehensive
analysis of peptides in complex biological
mixtures. Comb. Chem. High Throughput
Screen 4, 207–217.
7. Clynen, E., Baggerman, G., Veelaert,
D., Cerstiaens, A., Van der Horst, D.,
Harthoorn, L., Derua, R., Waelkens, E.,
De Loof, A., and Schoofs, L. (2001) Pep-
tidomics of the pars intercerebralis–corpus
cardiacum complex of the migratory locust,
Locusta migratoria. Eur. J. Biochem. 268,
1929–1939.
8. Scrivener, E., Barry, R., Platt, A., Calvert, R.,
Masih, G., Hextall, P., Soloviev, M., and Ter-
rett, J. (2003) Peptidomics: a new approach
to affinity protein microarrays. Proteomics 3,
122–128.
9. Barry, R., Diggle, T., Terrett, J., and
Soloviev, M. (2003) Competitive assay for-
mats for high-throughput affinity arrays. J.
Biomol. Screen. 8, 257–263.
10. Barry, R. and Soloviev, M. (2004) Quantita-
tive protein profiling using antibody arrays.
Proteomics 4, 3717–3726.
11. Community Trade Mark No. 001274646;
http://oami.europa.eu
12. Marko-Varga, G., Nilsson, J., and Laurell,
T. (2003) New directions of miniaturization
within the proteomics research area. Elec-
trophoresis 24, 3521–3532.
13. Hoa, X.D., Kirk, A.G., and Tabrizian, M.
(2007) Towards integrated and sensitive sur-
face plasmon resonance biosensors: a review
of recent progress. Biosens. Bioelectron. 23,
151–160.
14. Kurosawa, S., Aizawa, H., Tozuka, M.,
Nakamura, M., and Park, J.W. (2003)
Immunosensors using a quartz crys-
tal microbalance. Meas. Sci. Technol. 14,
1882–1887.
15. Lion, N., Rohner, T.C., Dayon, L., Arnaud,
I.L., Damoc, E., Youhnovski, N., Wu,
Z.Y., Roussel, C., Josserand, J., Jensen,
H., Rossier, J.S., Przybylski, M., and
Girault, H.H. (2003) Microfluidic sys-
tems in proteomics. Electrophoresis 24,
3533–3562.
16. Lion, N., Reymond, F., Girault, H.H.,
and Rossier, J.S. (2004) Why the move
to microfluidics for protein analysis?. Curr.
Opin. Biotechnol. 15, 31–37.
17. Soloviev, M. and Finch, P. (2005) Pep-
tidomics, current status. J. Chromatogr. B
Analyt. Technol. Biomed. Life Sci. 815,
11–24.
18. Gygi, S.P., Rist, B., Gerber, S.A., Turecek,
F., Gelb, M.H., and Aebersold, R. (1999)
Quantitative analysis of complex protein mix-
tures using isotope-coded affinity tags. Nat.
Biotechnol. 17, 994–999.
19. DeSouza, L., Diehl, G., Rodrigues, M.J.,
Guo, J., Romaschin, A.D., Colgan, T.J., and
Siu, K.W. (2005) Search for cancer markers
from endometrial tissues using differentially
labeled tags iTRAQ and cICAT with multi-
dimensional liquid chromatography and tan-
dem mass spectrometry. J. Proteome Res. 4,
377–386.
20. Soloviev, M., Barry, R., Scrivener, E.,
and Terrett, J. (2003) Combinatorial pep-
tidomics: a generic approach for protein
expression profiling. J. Nanobiotechnol. 1, 4.
21. Rash, L.D. and Hodgson, W.C. (2002) Phar-
macology and biochemistry of spider ven-
oms. Toxicon 40, 225–254.
22. Perumal, J., Filippi, M., Ford, C., John-
son, K., Lisak, R., Metz, L., Tselis, A.,
Tullman, M., and Khan, O. (2006) Glati-
ramer acetate therapy for multiple sclerosis: a
review. Expert Opin. Drug Metab. Toxicol. 2,
1019–1029.
23. Adermann, K., John, H., Ständker, L., and
Forssmann, W.G. (2004) Exploiting natu-
ral peptide diversity: novel research tools
and drug leads. Curr. Opin. Biotechnol. 15,
599–606.
24. Zimmerman, L.J., Wernke, G.R., Caprioli,
R.M., and Liebler, D.C. (2005) Identifica-
tion of protein fragments as pattern features
in MALDI-MS analyses of serum. J. Proteome
Res. 4, 1672–1680.
25. Vidal, B.C., Bonventre, J.V., and I-Hong
Hsu, S. (2005) Towards the application of
proteomics in renal disease diagnosis. Clin.
Sci. (Lond). 109, 421–430.
26. Desjardins, M., Houde, M., and Gagnon, E.
(2005) Phagocytosis: the convoluted way
from nutrition to adaptive immunity.
Immunol. Rev. 207, 158–165.
27. Cresswell, P., Ackerman, A.L., Giodini, A.,
Peaper, D.R., and Wearsch, P.A. (2005)
Mechanisms of MHC class I-restricted
antigen processing and cross-presentation.
Immunol. Rev. 207, 145–157.
28. Van der Merwe, P.A. and Davis, S.J. (2003)
Molecular interactions mediating T cell anti-
gen recognition. Annu. Rev. Immunol. 21,
659–684.
26.
Peptidomics: Divide etImpera 9
29. Metzger, J., Schanstra, J.P., and Mis-
chak, H. (2009) Capillary electrophoresis-
mass spectrometry in urinary proteome
analysis: current applications and future
developments. Anal. Bioanal. Chem. 393,
1431–1442.
30. Severini, C., Improta, G., Falconieri-
Erspamer, G., Salvadori, S., and Erspamer, V.
(2002) The tachykinin peptide family. Phar-
macol. Rev. 54, 285–322.
31. Miller, M.B. and Bassler, B.L. (2001) Quo-
rum sensing in bacteria. Annu. Rev. Micro-
biol. 55, 165–199.
32. Gibbs, R.A. (2005) Trp modification signals
a quorum. Nat. Chem. Biol. 1, 7–8.
33. Kalkum, M., Lyon, G.J., and Chait, B.T.
(2003) Detection of secreted peptides by
using hypothesis-driven multistage mass
spectrometry. Proc. Natl. Acad. Sci. USA.
100, 2795–2800.
14 Adkins etal.
plasminogen), and nutrient cycling. Despite the critical roles
of protein degradation in biological processes, there have been
surprisingly few systematic global analyses of protein degrada-
tion; the majority of studies that have been performed focus
on eukaryotic systems. Specific protein degradation processes are
very highly regulated in bacteria and determined by environ-
mental conditions. Selective degradation of proteins followed by
cannibalization of the released amino acids is the most efficient
process for bacterial adaptation to changing metabolic require-
ments (1, 2). Indeed, the ability of a pathogen to survive in the
host and exploit new resources is an essential virulence trait.
The development of novel antibiotics against bacterial patho-
gens represents just a single discipline that can benefit from the
elucidation of selective protein degradation processes. Recently
our group developed an LC–MS/MS-based approach to glob-
ally profile a sub-set of peptides in a biological sample. Peptides,
defined here, are short chains of amino acids linked via pep-
tide bonds and are typically composed of fewer than 100 amino
acids. The source of peptides in a biological system may result
from short genes or through targeted degradation of proteins.
Most of the peptides observed in this recent study were found to
be the products of protein degradation (3); regardless of source
we refer here to this naturally occurring peptide fraction as the
“peptidome”.
Interestingly, nearly 2% of the 4550 predicted proteins in
S. typhimurium are annotated as being involved in protein degra-
dation. Importantly, nearly all of these proteolytic proteins were
identified in an early analysis of the S. typhimurium proteome,
indicating that there is an upregulation of these functions under
some of the growth conditions studied (4). The following is a
step-by-step description of the sample preparation and analytical
procedures that were used in determining the Salmonella pep-
tidome. In addition, a discussion of the data analysis concerns
that are unique to analyzing peptidomics samples is included.
2. Materials
Unless stated otherwise, Materials were obtained from Sigma
Aldrich, St. Louis, MO.
2.1. Cell Growth and
Isolation
Cellgro Dulbecco’s Phosphate Buffered Saline (Mediatech,
Mannasas, VA).
2.2. Lysis/Peptide
Extraction Reagents
1. Water purified using a NANOpure R
or equivalent system
(≥ 18 M×cm, Barnstead International, Dubuque, Iowa).
30.
Comparative Peptidomics ofSalmonella 15
2. Ammonium bicarbonate, isopropanol, and methanol (Sigma
Aldrich).
3. Protease inhibitor cocktail formulated for use with bacterial
cell extracts (Cat. No. P8465, Sigma)
4. 0.1 mm zirconia/silica beads (BioSpec Products,
Bartlesville, OK) were used for cell lysis.
5. 10–20% Tris-Tricine Ready Gels R
(Bio-Rad, Hercules, CA)
and GelCode R
Blue reagent from Pierce for SDS-PAGE
analyses.
6. OMIX R
C-18 tips (100 L) (Varian, Inc, Palo Alto, CA) for
sample solid-phase extraction (SPE) clean-up prior to MS
analysis.
7. SpeedVac (Thermo Fisher Scientific, Waltham, MA) to con-
centrate samples.
2.3. Liquid
Chromatography–
Mass
Spectrometry/Mass
Spectrometry
1. Ion trap mass spectrometers (LTQ, Thermo Fisher Scien-
tific, San Jose, CA)
2. Water purified using a NANOpure R
or equivalent system
(≥ 18 M×cm)
3. Mobile phase A: Degassed 0.2% acetic acid, 0.05% trifluo-
roacetic acid in water (Sigma Aldrich)
4. Mobile phase B: Degassed 0.1% trifluoroacetic acid in 90%
acetonitrile (ACN), 10% water (Sigma Aldrich)
5. 5-m Jupiter C18 stationary phase (Phenomenex, Torrence,
CA) packed into 60-cm (360 m o.d. X 150 m i.d.)
fused silica capillary tubing (Polymicro Technologies Inc.,
Phoenix, AZ)
6. Liquid chromatography system is described elsewhere by
Livesay et al. (15)
2.4. Liquid
Chromatography–
High-Resolution
Mass Spectrometry
1. Fourier transform ion cyclotron resonance (FTICR) mass
spectrometer, either a custom-built 11 T instrument or 9.4
T instrument (Bruker Daltonics, Billerica, MA).
2. See Section 2.3 for details on mobile and stationary-phase
materials.
2.5. LC–MS/MS Data
Analyses
1. SEQUEST R
version [TurboSEQUEST R
(cluster) v.27 (rev.
12), Thermo Fisher Corp.]
.
2.6. Proteomics 1. RapiGestTM
(Waters, Milford, MA) is a surfactant to aid in
the solubilization and trypsin digestion of proteins.
2. Trypsin for protein digestion (Promega, Madison, WI)
31.
16 Adkins etal.
3. Bicinchoninic Acid (BCA) Protein Assay kit (Pierce, Rock-
ford, IL) for quantitation of peptides
2.7. Data
Visualization and
Cluster Analysis
1. DAnTE, freely available software for comparative anal-
ysis of proteomics data available at http://omics.pnl.
gov/software/
2. MultiExperiment Viewer (MEV) is also freely available and
designed for use in microarray experiments, but can be par-
ticularly useful for proteomics data visualization and cluster-
ing and is available at http://www.tm4.org/mev.html.
3. Methods
3.1. Culturing
Conditions
The culturing conditions of the bacteria are not the focus of
this review but are summarized here. The primary difference
between the culture conditions used is that various forms of
stresses, some relevant to pathogenesis, were compared relative
to a rich growth medium at middle logarithmic growth phase.
Wild-type S. typhimurium strains 14028 and LT2 were grown
to mid-logarithmic (Log) and stationary (Stat) phases in Luria-
Bertani (LB) broth and harvested for analysis. Two other cell
growth conditions were used that differed only in the pre-growth
conditions. In one, the bacteria were grown to stationary phase
in LB, the bacteria were isolated, washed, and then grown in
magnesium-minimal acidic medium (Shock); in the other, the
bacteria were diluted 1:100 and grown in acidic minimal media
overnight (Dilu). All cultures were harvested following stan-
dard batch culture techniques as outlined (see references (3–5)
for more detail of culture methods). Aliquots of cell cultures
(corresponding to 0.15 g cell pellets) were pelleted, washed in
PBS, flash frozen with liquid N2, and used as needed to prepare
samples.
3.2. Sampling
Preparation and
Peptide Extraction
The procedures outlined here are specific to samples that require
Biosafety Level 2 (BSL2) containment and treatment. Many of
the precautions (e.g., O-ring sealed cryovials, cooling following
vortexing) are to prevent aerosolization of unlysed pathogenic
organisms. When developing these protocols, we lysed cells in the
presence of a protease inhibitor cocktail formulated for use with
bacterial cell extracts. However, we did not evaluate correspond-
ing analyses without the protease inhibitor cocktail (see Note 1).
The listing of class-specific chemical inhibitors of proteases found
in the excellent review by Overall and Blobel (6) may be con-
sulted if protease inhibition is desired. In our previous work, we
32.
Comparative Peptidomics ofSalmonella 17
performed tests to mimic poor sample handling (incubation at
22◦
C for 20 min without inhibitors) and compared these results
to those obtained when the samples were prepared at ∼7◦
C with a
cooling block (normal handling temperatures with inhibitors) (see
Note 2). We found no significant variation in the peptides iden-
tified. The procedure below is based on an isopropanol extrac-
tion that causes larger proteins to precipitate while endogenous
peptides are maintained in solution. Different concentrations of
isopropanol were tested and it was determined that a ratio of
3:2 resulted in the best recovery of endogenous peptides from S.
typhimurium. This may not hold true for all biological samples.
1. Lysis of bacterial cells is accomplished by first resuspend-
ing the cell pellet in an equivalent volume of 100 mM
NH4HCO3, followed by transfer of the sample to a 2.0-mL
O-ring sealed cryovial. Next, 0.1-mm zirconia/silica beads
are added to half of the volume in the tube, and the tube
is then vortexed for 30 s, followed by cooling for 1 min in
a cold-block. Six cycles of vortexing and cooling are per-
formed. The lysate is then removed from the top of the set-
tled beads, and the beads are rinsed five times with buffer.
The lysates and rinses are then pooled separately in micro-
centrifuge tubes (see Note 3).
2. The pooled lysate is centrifuged at 16,000×g for 10 min at
room temperature to pellet insoluble and precipitated pro-
teins. Transfer the supernatant to a new microcentrifuge
tube, and ensure that the entire pellet is left behind. The
supernatant is now considered a cleared lysate. An aliquot of
the cleared lysate can be saved for SDS-PAGE as a reference.
3. Isopropanol is then added to the cleared lysate in an
appropriate ratio (we used 1:1, 3:2, 2:1, or 5:2 (v/v, iso-
propanol:lysate)), and the samples were mixed by vortexing.
Pre-cooling the isopropanol to 4◦
C before adding to the
lysate can assist with precipitation of proteins. The samples
are then incubated at 4◦
C for 15 min, then microcentrifuged
at 16,000×g for 10 min at 4◦
C to remove precipitated
proteins. The resulting supernatants are transferred to new
microcentrifuge tubes and concentrated in a SpeedVac to
∼75 L. Ten-microliter aliquots can be removed at this time
for SDS-PAGE.
4. Peptide concentrations are determined by BCA protein
assay, and SDS-PAGE analyses are performed using 10–20%
Tris-Tricine Ready Gels R
. The Tris-Tricine gels are used
because they are specific for the separation of extremely small
proteins and peptides. Gels are fixed for 30 min in 40%
methanol/10% acetic acid and then stained for 60 min using
GelCode R
Blue reagent.
33.
18 Adkins etal.
5. Prior to MS analysis, the concentrated isopropanol extracts
are cleaned via solid-phase extraction using OMIX C18
pipette tips. These tips are monolithic, rather than particu-
late, and are therefore much easier to use without clogging,
while providing better recovery and reproducibility. Thirty
micrograms of peptide mass from each sample is applied to
a 100-L tip. The directions provided by the manufacturer
are used to condition, wash, and load the samples. Peptides
are eluted from the tips with 80:20 ACN:H2O containing
0.1% TFA. Eluted peptides are concentrated to ∼15 L in a
SpeedVac.
6. Alternatively, samples can be fractionated using strong cation
exchange (SCX) HPLC to minimize sample complexity prior
to each LC–MS/MS analysis, as described previously (7).
Each fractionation is performed using approximately 150 g
(peptide mass) of concentrated isopropanol extract, result-
ing in 25 fractions that are concentrated in a SpeedVac
to dryness. The samples are then reconstituted in 25 mM
NH4HCO3 to a volume appropriate for LC–MS/MS
analysis.
3.3. Liquid
Chromatography–
Mass
Spectrometry/Mass
Spectrometry
Our analytical instrumentation consists of commercially avail-
able platforms [e.g., ion traps (LTQ from ThermoFisher) and
FTICR–MS (BrukerDaltonics)] that are in-house modified to
increase the sensitivity and throughput of the analyses. How-
ever, the below LC–MS(/MS) approaches can be applied at a
reasonable level of quality with more generally available off-the-
shelf instrumentation. LC–MS/MS analyses are useful for mak-
ing identifications and for semi-quantitation based on “spectrum
counting” techniques (4, 8–10). These analyses are also used
to build a database of identified peptides annotated with deter-
mined reversed-phase elution times (11) and calculated masses.
This database (also referred to as a mass and time tag lookup
table) is used with results from the high-resolution MS analyses
(Section 3.4) to increase throughput, perform label-free quanti-
tation, and improve peptide-sampling methods in the MS exper-
iment. This is a simplified description of the accurate mass and
time (AMT)-tag process developed in our laboratory, which has
been extensively discussed elsewhere (12–14).
1. The concentrated C18 SPE eluents from the peptide clean-
up procedure and the SCX fractions are then analyzed
by reversed-phase microcapillary HPLC (15) interfaced
through nanoelectrospray ionization (nanoESI) to an ion
trap mass spectrometer, as described previously (4). Briefly,
the technique used in our laboratory entails gradient elution
of peptides over 100 min using a 360 m OD × 150 m
ID × 65 cm long capillary column packed with 5 m Jupiter
C18 particles.
34.
Comparative Peptidomics ofSalmonella 19
2. For typical “bottom-up” proteomics experiments, in which
the proteins are digested with trypsin, the charge states of
peptides detected during LC–MS/MS are typically +2 and
+3. Detected peptides are then fragmented using collision-
induced dissociation. It should be noted that the pep-
tides detected from the S. typhimurium endogenous pep-
tidome include more +4 and +5 charge states than typically
observed for other sample types. Due to the larger num-
ber of higher charged species, electron transfer dissociation
may be considered for future analyses of the endogenous
peptidome.
3.4. LC–MS Analyses 1. Concentrated C18 SPE eluents are also analyzed in our lab-
oratory by reversed-phase microcapillary HPLC–nanoESI–
FTICR–MS (11.5 T) (16). The same chromatographic plat-
forms are used for LC–MS/MS analyses as is used with the
FTICR–MS, and during analysis of multiple samples to be
compared, the same chromatography column and electro-
spray emitter is preferred. This reduces the number of con-
founding variables during an experiment for downstream
data analysis.
2. The analysis order for an experiment such as this needs to
be addressed to minimize the effects of analysis time and
possibility of carryover from highly abundant peptides. This
is referred to as “randomized block design” and is meant
to remove experimental nuisance factors that can obscure
true differences between samples (see Note 4). These blocks
typically contain one replicate for each experiment and the
order of the analyses within a block is randomized.
3. Peptides from the LC–MS spectra are identified using the
AMT tag approach (14), including any peptides with +4
and +5 charge states. The necessary software tools are pub-
licly available (http://omics.pnl.gov). This approach uses
the calculated mass and the observed normalized elution
time (NET) of each filter-passing peptide identification (see
Section 3.5) from the previous LC–MS/MS analyses to
construct a reference database of AMT tags. Features from
LC–MS analyses (i.e., m/z peaks deconvoluted of isotopic
and charge state effects and then annotated by mass and
NET) are matched (13) to AMT tags to identify peptides
in a manner that results in roughly 5% false-positive identifi-
cations. For each protein, the sum of its peptide peak areas
(NET vs. peak height) is used as a measure of the abundance
of its fragments within the peptidome.
3.5. LC–MS/MS Data
Analyses
Peptides can be identified using a number of different publicly
available software packages.
35.
20 Adkins etal.
1. In this example, we utilize SEQUEST R
to search the result-
ing MS/MS spectra against the annotated S. typhimurium
FASTA data file of proteins translated from genetic code
provided by the J. Craig Venter Institute – formerly
TIGR (4550 protein sequences, http://www.jcvi.org/)
(17). These analyses used a standard parameter file with a
peptide mass tolerance = 3, fragment ion tolerance = 0, and
no amino acid modifications. Also, these analyses search for
all possible peptide termini (i.e., not limited to only tryptic
termini). Separate SEQUEST R
searches that use the above
FASTA data file but with scrambled amino acid sequences
are performed in parallel to estimate the false discovery rate.
2. SEQUEST R
generally returns multiple peptide identifica-
tions for each MS/MS spectrum and for each parent ion
charge state. Therefore, for each MS/MS spectrum and for
each parent ion charge state, only the peptide identification
with the highest XCorr value (i.e., the “top ranked hit”) is
retained here.
3. Limiting false identification of peptides is an especially
challenging issue for natively produced peptides because
cleavage state (i.e., trypsin cleavage sites) is often used in
making confident identifications. PeptideProphet (18) val-
ues are also not applicable because of a strong bias for
“tryptic” peptides. The estimated percentage of false-
positive peptide identifications can be defined as %FPest. =
100% × (number of scrambled peptide identifications) /
(number of normal peptide identifications) (19). %FPest.
should be calculated for each charge state, XCorr Cutoff
value (the minimum XCorr value requirement, which ranged
from 1.5 to 5 in units of 0.02), and Cn Cutoff value (i.e.,
the minimum Cn value requirement, which ranged from 0
to 0.4 in units of 0.005). In an effort to maximize identifi-
cations, a two-dimensional analysis of the XCorr Cutoff and
Cn Cutoff is used for each parent ion charge state. This
method is different from typical proteomics analyses in that
it does not use a single Cn Cutoff value.
4. The optimal XCorr Cutoff and Cn Cutoff values for each
parent ion charge state (+1 to +5) was determined in our
previous work to be 1.84 and 0.21 (+1), 2.1 and 0.21 (+2),
2.8 and 0.23 (+3), 3.56 and 0.265 (+4), and 4.16 and 0.22
(+5), respectively.
5. A rough measure of the abundance of each parent protein
and its fragments within the peptidome can be attained using
a spectrum counting (i.e., tallying of filter-passing peptide
identifications) approach (20).
36.
Comparative Peptidomics ofSalmonella 21
3.6. Comparison to
Proteomics
Peptidomics data (samples acquired without digestion) should
ideally be compared to proteomics data (samples acquired using
typical bottom-up proteomics approaches including the use of
trypsin) from the same source material. This comparison ensures
that the peptidomics results are interpreted and can be com-
pared with peptides resulting from abundant proteins being non-
specifically degraded. We performed a proteomics analysis with
the same starting sample material to that used in the peptidomics
experiment (4). Briefly, proteins are isolated and digested as
described in the protocol provided by Waters with the modi-
fication of 2.0% TFA rather than using concentrated HCL to
adjust to a pH of 3.0. Acid incubation occurred at 37◦
C for
1 h to fully precipitate the RapiGestTM
surfactant. The samples
are centrifuged in a microcentrifuge at full speed to pellet the
RapiGestTM
and the supernatant is returned to neutral pH with
NH4OH to allow for digested peptide concentration determina-
tion by BCA protein assay.
The resultant peptides are then fractionated using strong
SCX HPLC (7) into 25 fractions. A single unfractionated sam-
ple and the full set of 25 SCX fractions are then analyzed by
reversed-phase LC–MS/MS. MS/MS spectra are searched using
SEQUEST R
and filtered to reduce false-positive peptide identifi-
cations (3, 4, 20).
3.7. Data
Visualization and
Cluster Analysis
The comparative interpretation of the identified proteins and
peptides can present unique challenges. In the case of compar-
ing environmentally induced changes in the S. typhimurium pro-
teome and peptidome, one challenge is that many proteins are
not commonly observed across all conditions. If one generates
a matrix of protein/peptides (rows) by experimental conditions
(column) populated with values of spectral observations or peak
abundance measurements, the unobserved proteins/peptides are
sometimes referred to as “missing data”. The source of an unob-
served species can be the result of either of the following: (1)
its actual absence in a sample, (2) it is present, but below the
detection limit of the mass spectrometer, or (3) the identification
did not pass various quality thresholds used for confident pep-
tide identifications. This results in a less than ideal direct appli-
cation of statistical methods typically used for comparisons of
high-throughput data (microarrays) such as an analysis of variance
(ANOVA). For this reason, we typically try to combine the abun-
dance values for all peptides from a source protein into a single
representative protein abundance for comparison across condi-
tions. This collapsing of peptide abundance to protein abundance
is often referred to by us as “protein roll-up” (see Note 5). These
protein values are then grouped by similar abundance profile
37.
22 Adkins etal.
changes using methods such as a hierarchical clustering, which are
common for microarray analysis comparisons. The comparative
analyses of the peptide and protein abundances are enabled with
the use of data mining tools that offer clustering and heatmap
visualization of the matrix form of the experimental results, e.g.,
DAnTE (21), OmniViz R
(22), or MeV (23). Some considerations
that must be made when analyzing the data are listed below:
Fig. 2.1. Example heat map showing endogenous peptidomics results compared to
global proteomics results. Observations across conditions were scaled using the Z-score
across protein (with black representing a Z-score of 2.5 and white a Z-score of –1.0).
Two selected regions were taken from data found elsewhere. “Stress response factors”,
in this case endogenously occurring peptides, correspond well with the abundance of
the proteins in the proteomics experiments. The “stress turn-over” peptides appear to
be scavenged in the “Dilu” stress condition, and these proteins appear to only be overly
abundant in the rich logarithmic growth condition.
38.
Comparative Peptidomics ofSalmonella 23
1. One of the first decisions is whether to fill arbitrary values
into the unobserved peptide/protein abundances to make
the analysis more amenable to various downstream data anal-
ysis methods typically applied in transcriptional microarray
data analysis, such as ANOVA, principle component anal-
ysis, and/or clustering methods. If the number of spec-
tra observed in a protein are used as a surrogate for an
abundance measurement, filling might include applying the
Fig. 2.2. A demonstration of proteomics results in the context of endogenous peptidomics. Although tryptic digestion
was use in this example, the disappearance of a number of peptides between the stationary (stat) and shock conditions
indicates that the protein is being differentially acted upon by proteases in the cell between the two conditions. This is
especially true when this protein YciF was observed to be particularly abundant in the peptidome in the shock condition
previously (3). As a secondary confirmation, new peptides that were not observed in the stationary condition appear
in the shock condition. New “partially tryptic peptides” also appear and are highlighted with ‘==’ in the figure. The
numbers under the conditions represent the biological replicate of that growth condition.
39.
24 Adkins etal.
minimum number of required peptides for protein identifi-
cation (see Note 6).
2. In both the peptide-centric and protein-centric (using a sin-
gle abundance value for the protein) analysis, the difference
in abundance between the most abundant peptide/protein
versus the least abundant species may range several orders of
magnitude. This large dynamic range of measurements may
lead to difficulty comparing proteins with similar trends in
a set of experiments. To use clustering tools, this dynamic
range must be compensated for by scaling to similar mag-
nitudes for comparison (i.e., a trend that is varied across
2 orders of magnitude should be grouped with other sim-
ilar trends varying across 2 orders of magnitude even if the
most abundant value to least abundant value between pro-
tein is across 6 orders of magnitude). Depending on the
nature of quantitation (spectrum count versus peak area) and
the number of experiments being compared (fewer than six
versus thirty or more), different scaling approaches are pre-
ferred (see Note 7).
3. Once these steps are performed, comparisons between the
experimental samples (both from the undigested native pep-
tidome and the digested proteome) can be performed using
heat maps of the clustered results (Fig. 2.1).
4. Once an endogenous peptidome analysis has been per-
formed, and knowledge of proteins that are subject to native
proteolysis is obtained, it is then possible to extract some
additional information utilizing only a proteomics (i.e.,
trypsin was used) analysis by looking for non-tryptic cleav-
age sites (for an example Fig. 2.2 ).
4. Notes
1. It is reasonable to consider the goals of the experiment, there
may be a specific desire to leave a class of proteases active to
amplify the abundance of the cleaved products.
2. Set cooling block between 6 and 8◦
C, leaving the cooling
blocks in the refrigerator 1 day prior to the experiment. Be
sure to confirm that freezing will not occur by using micro-
centrifuge tubes of ∼100 L of water in the block during
cooling.
3. All microcentrifuge tubes from this point forward should
be siliconized (Fisher 02-681-332) to prevent polymer
contamination, which is detrimental to downstream LC–
MS(/MS) analyses.
40.
Comparative Peptidomics ofSalmonella 25
4. The USA National Institute of Standards and Technol-
ogy maintains an electronic Engineering Statistics Handbook
(http://www.itl.nist.gov/div898/handbook) with a useful
discussion of “Randomized block designs” for experiments.
5. “Protein roll-up” refers to methods that attempt to give
a single value for each protein for quantitative purposes,
even though each protein identification in a bottom-up pro-
teomics experiment typically is based on more than one pep-
tide identification. As of this writing, DAnTE offers multiple
methods for protein roll-up (21).
6. Typically, an identification of a specific protein based on its
tryptic cleavage products requires identification of three sep-
arate tryptic peptides. For native peptidomics this is not real-
istic because there is a high likelihood that only a single
species will be present. Biological conclusions based on sin-
gle peptide identifications should be based on methods with
better relative abundance measurements such as the spectral
peak abundance.
7. For large experiments, a Z-score (24) analysis can be helpful
to visualize significant trends that are further than expected
by a normal distribution. This is also better suited for peak
area-based quantitation where the values are non-integers.
For smaller experiments, dividing each value in a peptide or
protein row by the associated sum, mean, or median of that
entire row can be a useful method to scale the results.
Acknowledgments
This work was supported by the National Institute of Allergy and
Infectious Diseases (NIH/DHHS through interagency agree-
ment Y1-AI-4894-01 and Y1-AI-8401-01). The authors also
acknowledge the US Department of Energy Office of Biological
and Environmental Research and National Center for Research
Resources (RR18522) for the development of the instrumen-
tal capabilities used for the research. Significant portions of this
research were performed in the Environmental Molecular Sci-
ences Laboratory, a US Department of Energy (DOE) national
scientific user facility located at the Pacific Northwest National
Laboratory (PNNL) in Richland, Washington. PNNL is a multi-
program national laboratory operated by Battelle Memorial Insti-
tute for the DOE under Contract No. DE-AC05-76RLO-1830.
41.
26 Adkins etal.
References
1. El-Sharoud, W.M. and Niven, G.W. (2007)
The influence of ribosome modulation fac-
tor on the survival of stationary-phase
Escherichia coli during acid stress. Microbiol-
ogy 153, 247–253.
2. Wada, A., Mikkola, R., Kurland, C.G., and
Ishihama, A. (2000) Growth phase-coupled
changes of the ribosome profile in natural iso-
lates and laboratory strains of Escherichia coli.
J. Bacteriol. 182, 2893–2899.
3. Manes, N.P., Gustin, J.K., Rue, J., Mot-
taz, H.M., Purvine, S.O., Norbeck, A.D.,
Monroe, M.E., Zimmer, J.S., Metz, T.O.,
Adkins, J.N., Smith, R.D., and Heffron,
F. (2007) Targeted protein degradation by
Salmonella under phagosome-mimicking cul-
ture conditions investigated using compar-
ative peptidomics. Mol. Cell Proteomics 6,
717–727.
4. Adkins, J.N., Mottaz, H.M., Norbeck, A.D.,
Gustin, J.K., Rue, J., Clauss, T.R., Purvine,
S.O., Rodland, K., Heffron, F., and Smith,
R.D. (2006) Analysis of the Salmonella
typhimurium proteome through environ-
mental response toward infectious condi-
tions. Mol. Cell Proteomics 5, 1450–1461.
5. Beuzon, C.R., Meresse, S., Unsworth, K.E.,
Ruiz-Albert, J., Garvis, S., Waterman, S.R.,
Ryder, T.A., Boucrot, E., and Holden, D.W.
(2000) Salmonella maintains the integrity of
its intracellular vacuole through the action of
SifA. EMBO J. 19, 3235–3249.
6. Overall, C.M. and Blobel, C.P. (2007) In
search of partners: linking extracellular pro-
teases to substrates. Nat. Rev. Mol. Cell Biol.
8, 245–257.
7. Adkins, J.N., Varnum, S.M., Auberry, K.J.,
Moore, R.J., Angell, N.H., Smith, R.D.,
Springer, D.L., and Pounds, J.G. (2002)
Toward a human blood serum proteome:
analysis by multidimensional separation cou-
pled with mass spectrometry. Mol. Cell Pro-
teomics 1, 947–955.
8. Gao, J., Friedrichs, M.S., Dongre, A.R.,
and Opiteck, G.J. (2005) Guidelines for
the routine application of the peptide hits
technique. J. Am. Soc. Mass Spectrom. 16,
1231–1238.
9. Gao, J., Opiteck, G.J., Friedrichs, M.S., Don-
gre, A.R., and Hefta, S.A. (2003) Changes
in the protein expression of yeast as a func-
tion of carbon source. J. Proteome Res. 2,
643–649.
10. Ishihama, Y., Oda, Y., Tabata, T., Sato, T.,
Nagasu, T., Rappsilber, J., and Mann, M.
(2005) Exponentially modified protein abun-
dance index (emPAI) for estimation of abso-
lute protein amount in proteomics by the
number of sequenced peptides per protein.
Mol. Cell Proteomics 4, 1265–1272.
11. Monroe, M.E., Shaw, J.L., Daly, D.S.,
Adkins, J.N., and Smith, R.D. (2008)
MASIC: A software program for fast quanti-
tation and flexible visualization of chromato-
graphic profiles from detected LC–MS(/MS)
features. Comput. Biol. Chem. 32(3),
215–217.
12. Norbeck, A.D., Monroe, M.E., Adkins, J.N.,
Anderson, K.K., Daly, D.S., and Smith, R.D.
(2005) The utility of accurate mass and LC
elution time information in the analysis of
complex proteomes. J. Am. Soc. Mass Spec-
trom. 16, 1239–1249.
13. Monroe, M.E., Tolic, N., Jaitly, N.,
Shaw, J.L., Adkins, J.N., and Smith, R.D.
(2007) VIPER: an advanced software pack-
age to support high-throughput LC–MS
peptide identification. Bioinformatics 23,
2021–2023.
14. Zimmer, J.S., Monroe, M.E., Qian, W.J., and
Smith, R.D. (2006) Advances in proteomics
data analysis and display using an accurate
mass and time tag approach. Mass Spectrom.
Rev. 25, 450–482.
15. Livesay, E.A., Tang, K., Taylor, B.K.,
Buschbach, M.A., Hopkins, D.F., LaMarche,
B.L., Zhao, R., Shen, Y., Orton, D.J.,
Moore, R.J., Kelly, R.T., Udseth, H.R., and
Smith, R.D. (2008) Fully automated four-
column capillary LC–MS system for max-
imizing throughput in proteomic analyses.
Anal. Chem. 80, 294–302.
16. Gorshkov, M.V., Pasa Tolic, L., Udseth,
H.R., Anderson, G.A., Huang, B.M., Bruce,
J.E., Prior, D.C., Hofstadler, S.A., Tang, L.,
Chen, L.Z., Willett, J.A., Rockwood, A.L.,
Sherman, M.S., and Smith, R.D. (1998)
Electrospray ionization-Fourier transform
ion cyclotron resonance mass spectrometry
at 11.5 tesla: instrument design and ini-
tial results. J. Am. Soc. Mass Spectrom. 9,
692–700.
17. Peterson, J.D., Umayam, L.A., Dickinson,
T., Hickey, E.K., and White, O. (2001) The
comprehensive microbial resource. Nucleic
Acids Res. 29, 123–125.
18. Keller, A., Nesvizhskii, A.I., Kolker, E., and
Aebersold, R. (2002) Empirical statistical
model to estimate the accuracy of peptide
identifications made by MS/MS and database
search. Anal. Chem. 74, 5383–5392.
19. Qian, W.J., Liu, T., Monroe, M.E., Strittmat-
ter, E.F., Jacobs, J.M., Kangas, L.J.,
Petritis, K., Camp, D.G., 2nd, and Smith,
42.
Comparative Peptidomics ofSalmonella 27
R.D. (2005) Probability-based evaluation of
peptide and protein identifications from tan-
dem mass spectrometry and SEQUEST anal-
ysis: the human proteome. J. Proteome Res. 4,
53–62.
20. Qian, W.J., Jacobs, J.M., Camp, D.G.,
2nd, Monroe, M.E., Moore, R.J., Gritsenko,
M.A., Calvano, S.E., Lowry, S.F., Xiao, W.,
Moldawer, L.L., Davis, R.W., Tompkins,
R.G., and Smith, R.D. (2005) Comparative
proteome analyses of human plasma follow-
ing in vivo lipopolysaccharide administration
using multidimensional separations coupled
with tandem mass spectrometry. Proteomics
5, 572–584.
21. Polpitiya, A.D., Qian, W.J., Jaitly, N., Petyuk,
V.A., Adkins, J.N., Camp, D.G., 2nd,
Anderson, G.A., and Smith, R.D. (2008)
DAnTE: a statistical tool for quantitative
analysis of -omics data. Bioinformatics 24,
1556–1558.
22. Saffer, J.D., Burnett, V.L., Chen, G., and
van der Spek, P. (2004) Visual analytics in
the pharmaceutical industry. IEEE Comput.
Graph Appl. 24, 10–15.
23. Saeed, A.I., Sharov, V., White, J., Li, J.,
Liang, W., Bhagabati, N., Braisted, J., Klapa,
M., Currier, T., Thiagarajan, M., Sturn,
A., Snuffin, M., Rezantsev, A., Popov, D.,
Ryltsov, A., Kostukovich, E., Borisovsky, I.,
Liu, Z., Vinsavich, A., Trush, V., and Quack-
enbush, J. (2003) TM4: a free, open-source
system for microarray data management and
analysis. Biotechniques 34, 374–378.
24. Adkins, J.N., Monroe, M.E., Auberry, K.J.,
Shen, Y., Jacobs, J.M., Camp, D.G., 2nd,
Vitzthum, F., Rodland, K.D., Zangar, R.C.,
Smith, R.D., and Pounds, J.G. (2005) A
proteomic study of the HUPO Plasma Pro-
teome Project’s pilot samples using an accu-
rate mass and time tag strategy. Proteomics 5,
3454–3466.
30 Husson etal.
grow and maintain. Since its introduction as a model system in
the 1960s, C. elegans was used widely in many research labora-
tories due to the ease of handling and the well-defined anatomy.
C. elegans contains exactly 959 cells that are ordered in sets of
fully differentiated tissues. There are two sexes. Hermaphrodites
can self-fertilize or mate with males in order to produce over
300 offspring. Although hermaphrodites are the most common
sex in nature, mating with males will yield a 50% male progeny.
In the laboratory, self-fertilization of the hermaphrodites or
crossing with males can easily be manipulated for genetic studies.
In addition, C. elegans has a short life cycle. It takes about 3–4
days from egg to egg and it goes through four larval stages
(L1–L4) until reaching adulthood. A developmentally arrested
“dauer” larva can be formed under conditions of starvation or
overcrowding. These thinner dauers have a relative impermeable
cuticle, are non-feeding, and can survive for months, in contrast
to the average life span of around 2–3 weeks under standard
conditions.
A sophisticated knowledge infrastructure has been devel-
oped, with many research methods and protocols that are
widely shared in the “worm-community.” Most information
can be found in the easily accessible database “WormBase”
at (http://www.wormbase.org). The “WormBook” (http://
www.wormbook.org) can be considered as the open-access col-
lection of peer-reviewed chapters that covers all kinds of differ-
ent topics and protocols related to C. elegans. This nematode
is also perfectly suited for light microscopy due to its trans-
parency. For high-end visual analysis of C. elegans, the micro-
scope has to be equipped with differential interference contrast
(DIC; Nomarski) optics for obtaining 3D-like view of the tis-
sues. This way, individual neurons can be observed and recog-
nized. As an example, a DIC image of an L1 larva is shown in
Fig. 3.1. Detailed DIC and electron microscopic images are avail-
able on “WormAtlas” (http://www.wormatlas.org), together
with a plethora of detailed schematic representations. C. elegans
was the first multicellular organism to have its genome fully
sequenced (1). Its genome (about 100 Mb) encodes for over
20,000 proteins and its size is about 1/30th of that of a human.
The awarding of the Nobel Prize to the three “worm-pioneers”
Sydney Brenner, Robert Horvitz, and John Sulston in 2002 for
their discoveries concerning genetic regulation of organ devel-
opment and programmed cell death, to Andrew Fire and Craig
Mello in 2006 for their discovery of RNA interference in this
nematode, and to Martin Chalfie in 2008 for the discovery and
development of the green fluorescent protein (GFP), emphasizes
the great potential of this tiny nematode of being a model organ-
ism, just like the fruit fly Drosophila melanogaster, the marine snail
Aplysia californica, and the mouse Mus musculus.
45.
Peptidomics of C.elegans 31
Fig. 3.1. Differential interference contrast image of a C. elegans L1 larva. The first larval stage of the nematode C. elegans
is shown. This picture was taken using an Axio Observer Z1 instrument (Zeiss) equipped with differential interference
contrast (DIC) or Nomarski optics to allow a clear 3D-like structure of individual neurons.
1.2. Peptidomics of
C. elegans
(Neuro)peptides are small messenger molecules that are derived
from larger precursor proteins by the highly controlled action
of processing enzymes. These biologically active peptides can
be found in all metazoan species where they orchestrate a
wide variety of physiological processes. The knowledge of the
primary amino acid sequence of the neuropeptidergic signaling
molecules is absolutely necessary to understand their function
and interactions with G-protein-coupled receptors. Three classes
of neuropeptide-encoding genes have been predicted from the
genomic data of C. elegans. Initially, 24 FMRFamide-like peptide
(flp) genes have been found by searching cDNA libraries and
genomic sequences (2–4); more flp genes were identified by
mining the EST data (5) (see Table 3.1). By searching the
C. elegans genome for predicted proteins with the structural hall-
marks of neuropeptide precursors, 32 so-called neuropeptide-like
protein (nlp) genes have been identified (6) (see Table 3.2).
These neuropeptide preproproteins all contain peptides without
the RFamide motif, but display sequence homology with other
36 Husson etal.
invertebrate neuropeptides. Finally, a systematic search for genes
encoding members of the insulin superfamily revealed the pres-
ence of 40 insulin-like genes (7). Neuropeptidergic signaling
in the nematode C. elegans has recently been reviewed (8).
Based on such sequence information alone, one cannot deduce
whether all the predicted peptides are actually expressed and
properly processed. Therefore, each such neuropeptide needs to
be purified and characterized biochemically. In the past, bio-
chemical purification and elucidation of neuropeptide sequences
required multiple chromatographic separation steps to purify an
individual biologically active peptide. This approach appeared
to be problematic, especially for small-sized animals, such as
C. elegans. Previously, only 12 neuropeptides of C. elegans
could be biochemically isolated and identified using Edman
degradation analysis or gas-phase sequencing (9–14). Recently
we set out to systematically search for and characterize neu-
ropeptides of C. elegans using high-throughput peptidomics tech-
niques. A peptidomics approach aims to identify endogenous
(neuro)peptides using liquid chromatography and mass spec-
trometry. We aimed to elucidate which peptides were actually
present in the nematode and to identify any post-translational
modifications, which are often required for the peptide’s bioactiv-
ity. We successfully analyzed the peptidome of C. elegans (15, 16),
and C. briggsae (17), while the Ascaris suum peptidome has been
explored by others (18, 19). Differential peptidomics techniques
allowed us to characterize the neuropeptide precursor processing
enzymes EGL-3 (20, 21) and EGL-21 (22) and the neuroen-
docrine chaperone protein 7B2 (23). In this chapter we mainly
focus on the basic techniques and methods required to culture
the nematodes and to perform the sample preparation. Then, dif-
ferent technologies that can be used in peptidomic research are
described and a short overview is provided of the instrumenta-
tion needed.
2. Materials
2.1. C. elegans
Culture
1. C. elegans strains can be ordered from the Caenorhab-
ditis Genetics Center (CGC, http://www.cbs.umn.edu/
CGC/), which is supported by the National Institutes of
Health–National Center for Research Resources. This cen-
ter collects, maintains, and distributes all kinds of C. elegans
strains at a $7 fee per strain in the case of academic/non-
profit organizations or a $100 fee per strain for commer-
cial organizations, in addition to the annual fee of $25.
C. elegans N2 (Bristol) is referred to as the wild-type
+
+ +
+
+ ++
+
+ +
+ –
+ + –
+
Acad. 69: 1330. D. 23, ’05. 670w.
“He tells his tale modestly and sincerely, without striving to put
his best foot foremost and without any trace of bitterness towards
opponents.”
Cath. World. 83: 107. Ap. ’06. 990w.
Critic. 48: 380. Ap. ’06. 110w.
“Mr. O’Brien’s book takes rank with Mr. Justin McCarthy’s
politico-autobiographical reminiscences. While its scope is
narrower, its vividness is more intense. The author at times writes,
as it were, with his very heart’s blood; and thus writing he cannot
fail to command a reading.” Percy F. Bicknell.
Dial. 40: 37. Ja. 16, ’06. 1910w.
“Lacks the historic value which attaches to Mr. Michael Davitt’s
‘Fall of feudalism.’”
Ind. 60: 930. Ap. 19, ’06. 380w.
“They constitute in fact a human document wherein may be read
not merely the personal characteristics of their author, but the
predominating traits of his countrymen.”
Lit. D. 32: 453. Mr. 24, ’06. 470w.
“Unfortunately, too. Mr. O’Brien is throughout careless about
dates, and the index is little help to anybody who wishes to follow
in a serious spirit a rambling and disjointed story.”
Lond. Times. 4: 439. D. 15, ’05. 1710w.
“The book will be read with interest by all who have lived
through those days and who are interested in Irish affairs.”
Nation. 82: 120. F. 8, ’06. 320w.
Pub. Opin. 40: 59. Ja. 13, ’06. 450w.
R. of Rs. 33: 115. Ja. ’06. 110w.
53.
+
+ –
“So longas Mr. O’Brien keeps to personal touches, and to his
delightful Irish humor and sentiment, we find him a very pleasant
storyteller.”
Sat. R. 101: 493. Ap. 21, ’06. 1550w.
“Both in tone and style the book is a pleasant one, and every one
who wishes to form a clear idea of the Nationalist case against the
British Government from 1865 to 1883 should make a point of
studying it though unquestionably it requires careful checking
from other sources.”
Spec. 96: 302. F. 24, ’06. 1640w.
Ogden, Horatio Nelson. Child in the church. 25c. Meth. bk.
The order for the administration of baptism to infants according
to the discipline and usage of the Methodist Episcopal church,
together with the duties of parents, the apostles’ creed, and the
catechism, make up this booklet, which has as a frontispiece a
blank certificate of baptism. The volume forms a dainty baptismal
gift.
O’Higgins, Harvey Jerrold. Don-a-dreams: a story of love and
youth. †$1.50. Century.
The practical, everyday world seems a very sordid thing to one
who follows the story of this dreamer of dreams, who from nursery
make-believes and childish day dreams passes into a youth of
ideals and is left in his early manhood still a visionary but with
many dreams come true. With a skilful touch Don is put before us;
misunderstood by a commonplace father, an acknowledged failure
at a practical college course, a failure in New York where he tries to
make a living as a super at a second class theatre or at anything
else, he suddenly blossoms into a recognized genius as a writer of
plays. And through years of struggle, from earliest childhood, his
love for Margaret, his ideal, burns like a white flame, and in return
she loves him, marries him and makes him happy, altho like the
rest of the world, she may not always understand him.
54.
+ –
+
+
+
+
+
+ +
“Allthe earlier part of the book is shadowy, and hardly prepares
us for the vivid, admirable picture of life in New York that comes
later.”
Acad. 71: 527. N. 24, ’06. 190w.
“It is a book of fine fibre in purpose and execution, romantic,
touching, amusing.”
Nation. 83: 333. O. 18, ’06. 460w.
“‘Don-a-dreams’ is his first novel, but Mr. O’Higgins has made
no mistake in his new departure.” Otis Notman.
N. Y. Times. 11: 623. O. 6, ’06. 550w.
“It is all very tenderly and charmingly told, and we like it better
because our dreamer is not of those who think wallowing in the
mire synonymous with ‘knowing life.’”
N. Y. Times. 11: 705. O. 27, ’06. 400w.
N. Y. Times. 11: 796. D. 1, ’06. 200w.
“Its consistent literary quality lifts it far above the level of
ordinary fiction.”
Outlook. 84: 840. D. 1, ’06. 100w.
O’Higgins, Harvey Jerrold. Smoke-eaters. $1.50. Century.
Reviewed by Mary Moss.
Atlan. 97: 47. Ja. ’06. 140w.
Okakura-Kakuzo. Book of tea. **$1.50. Duffield.
These essays relate to tea, not as a beverage but as an aesthetic
symbol. “Within the pages of this volume is condensed the whole
philosophy of tea, together with its history, poetry, symbolism and
a synopsis of its relation to religion and art as they exist in Japan.
55.
+
+
–
+
The author writeswith sympathy ... and with a graceful felicity of
expression.” (Ind.)
Ath. 1906, 2: 512. O. 27. 160w.
“Charming group of essays.” Frederick W. Gookin.
Dial. 41: 105. S. 1, ’06. 1260w.
“What ‘Sartor resartus’ is to the realm of the utilitarian ‘The book
of tea’ is to the realm of the esthetic.”
Ind. 61: 461. Ag. 23, ’06. 280w.
R. of Rs. 34: 128. Jl. ’06. 50w.
Okey, Thomas. Story of Paris. $2. Macmillan.
“The greater part of the 400–odd pages of this volume are taken
up with the story of the city from its beginnings as a Gallo-Roman
camp to its expansive latter days. The last pages contain generous
descriptions of the landmarks, museums, galleries, churches, and
theatres of the present.” (N. Y. Times.) “It is not too much praise to
say that the book supplements the information contained in
Baedeker, and supplies as well a background for the greater
enjoyment of such volumes as Theodore Child’s ‘The praise of
Paris,’ Richard Whiteing’s ‘Paris of to-day,’ of Amicis’s ‘Ricordi di
Parigi.’” (Outlook.)
Ind. 61: 753. S. 27, ’06. 170w.
“The guide is a curious cross between a Baedeker and a Hare,
without the satisfying definiteness of the former or the charm of
the latter.”
Nation. 83: 167. Ag. 23, ’06. 240w.
N. Y. Times. 11: 434. Jl. 7, ’06. 130w.
56.
+ +
+
+
+
“The intendingvisitor to Paris could hardly have a more valuable
vade mecum than Mr. Okey’s little volume.”
Outlook. 83: 673. Jl. 21, ’06. 100w.
“We are glad to be able to commend highly this little book which
fully maintains the high standard which the volumes in this series
nearly always attain.”
Sat. R. 102: 277. S. 1, ’06. 190w.
“The historical, literary, and artistic aspects of the city are
worthily treated.”
Spec. 97: 65. Jl. 14, ’06. 60w.
Oliver, Frederick Scott. Alexander Hamilton: an essay on
American union. *$3.75. Putnam.
The work of an Englishman which gives an estimate of
Alexander Hamilton’s character and presents a record of political
and historical conditions in the United States in Hamilton’s day.
“Mr. Oliver calls his work an essay on American union; but it is far
more than that. At bottom it is a grave and singularly eloquent plea
for the great union of a close and lofty and disinterested
Imperialism. And it is an immense compliment to Mr. Oliver to say
that his conclusions and his exhortations are worthy of having been
directly inspired by such a figure as Alexander Hamilton.” (Lond.
Times.)
“A very thoughtful and clever essay on the life and work of
Alexander Hamilton.”
Ath. 1906, 2: 39. Jl. 14. 470w.
“Tho the book has some marked blemishes, it is so filled with
deep and original thinking that it is worthy the careful attention of
every student of Hamilton and our early political history. It is
written in an interesting, cultured style, which at times becomes
brilliant.”
57.
+ –
+
+ +
++ –
– –
+ –
Ind. 61: 1117. N. 8, ’06. 470w.
Ind. 61: 1170. N. 15, ’06. 20w.
“He has depicted Hamilton with force and clearness, with
humour, with sympathy and charm. He has treated a big subject in
a large and masterly way. No book has appeared lately which
conveys a more valuable lesson or one more tactfully and skilfully
unfolded.”
Lond. Times. 5: 165. My. 11, ’06. 3340w.
“To our minds, his narrative is by far the most interesting and
vivid account that has yet been published; but, being neither a
publicist nor an economist ... he is positively disqualified from the
task of estimating Hamilton’s work.”
Nation. 83: 204. S. 6, ’06. 1770w.
“There are some errors of fact, due perhaps to faulty proof
reading, but the worst fault is the author’s bias and distortion of
facts, which frequently make his conclusions valueless.” R. L.
Schuyler.
N. Y. Times. 11: 357. Je. 2, ’06. 1120w.
“As a portrait of Hamilton the work exhibits most of the defects
inherent in all admittedly partisan productions, and it further
suffers from the animus apparent in the treatment of those within
as well as without the Federalist party who placed themselves in
opposition to ‘the little lion.’ But his is a singularly fresh and in
many respects a singularly charming study, distinctive alike in
point of view, in method, and in style.”
Outlook. 83: 204. N. 3, ’06. 450w.
“Mr. Oliver’s book seems to us the most brilliant piece of
political biography which has appeared in England for many years.
A clear and vigorous style, wit, urbanity, a high sense of the
picturesque, and a remarkable power of character-drawing raise
much of it to the rank of a literary masterpiece.”
58.
+ + –
+–
+
+
–
Spec. 97: 58. Jl. 14, ’06. 2040w.
Olmsted, Stanley, Nonchalante. †$1.25. Holt.
Student life, especially the life of two American students in a
German university town, is cleverly handled in this story, and the
nonchalant heroine, with musical aspirations, is well suited to her
surroundings. The book presents a phase, a passing episode,
interesting and amusing, but superficial in that it deals with that
frivolous side of things which is so typical of student days. The
cafés, the theatres, the bleak boarding houses are well drawn, and
poor Fraulein Mittelini’s tragic struggle for fame is really worthy of
sympathy.
“The author has succeeded ... in giving [the heroine] some
genuine fascination. The style is too obviously imitative of that of
Mr. James.”
Critic. 48: 476. My. ’06. 50w.
“The grip of the book is the grip of Miss Bilton—but it is
entertaining even when she is off the stage.”
N. Y. Times. 11: 287. My. 5, ’06. 540w.
N. Y. Times. 11: 388. Je. 16, ’06. 160w.
R. of Rs. 33: 758. Je. ’06. 100w.
Oman, Charles William Chadwick. Inaugural lecture on the
study of history delivered on Wednesday, Feb. 7, 1906. *35c.
Oxford.
In this lecture on the teaching and study of history the Chichele
professor “perceives the great virtues of the tutorial system. He
recognizes a fact which is often overlooked by zealous reformers,
that no system of teaching can flourish which does not meet the
wants of the learners; and this general truth is in a particular sense
applicable to the universities of England.... The fact ‘that must be
59.
+ +
+
faced is,that Oxford is a place of education as well as a place of
research,’—these words strike the real keynote of Professor Oman’s
inaugural address.” (Nation.)
“It is remarkable for several characteristics and for a good deal of
courage. From start to finish it is lively; the writing, while it is
occasionally of great dignity is sometimes brilliant and even
humorous.”
Ath. 1906, 1: 322. Mr. 17. 1100w.
Nation. 82: 388. My. 10, ’06. 1100w.
Omar Khayyam. Rubaiyat: a new metrical version; rendered into
English from various Persian sources, by George Roe, with
introd. and notes. **$1.50. McClurg.
The translation adopts a middle course between the versions of
Omar which sacrifice the letter to the requirements of good verse
and those which in order to be literal, sacrifice the spirit to the
letter.
Dial. 41: 400. D. 1, ’06. 70w.
Omond, George William Thomson. Bruges and West Flanders;
painted by Amedee Forestier; described by G. W. T. Omond. *$3.
Macmillan.
In the main Mr. Omond treats his subject historically, but even
from this point of view, he catches the spirit of sentiment and
romance. “Each one of these quaint, often-despoiled towns has
remaining some romantic relics and picturesque buildings—belfry,
market-place, Hotel de Ville—old gateways, or churches enriched
with paintings.” (Outlook.) “And what Mr. Omond so successfully
does for Bruge-LaMorte, he also does for the other towns of West
Flanders—Ypres, Furnes, Nieuport—revivifying them with the
story of a glorious past.” (N. Y. Times.)
60.
+
+ –
+
+
+
+ –
+
Ind.61: 754. S. 27, ’06. 160w.
Nation. 82: 279. Ap. 5, ’06. 80w.
“He has been deeply touched by the ruined greatness that
surrounds prosperous Ostend and would show others how they
may come under the spell.”
N. Y. Times. 11: 145. Mr. 10, ’06. 870w.
“While the text of the book is not remarkable in any way, it is
written in clear, simple style.”
Outlook. 82: 715. Mr. 24, ’06. 340w.
R. of Rs. 33: 508. Ap. ’06. 120w.
Sat. R. 101: 664. My. 26, ’06. 320w.
Oppenheim, E. Phillips. Maker of history. †$1.50. Little.
The plot of Mr. Oppenheim’s new story with a mystery grows out
of an episode in which an English youth actually witnesses a
meeting between the Czar of Russia and the Emperor of Germany,
and turns up in Paris with a loose sheet of a treaty between the two,
relative to an attack upon England. How this same Englishman is
hidden away in Paris by spies, and why his sister is also abducted,
and what sympathies stir one Sir George Duncombe to action in
their behalf furnish motive power for a lively story.
“Is a capital story filled with mysterious and exciting
happenings, but one regrets to see Mr. Oppenheim writing down to
this level after he has shown that he is capable of such work as ‘A
prince of sinners.’” Amy C. Rich.
Arena. 35: 447. Ap. ’06. 330w.
“It is an amazing medley, highly characteristic of the author.
Without trenching on politics, one may be permitted to doubt the
wisdom just now of accentuating the jealousies of nations.”
61.
+ –
+
+
+
+ +
+–
+
Ath. 1905, 2: 432. S. 30. 190w.
“In substance, of course, it is merely a sort of exalted dime novel.
But is written with such admirable restraint, such a matter-of-fact
style, as though the events were being chronicled for the columns
of a conservative daily newspaper, that you are cleverly led on from
mild curiosity to a breathless sort of interest.” Frederic Taber
Cooper.
Bookm. 22: 633. F. ’06. 560w.
“This stirring story is told with neatness and dispatch.” Wm. M.
Payne.
Dial. 40: 154. Mr. 1, ’06. 250w.
“Mr. Oppenheim handles his material cleverly and makes of it a
good story of adventure.”
Ind. 60: 1166. My. 17, ’06. 300w.
“The story is told with the vim and dash characteristic of Mr.
Oppenheim’s work, and is one of the best tales he has yet
produced.”
Lit. D. 32: 332. Mr. 3, ’06. 140w.
N. Y. Times. 10: 925. D. 30, ’05. 90w.
“The story proceeds with cumulative interest to the end. The love
interest of the story is secondary, but good, although the character
drawing is occasionally exaggerated.” Stephen Chalmers.
N. Y. Times. 11: 15. Ja. 13, ’06. 820w.
N. Y. Times. 11: 385. Je. 16, ’06. 110w.
“Altogether the romance is an exceptionally good specimen of
sensational story-telling.”
Outlook. 82: 231. Ja. 27, ’06. 150w.
“It is all nonsense, but it is not boring nonsense.”
62.
+
+ –
+ –
+
Sat.R. 100: sup. 8. O. 14, ’05. 420w.
Spec. 95: 571. O. 14, ’05. 130w.
Oppenheim, Edward Phillips. Man and his kingdom. † $1.50.
Little.
Love, intrigue and revolution in a South American state make a
riotous setting for Mr. Oppenheim’s story. The man of the hour is a
wealthy young Englishman who sides neither with the
revolutionists nor yet with the president’s party, but is a friend to
both. His Beau Desir, a fertile valley near the town, with two
hundred Englishmen to till it would fain express the temper of his
neutrality, but the disquieting elements of the town creep into it.
There are lively quarrels, attempted murders, and thrilling escapes,
all of which have local color and atmosphere.
Oppenheim, E. Phillips. Master mummer. †$1.50. Little.
“Mr. Oppenheim has trodden a beaten path when, it would seem
from his earlier success in invention, he might have struck out
afresh for himself.”
Reader. 7: 562. Ap. ’06. 180w.
Oppenheim, Edward Phillips. Millionaire of yesterday. †$1.50.
Little.
A new illustrated edition of Mr. Oppenheim’s story that gives a
vivid picture of two men, widely divergent types, one an invincible
hero, the other a leaner, in the African bush making a grim fight for
life and fortune.
Oppenheim, Lassa. International law. *$6.50. Longmans.
Edinburgh R. 203: 471. Ap. ’06. 8240w.
“The best and most important part of this system is his rule of
giving his readers the law as it is, and not as it ought to be. This,
63.
+ +
+ +
+
+
combinedwith his natural impartiality, makes his book an
extremely fair and rational one.”
Nation. 82: 373. My. 3, ’06. 460w.
“The arrangement is clear and logical, and the matter of the work
is, so far as we have examined it, fully up to date, and presented
with acumen and moderation.”
Spec. 96: 544. Ap. 7, ’06. 280w.
Orczy, Emma Magdalena Rosalia Maria Josefa Barbara,
baroness. I will repay. †$1.50. Lippincott.
The scenes of Baroness Orczy’s dramatic tale are enacted in the
terrible days of the French revolution. Ten years before its reign of
terror, Juliette Marny is compelled by her father to take a vow to
bring about the ruin and death of Paul Déroulède, the man who,
tho against his will, had killed her brother in a duel. So much for
the prologue. When the story opens, the revolution is well under
way. Déroulède is a popular leader. Juliette, housed with his
mother for safety, loves him, yet is obedient to relentless Fate
which is dragging her to the fulfillment of her vow. She denounces
him to the terrorists, and in attempting to undo her treachery
brings both herself and Déroulède under the Merlin suspect law.
Their escape from France closes a chapter of thrilling incidents.
“There are not so many characters to stage in this book as in a
former success of the same author’s, dealing, like this with
revolutionary Paris, and we find less variety of scene, less incident:
but the same dramatic power is abundantly demonstrated.”
Ath. 1906. 2: 579. N. 10. 150w.
“It is, in truth, a very fair story of its semihistoric wholly
respectable sort.”
Nation. 83: 539. D. 20, ’06. 390w.
“The story is full of exciting situations and thrilling moments.”
64.
+
+ –
– +
+
+–
+
+ –
N. Y. Times. 11: 798. D. 1, ’06. 100w.
N. Y. Times. 11: 869. D. 15, ’06. 630w.
Sat. R. 102: 648. N. 24, ’06. 90w.
Orczy, Baroness. Scarlet pimpernel. †$1.50. Putnam.
“A brilliantly vivid story abounding in dramatic incident.”
Critic. 48: 476. My. ’06. 80w.
Orczy, Emma Magdalena Rosalia Maria Josefa Barbara,
baroness. Son of the people: a romance of the Hungarian
plains. †$1.50. Putnam.
The old story of the rich and handsome peasant who wins the
hand of an impoverished nobleman’s daughter against her will and
later, by proving his nobility of soul, turns her scorn to love, is
given a charming Hungarian setting in this romance of the plains.
The peasant life and character are strongly contrasted with the
traditional pride of the nobility; the lines of caste are well
portrayed, the priest, the Jew, the aristocrat, and the son of the
soil, the thrift of the peasant, the prodigality of the lord are all
interwoven with the love story.
“It is sentimental and of a conventional type, but the setting is
new, and so it takes on a novel air.”
Ath. 1906, 1: 227. F. 24. 260w.
“It is a strong and attractive piece of work, vivid in description
and characterization, dramatic in action.” Wm. M. Payne.
Dial. 41: 241. O. 16, ’06. 230w.
“The story is well told, and as interesting as any other thrice told
tale.”
Ind. 61: 825. O. 4, ’06. 110w.
65.
+ –
+ –
+
+–
+ –
“This really interesting book is hurt by wordiness and repetitions
of good effects, yet not unto destruction.”
Nation. 83: 59. Jl. 19, ’06. 430w.
N. Y. Times. 11: 383. Je. 16, ’06. 110w.
“Judicious condensation and elimination would have greatly
improved and strengthened ‘A son of the people,’ but it has decided
merits as it is.”
N. Y. Times. 11: 506. Ag. 18, ’06. 490w.
Putnam’s. 1: 224. N. ’06. 160w.
“The book interests and attracts despite the poverty of the plot.”
Sat. R. 101: 244. F. 24, ’06. 200w.
Orr, Rev. James. God’s image in man and its defacement in the
light of modern ideals. **$1.75. Armstrong.
Professor Orr discusses “the conflict between the Biblical and the
modern view of man—his nature, origin, and primitive condition,
his sinfulness and the divine redemption from it. The difference
between the so-called Biblical and the modern views is that the
former regards God’s image in man as aboriginal, the latter regards
it as ultimate. Man’s redemption from sin, therefore, the former
regards as a reconstructive work, the latter as constructive or
evolutionary.”—Outlook.
“There can be no question of Professor Orr’s deep religious
interest, his courage, his marvelous grasp of the material of
present-day learning, and his perception of the seriousness of the
questions now pressing for solution; but I do not think that the
work under review can give much help to a man who is seized of
the significance of the great intellectual and religious movements
of the present and feels a sympathetic interest in them.” George
Cross.
Bib. World. 28: 220. S. ’06. 1290w.
66.
+ –
+
+ +–
+ +
+ +
Ind. 61: 823. O. 4, ’06. 510w.
“What seems hardly fair in Professor Orr’s argument is the
prominence given to Haeckel as the representative of the modern
view.”
Outlook. 81: 940. D. 16, ’05. 220w.
“Dr. Orr conducts his argument with a creditable moderation of
language, and states the problems which he discusses fairly.”
Spec. 95: 986. D. 9, ’05. 250w.
Orr, Rev. James. Problem of the Old Testament considered with
reference to recent criticism. **$1.50. Scribner.
“A volume of lectures given at Lake Forest college by Dr. James
Orr, of Glasgow. Dr. Orr represents the conservative view in his
attitude toward modern criticism. The present volume is largely
devoted to the repetition of the Graf-Wellhausen hypothesis.”—R.
of Rs.
“The temper of the book is admirable. Dr. Orr’s disposition of his
material appears to be excellent. We think it is safe to say that
nowhere will the student find in so compact a form an abler
arraignment of the Graf-Wellhausen hypothesis, which is Dr. Orr’s
immediate object of attack, than in the present work.” Kemper
Fullerton.
Am. J. Theol. 10: 705. O. ’06. 1720w.
“A comprehensive survey of the chief problems of the Old
Testament from the conservative point of view, but considered
with fairness and candor.”
Bib. World. 27: 399. My. ’06. 20w.
Bibliotheca Sacra. 63: 374. Ap. ’06. 440w.
“There is no book in English that presents with such fulness and
strength, from the conservative point of view, the problem of the
67.
+ +
+
+
+ –
–+
+ –
Old Testament.”
Dial. 41: 41. Jl. 16, ’06. 350w.
“Professor Orr is astute, a keen logician, and he has made
himself a thoro master of his material.”
Ind. 60: 1490. Je. 21, ’06. 940w.
Ind. 61: 1166. N. 15. ’06. 120w.
“The problem of old Testament is twofold—religious and literary.
So far as the principles of the religious aspect of the problem are
concerned, we agree with him; but so far as the literary aspect of
the problem is concerned, we take leave to doubt.”
Lond. Times. 5: 130. Ap. 12, ’06. 1390w.
“The multitudinous points taken by Dr. Orr against the
prevailing critical opinions present to the unlearned reader a
formidable array.”
Outlook. 82: 570. Mr. 10, ’06. 500w.
R. of Rs. 33: 510. Ap. ’06. 50w.
“We may say that Dr. Orr is a strong conservative, though fifty
years ago he would have been regarded as a dangerous radical, that
he has stated his subject thoroughly, though not we cannot but
think, with an open mind; and that he always expresses himself
with courtesy and good taste.”
Spec. 96: 305. F. 24, ’06. 230w.
Osborn, Albert. John Fletcher Hurst: a biography. *$2. Meth. bk.
A biography which is autobiographic in nature so successfully
has the compiler eliminated himself in producing what the Bishop
said or what has been said about him. The sketch touches upon his
boyhood, education, European experiences, ministerial work, and
duties as president of the Drew theological seminary.
68.
+
+
+ +
+
+
+
+ –
+
Reviewedby Erl B. Hulbert.
Am. J. Theol. 10: 362. Ap. ’06. 90w.
“The task has been performed with equal loyalty and ability, and
the book is every way a fitting memorial of a man of great gifts,
high character, and broad influence.”
Critic. 48: 285. Mr. ’06. 60w.
“Mr. Osborn’s biography, in a word, is a worthy memorial of a
great Christian and a great American, and a book which should
enlarge the horizon and stimulate to a higher life all into whose
hands it falls.”
Lit. D. 32: 492. Mr. 31, ’06. 430w.
“The only criticism to be brought against this biography is that
the index is extravagant in its dimensions.”
N. Y. Times. 11: 14. Ja. 13, ’06. 520w.
Outlook. 81: 1039. D. 23, ’05. 160w.
R. of Rs. 33: 253. F. ’06. 90w.
Osbourne, Lloyd. Motormaniacs. [+]75c. Bobbs.
“Pretty little stories they are too, when we are permitted to pause
and enjoy them, and the motormaniacs are always entertaining
and capital company to the end of the run.”
Acad. 71: 399. O. 20, ’06. 110w.
“The dialogue is comic, and the narrative runs with a swing and
zest which are valuable aids to easy reading.”
Ath. 1906, 2: 545. N. 3. 100w.
“The book is full of humour and energy.”
Spec. 97: 497. O. 6, ’06. 110w.
69.
+
+
+ –
+
+ –
+
Osbourne,Lloyd. Wild justice. †$1.50. Appleton.
Nine stories of life in the South sea islands which take their title
from the first tale. The author spent a number of years among
crude Pacific natives with his step-father Robert Louis Stevenson.
His characters are drawn from these inhabitants “well-meaning
but generally inefficient missionaries, unscrupulous traders, and
refugees and adventurers in search of victims. It is not an edifying
life, and the manly virtues seem to be conspicuously absent.”
(Outlook.)
“They are all good, but of no one of them can it be said that it is
strikingly and exceptionally good.”
Ath. 1906, 1: 510. Ap. 28. 240w.
“The tales all have a swing in the telling and show that the author
is in his own field.”
Critic. 48: 476. My. ’06. 70w.
Lond. Times. 5: 149. Ap. 27, ’06. 550w.
“The fascination of the unusual pervades its pages.”
N. Y. Times. 11: 197. Mr. 31, ’06. 320w.
“There is a certain bizarre humor, however, in these tales which
somewhat redeems the sordidness of their subject matter.”
Outlook. 82: 859. Ap. 14, ’06. 70w.
Sat. R. 101: 625. My. 19, ’06. 60w.
Osgood, Herbert Levi. American colonies in the 17th century. 2v.
**$5. Macmillan.
“As a whole the work is the first adequate account of the origin,
character, and development of the American colonies as
institutions of government and as parts of a great colonial system;
and it displays, on the part of the author wide and deep knowledge
of the documentary evidence for colonial history and rare powers
70.
+ + –
+–
– –
+
of analysis and interpretation. In a style remarkably clear, forcible
and accurate the reader will regret the presence of so many cleft
infinitives.” Charles M. Andrews.
Am. Hist. R. 11: 397. Ja. ’06. 3060w. (Review of v. 1 and 2.)
O’Shea, Michael Vincent. Dynamic factors in education. *$1.25.
Macmillan.
“It has been the author’s object to show that in the early years of
a child’s school life, ‘motor expression’ in his teaching is ‘essential
to all learning.’ He has endeavored to indicate mainly in outline,
‘how the requirements of dynamic education can be provided for in
all departments of school work.’ Further, he says in his preface, ‘I
have sought to point out that there is a definite order in which the
motor powers develop, and that in our instruction we will achieve
the highest success only as we conform quite closely to this
order.’”—N. Y. Times.
“It is clear and, if one is not annoyed by its diffuseness,
interesting.”
Bookm. 23: 654. Ag. ’06. 180w.
“The book seems poorly suited for the use of the practical
teacher for whom it is announced, or the professional student. In
spite of the author’s resolution to the contrary, it is burdened with
methods of investigation, where results alone should be given.”
Edward O. Sisson.
Dial. 41: 89. Ag. 16, ’06. 580w.
“A fair and comprehensive book. It is sound psychology sensibly
applied.”
Ind. 61: 263. Ag. 2, ’06. 50w.
N. Y. Times. 11: 201. Mr. 31, ’06. 310w.
“The whole volume is what the subject is, dynamic, and is as
important for parents as for teachers.”
71.
+ +
+ +
++
+ +
+
Outlook. 83: 90. My. 12, ’06. 200w.
“It is admirably suited to be a handbook for advanced classes
who desire to pursue special topics exhaustively, by first reading a
guidebook and then following up the literature of the subject. The
style is so clear and the treatment so concrete and inductive that
the general reader will understand most of it. One of Professor
O’Shea’s chief contributions is in selecting those laws and
phenomena that have an educational application and clearly
showing the application.” Frederick E. Bolton.
Psychol. Bull. 3: 367. N. 15, ’06. 510w.
“Is not epoch marking; it is in part what has been said before by
other writers, but it has two virtues—it is reasonably complete, and
it is of great importance.”
Pub. Opin. 40: 639. My. 19, ’06. 90w.
“On the whole, we know of no more satisfactory discussion of
what is thus far known of the evolution of motor control, its
relation to education, and of the place of the manual arts in
education.”
School R. 14: 459. Je. ’06. 510w.
“The style is not so overburdened with ‘educational jargon’ as to
interfere with the enjoyment and edification of the general reader.”
World To-Day. 11: 764. Jl. ’06. 240w.
Osler, William. Counsels and ideals; from the writings of William
Osler. **$1.25. Houghton.
In culling selections from his less technical lectures and
addresses, Dr. Osler aims to offer “individual influence” and
“inspiration” to the student or general reader. “Wise counsels
abound in this volume—counsels inspired by high ideals and wide
experience. The real man whom they present is no more like the
individual whose words were so travestied by the press on a recent
occasion as to threaten the dictionary-makers with a new word,
72.
+ +
+
+
+ +
+
+
+
+
‘oslerize,’than the caricature of the political cartoonist is like its
original.” (Outlook.)
“A book which may be read with pleasure and lasting profit, not
only by every member of the medical profession, but also by the
general public. Dr. Camac has made his selection with judgment.”
Ath. 1906, 1: 301. Mr. 10. 730w.
Critic. 49: 95. Jl. ’06. 190w.
“What most impresses one on examining this selection from
forty-seven of the author’s fugitive pieces is not only the
professional and practical wisdom displayed, and the breadth of
view revealed, but also the wide reading in writers not commonly
held to be a necessary part of a doctor’s library.”
Dial. 40: 93. F. 1, ’06. 390w.
Ind. 60: 929. Ap. 19, ’06. 340w.
“They afford very interesting reading.”
N. Y. Times. 11: 18. Ja. 13, ’06. 680w.
“What he writes, however, is of household, individual interest,
and it is presented in a manner which causes facts to breathe
eloquence and conviction.”
N. Y. Times. 11: 48. Ja. 27, ’06. 350w.
“To dip into these pages anywhere is to meet with a thoughtful,
strong, and sagacious man.”
Outlook. 82: 140. Ja. 20, ’06. 130w.
R. of Rs. 33: 256. F. ’06. 90w.
Ostwald, Wilhelm. Conversations on chemistry. Pt. 1, General
chemistry; authorized tr. by Elizabeth Catherine Ramsay, $1.50;
Pt. 2, Chemistry of the most important elements and
compounds; authorized tr. by Stuart K. Turnbull, $2. Wiley.
73.
+ +
+ +–
+
The authorized translation of Ostwald’s “Die schule der chemie.”
Addressed distinctly to elementary pupils, the subject is presented
in dialogue, the conversations taking place between master and
pupil. Such subjects are treated as substance, properties, solution,
melting and freezing, density, compounds, elements, oxygen,
hydrogen, nitrogen, air, etc.
“Miss Ramsay has done her work with much skill, and has made
the dialogue not less natural and vivacious than it is in the
original.”
Nature. 72: 364. Ag. 17, ’05. 330w. (Review of pt. 1.)
“Most points are worked out with great ingenuity and address to
an entirely logical conclusion. The allusion to things and
phenomena of real human interest and the suppression of
pedantry are also to be warmly commended. The actual work of
translation has, on the whole, been well done.” A. S.
Nature. 74: 173. Je. 21, ’06. 570w. (Review of pt. 2.)
“The chief value of the book, must lie, therefore, in showing
something of the spirit and the methods best adapted for arousing
the interest of the young pupils in elementary science.” William
McPherson.
Science, n.s. 22: 829. D. 22, 05. 220w. (Review of pt. 1.)
Ostwald, Wilhelm. Individuality and immortality: the Ingersoll
lectures, 1906. **75c. Houghton.
Professor Ostwald, professor of chemistry at the university of
Leipzig, treats the question scientifically. “At the very outset, the
lecturer calls attention to the fact that our knowledge ‘is an
incomplete piece of patchwork;’” but, he adds, “each one is bound
to make the best possible use of it, such as it is, never forgetting
that it may at any time be superseded by new discoveries or ideas.
In this truly scientific spirit, very remote from the dogmatism of
the churches, Professor Ostwald proceeds to consider what
74.
+
+ –
+
–
+
+ +
immortalitymay be supposed to be, and what reasons we have for
believing it.” (Dial.)
“The chief value of this work is in showing the attitude which the
scientifically trained mind tends to take to those problems where
the clear principles and positive methods of the physical sciences
do not obtain.” W. C. Keirstead.
Am. J. Theol. 10: 555. Jl. ’06. 560w.
“The discussion is an interesting one, both from its statement of
scientific views and from the glimpse it affords of the mind of the
author. It is, nevertheless, strangely incomplete, almost ignoring
the deeper questions at issue.” T. D. A. Cockerell.
Dial. 40: 228. Ap. 1, ’06. 1680w.
“It is an exceedingly interesting discourse, and quite up to date,
scientifically speaking; it is full of fine moral thoughts, but it
contains very little Christian consolation.”
N. Y. Times. 11: 116. F. 24, ’06. 230w.
Outlook. 82: 716. Mr. 24, ’06. 150w.
Ottley, Rev. Robert Lawrence. Religion of Israel: a historical
sketch. *$1. Macmillan.
“It is a readable outline of the history from a modern point of
view, chiefly at second-hand.” George F. Moore.
Am. J. Theol. 10: 144. Ja. ’06. 70w.
Outram, James. In the heart of the Canadian Rockies. **$3.
Macmillan.
“His counsel is sound, and his knowledge reaches far. The
volume was well worth writing.”
Ath. 1906, 1: 13. Ja. 6. 560w.
75.
+
+ +
+ +–
+
Critic. 48: 94. Ja. ’06. 20w.
Ind. 60: 457. F. 22, ’06. 420w.
“He has succeeded in producing a useful piece of work, which
brings together an account of all that has been accomplished in the
Canadian Rockies by himself and by other kindred spirits.” G. W.
L.
Nature. 73: 362. F. 15, ’06. 720w.
“The book is written by a man who has his soul in the story.”
Pub. Opin. 40: 26. Ja. 6, ’06. 210w.
77.
+ + –
+
+
++
P
“P., Q.” How-to buy life insurance. **$1.20. Doubleday.
A book that “has been written and published in the interest of
the policyholder primarily. It undertakes to free the subject from
the technical obscurities that so frequently interfere with a clear
understanding of its elements and to give the plain citizen
straightforward advice and information as to the various types of
policies in the market and the relative advantages of each.” (R. of
Rs.)
“As a practical guide to the policyholder desirous of figuring out
for himself the real cost of his insurance and of choosing between
rival companies, ought to be found of substantial value by the busy
man, because of the comparative tables and specimen blanks given
in the appendix. These could be considerably improved upon in
certain respects, but they are a distinct advance over what has been
furnished by most other books on the subject.”
Dial. 41: 117. S. 1, ’06. 350w.
N. Y. Times. 11: 408. Je. 23, ’06. 720w.
“It is a helpful and suggestive manual.”
R. of Rs. 34: 126. Jl. ’06. 80w.
Page, Curtis Hidden, ed. Chief American poets: selected poems by
Bryant, Poe, Emerson, Longfellow, Whittier, Holmes, Lowell,
Whitman, and Lanier. *$1.75. Houghton.
“The selections have been made with good taste and judgment
and the notes are ample and to the point.”
Critic. 48: 91. Ja. ’06. 90w.
78.
+ +
+ +–
+
+
+
+
+
+
+ +
Dial. 40: 96. F. 1, ’06. 150w.
Nation. 82: 158. F. 22, ’06. 170w.
“Such a book would be a great convenience for the use of a class
studying American literature.”
School R. 14: 233. Mr. ’06. 100w.
Page, Thomas Nelson. Negro: the southerner’s problem. **$1.25.
Scribner.
“These essays are characterized by a sanity of spirit and a
painstaking thoroughness.” C: A. Elwood.
Am. J. Soc. 11: 698. Mr. ’06. 440w.
Outlook. 83: 88. My. 12, ’06. 510w.
Page, Thomas Nelson. On Newfound river. †$1.50. Scribner.
“In the story we meet ... the Southern life of an earlier day: hot-
tempered men and gracious women, trusty slaves, negro-hunting
whites, the grocery-store-town-meeting, and the open-air court of
justice. The love-story, however, is the thing and is young,
Arcadian, rough-running, happily arriving. Mr. Page explains that
it is a story enlarged; explicitly not a novel, but ‘a love story, pure
and simple,’ and such it will be found.”—Nation.
Dial. 41: 286. N. 1, ’06. 40w.
Lit. D. 33: 596. O. 27, ’06. 130w.
Lit. D. 33: 858. D. 8, ’06. 80w.
“A delicate, finished specimen of its author’s art.”
Nation. 83: 332. O. 18, ’06. 170w.
“It is a story pure and sweet amid the poisonous blossoms of
fiction that nowadays spring thick, an idyll of loyalty and of love,
79.
+ +
+
+ +
+
thrilledthrough and through with ‘the tender grace of a day that is
dead.’”
N. Y. Times. 11: 744. N. 10, ’06. 440w.
“The most appreciative comment that can be made on this story
is that he has not spoiled it; the old charm still lingers.”
Outlook. 84: 709. N. 24, ’06. 80w.
Paine, Albert Bigelow. Little garden calendar for boys and girls.
$1. Altemus.
“This is one of the best children’s books in recent years. It is
bright and entertaining and while holding the interest of the young
in the story that is told, it imparts a vast fund of information which
every child should know.”
Arena. 35: 222. F. ’06. 320w.
Paine, Albert Bigelow. Lucky piece: a story of the North woods.
$1.50. Outing pub.
A tale of the Adirondacks whose hero is an idle young man of
more wealth than ambition, and whose heroine undertakes to
teach him the definite purpose in life. A Spanish luck piece brings
friends, wealth and happiness in its train of talismanic bestowals.
“This is a pleasant story, with some well-drawn characters and
just enough plot to carry the reader comfortably along to the last
chapter.”
Critic. 49: 191. Ag. ’06. 50w.
N. Y. Times. 11: 274. Ap. 28, ’06. 370w.
Paine, Albert Bigelow. Sailor of fortune; memoirs of Capt. B. S.
Osbon. **$1.20. McClure.
80.
+
+
+
+
+
“Captain Osbon, whosememoirs are given practically as he
detailed them to the writer, Mr. Albert Bigelow Paine, lived among
some of the most stirring scenes of the past century, and his
narrative presents with extraordinary vividness events of which he
was an actor or an eye-witness.” (Lit. D.) “This lively record covers
whaling, buccaneering, the Civil war, journalism, and almost
everything but love.” (World To-Day.)
“Mr. Paine, the redactor of these stories of sea life, has succeeded
admirably in preserving the personal quality of the actor-narrator,
and we easily accept the ‘yarns’ as a long succession of fireside
talks face to face with the man who lived them.”
Lit. D. 33: 556. O. 20, ’06. 180w.
“Cannot fail to be a joy to old and young.”
N. Y. Times. 11: 667. O. 13, ’06. 190w.
“His reminiscences of famous men are numerous and
characteristic.”
N. Y. Times. 11: 800. D. 1, ’06. 250w.
“Mr. Paine has done well what must have been a difficult task.
The book will amuse and enchain the reader who has a love for the
unusual and picturesque.”
Putnam’s. 1: 381. D. ’06. 110w.
“Every chapter reads like a condensed historical novel.”
World To-Day. 11: 1222. N. ’06. 50w.
Paine, Dorothy C. Maid of the mountains. †$1. Jacobs.
To Carol, a mountain maid of North Carolina, comes a good fairy
in the guise of Beth, a happy tender-hearted little girl, who brings
real aid to the sufferings of the mountain family. Among other
things, she gives a benefit entertainment in which it is discovered
that Carol has a beautiful voice, and a wealthy but childless woman
81.
Welcome to ourwebsite – the ideal destination for book lovers and
knowledge seekers. With a mission to inspire endlessly, we offer a
vast collection of books, ranging from classic literary works to
specialized publications, self-development books, and children's
literature. Each book is a new journey of discovery, expanding
knowledge and enriching the soul of the reade
Our website is not just a platform for buying books, but a bridge
connecting readers to the timeless values of culture and wisdom. With
an elegant, user-friendly interface and an intelligent search system,
we are committed to providing a quick and convenient shopping
experience. Additionally, our special promotions and home delivery
services ensure that you save time and fully enjoy the joy of reading.
Let us accompany you on the journey of exploring knowledge and
personal growth!
ebookultra.com



















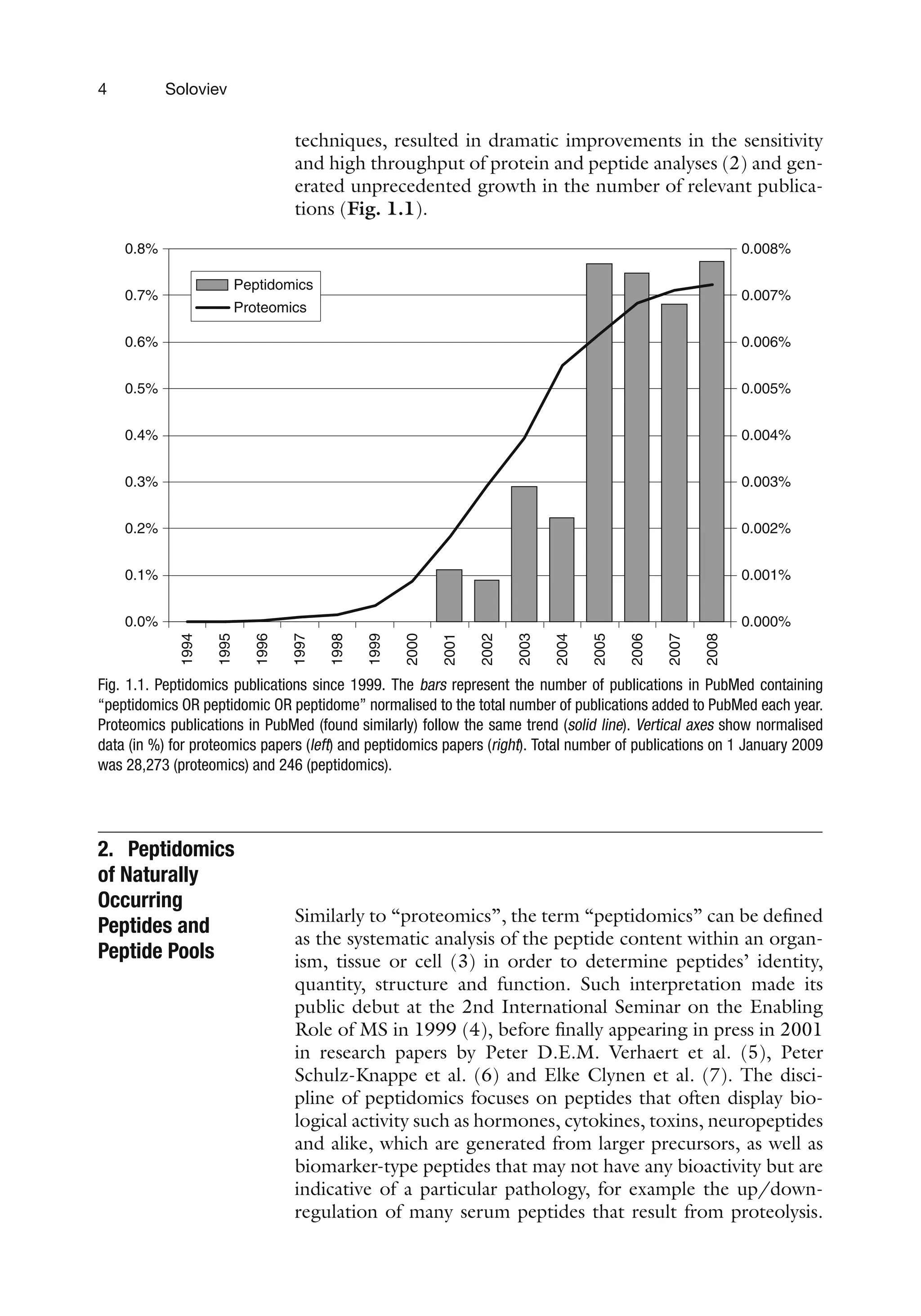








![Comparative Peptidomics of Salmonella 15
2. Ammonium bicarbonate, isopropanol, and methanol (Sigma
Aldrich).
3. Protease inhibitor cocktail formulated for use with bacterial
cell extracts (Cat. No. P8465, Sigma)
4. 0.1 mm zirconia/silica beads (BioSpec Products,
Bartlesville, OK) were used for cell lysis.
5. 10–20% Tris-Tricine Ready Gels R
(Bio-Rad, Hercules, CA)
and GelCode R
Blue reagent from Pierce for SDS-PAGE
analyses.
6. OMIX R
C-18 tips (100 L) (Varian, Inc, Palo Alto, CA) for
sample solid-phase extraction (SPE) clean-up prior to MS
analysis.
7. SpeedVac (Thermo Fisher Scientific, Waltham, MA) to con-
centrate samples.
2.3. Liquid
Chromatography–
Mass
Spectrometry/Mass
Spectrometry
1. Ion trap mass spectrometers (LTQ, Thermo Fisher Scien-
tific, San Jose, CA)
2. Water purified using a NANOpure R
or equivalent system
(≥ 18 M×cm)
3. Mobile phase A: Degassed 0.2% acetic acid, 0.05% trifluo-
roacetic acid in water (Sigma Aldrich)
4. Mobile phase B: Degassed 0.1% trifluoroacetic acid in 90%
acetonitrile (ACN), 10% water (Sigma Aldrich)
5. 5-m Jupiter C18 stationary phase (Phenomenex, Torrence,
CA) packed into 60-cm (360 m o.d. X 150 m i.d.)
fused silica capillary tubing (Polymicro Technologies Inc.,
Phoenix, AZ)
6. Liquid chromatography system is described elsewhere by
Livesay et al. (15)
2.4. Liquid
Chromatography–
High-Resolution
Mass Spectrometry
1. Fourier transform ion cyclotron resonance (FTICR) mass
spectrometer, either a custom-built 11 T instrument or 9.4
T instrument (Bruker Daltonics, Billerica, MA).
2. See Section 2.3 for details on mobile and stationary-phase
materials.
2.5. LC–MS/MS Data
Analyses
1. SEQUEST R
version [TurboSEQUEST R
(cluster) v.27 (rev.
12), Thermo Fisher Corp.]
.
2.6. Proteomics 1. RapiGestTM
(Waters, Milford, MA) is a surfactant to aid in
the solubilization and trypsin digestion of proteins.
2. Trypsin for protein digestion (Promega, Madison, WI)](https://image.slidesharecdn.com/34348-250226191938-6b836bd8/75/Peptidomics-Methods-and-Protocols-1st-Edition-Mikhail-Soloviev-Auth-30-2048.jpg)


![18 Adkins et al.
5. Prior to MS analysis, the concentrated isopropanol extracts
are cleaned via solid-phase extraction using OMIX C18
pipette tips. These tips are monolithic, rather than particu-
late, and are therefore much easier to use without clogging,
while providing better recovery and reproducibility. Thirty
micrograms of peptide mass from each sample is applied to
a 100-L tip. The directions provided by the manufacturer
are used to condition, wash, and load the samples. Peptides
are eluted from the tips with 80:20 ACN:H2O containing
0.1% TFA. Eluted peptides are concentrated to ∼15 L in a
SpeedVac.
6. Alternatively, samples can be fractionated using strong cation
exchange (SCX) HPLC to minimize sample complexity prior
to each LC–MS/MS analysis, as described previously (7).
Each fractionation is performed using approximately 150 g
(peptide mass) of concentrated isopropanol extract, result-
ing in 25 fractions that are concentrated in a SpeedVac
to dryness. The samples are then reconstituted in 25 mM
NH4HCO3 to a volume appropriate for LC–MS/MS
analysis.
3.3. Liquid
Chromatography–
Mass
Spectrometry/Mass
Spectrometry
Our analytical instrumentation consists of commercially avail-
able platforms [e.g., ion traps (LTQ from ThermoFisher) and
FTICR–MS (BrukerDaltonics)] that are in-house modified to
increase the sensitivity and throughput of the analyses. How-
ever, the below LC–MS(/MS) approaches can be applied at a
reasonable level of quality with more generally available off-the-
shelf instrumentation. LC–MS/MS analyses are useful for mak-
ing identifications and for semi-quantitation based on “spectrum
counting” techniques (4, 8–10). These analyses are also used
to build a database of identified peptides annotated with deter-
mined reversed-phase elution times (11) and calculated masses.
This database (also referred to as a mass and time tag lookup
table) is used with results from the high-resolution MS analyses
(Section 3.4) to increase throughput, perform label-free quanti-
tation, and improve peptide-sampling methods in the MS exper-
iment. This is a simplified description of the accurate mass and
time (AMT)-tag process developed in our laboratory, which has
been extensively discussed elsewhere (12–14).
1. The concentrated C18 SPE eluents from the peptide clean-
up procedure and the SCX fractions are then analyzed
by reversed-phase microcapillary HPLC (15) interfaced
through nanoelectrospray ionization (nanoESI) to an ion
trap mass spectrometer, as described previously (4). Briefly,
the technique used in our laboratory entails gradient elution
of peptides over 100 min using a 360 m OD × 150 m
ID × 65 cm long capillary column packed with 5 m Jupiter
C18 particles.](https://image.slidesharecdn.com/34348-250226191938-6b836bd8/75/Peptidomics-Methods-and-Protocols-1st-Edition-Mikhail-Soloviev-Auth-33-2048.jpg)



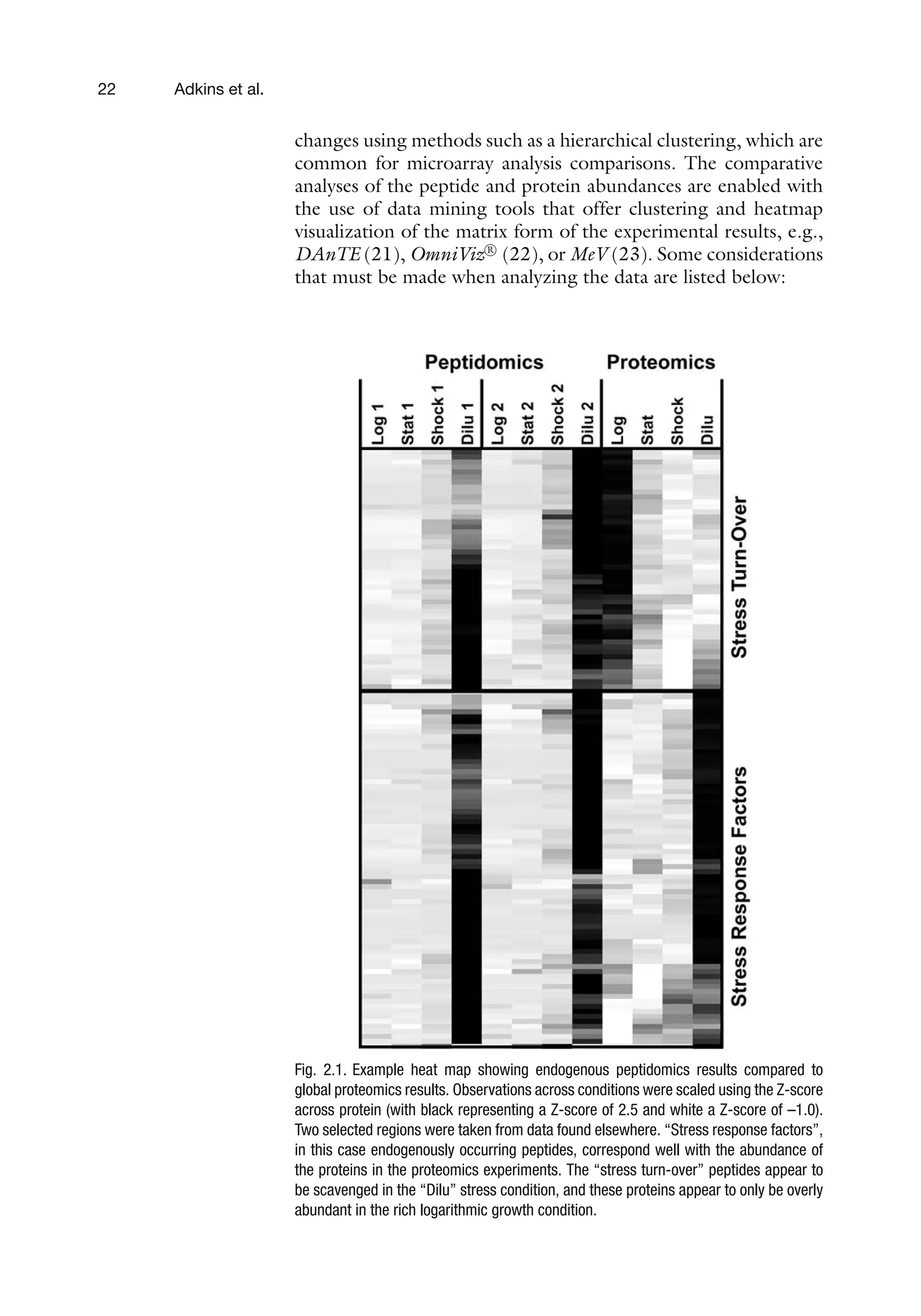







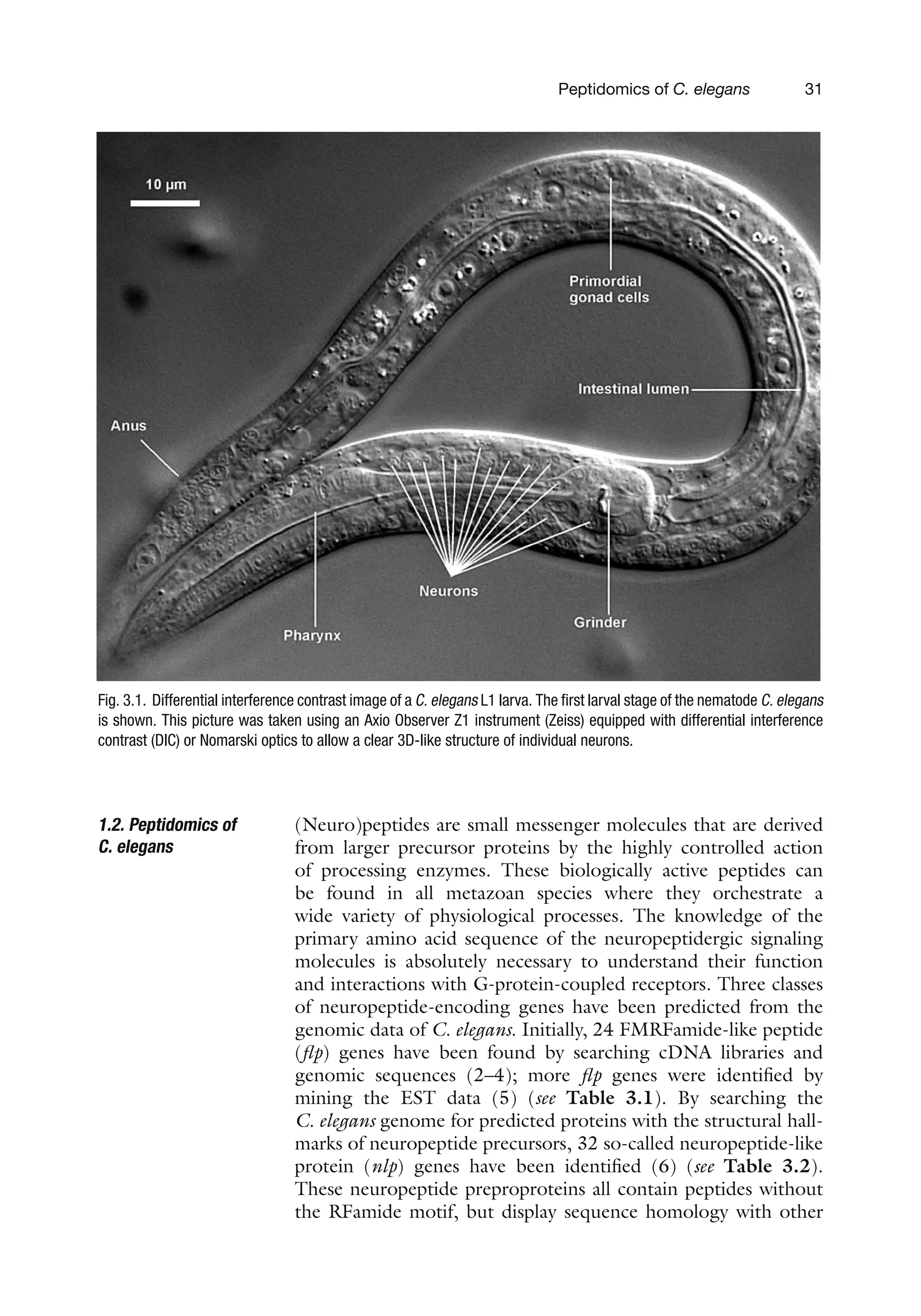
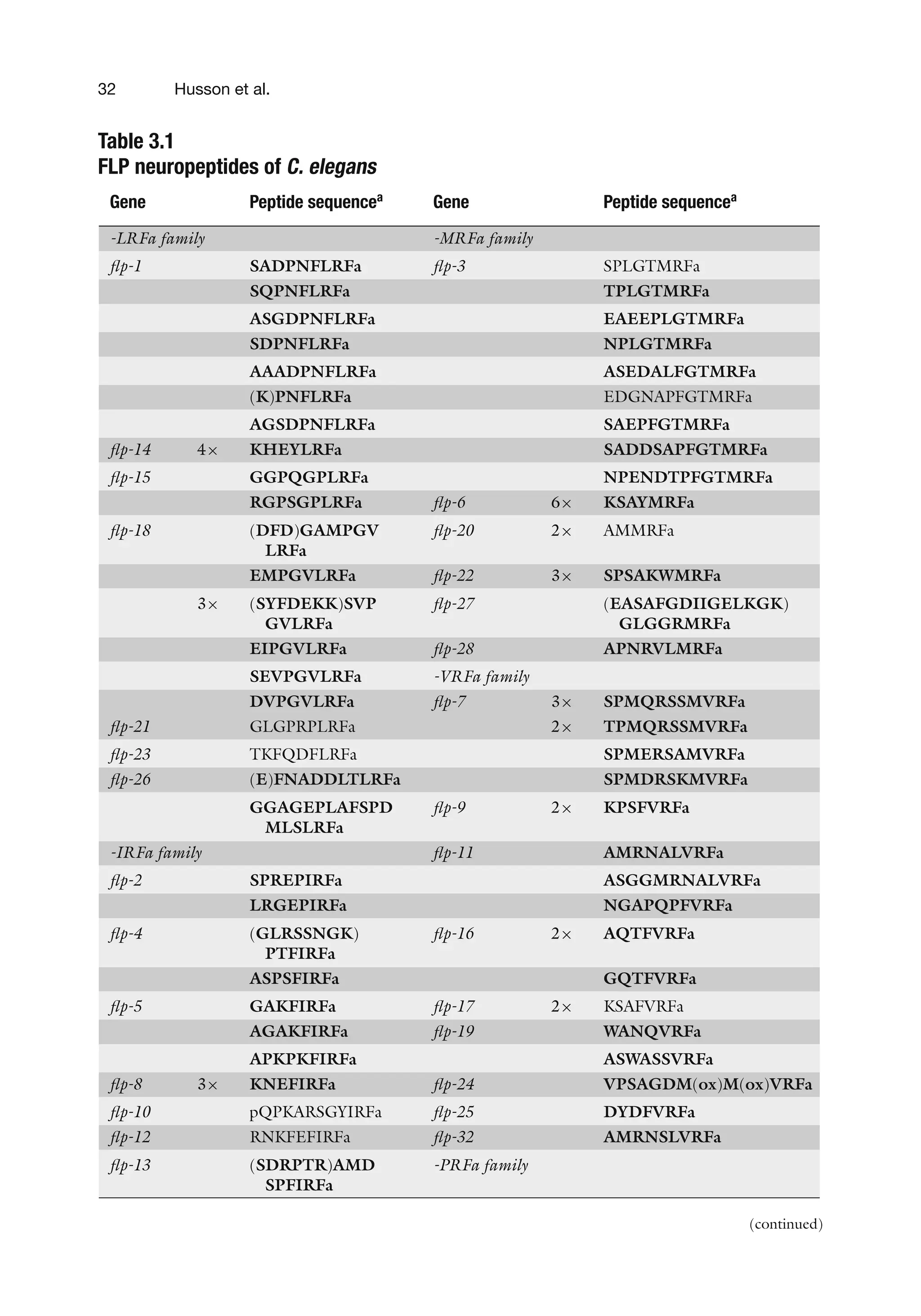
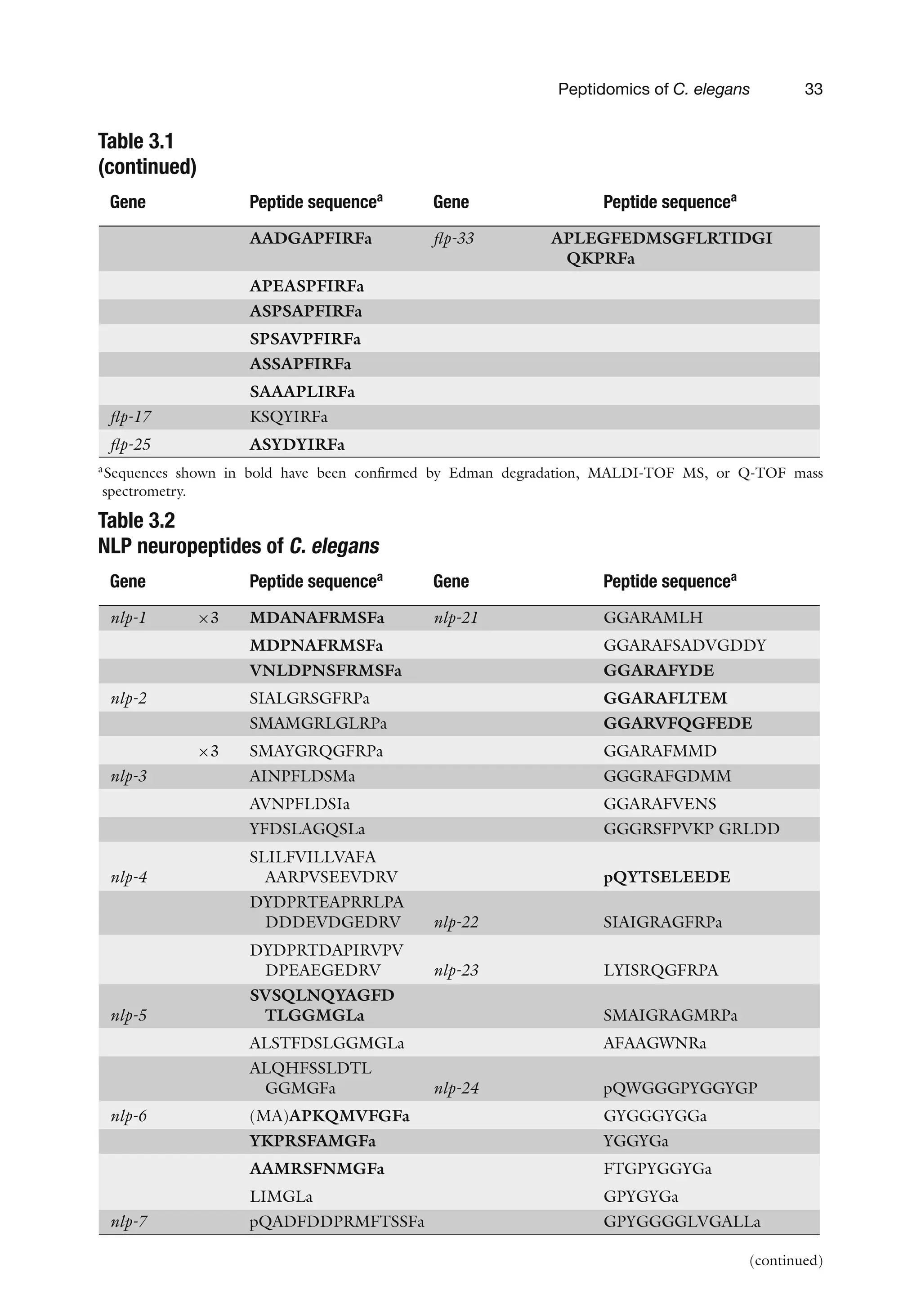
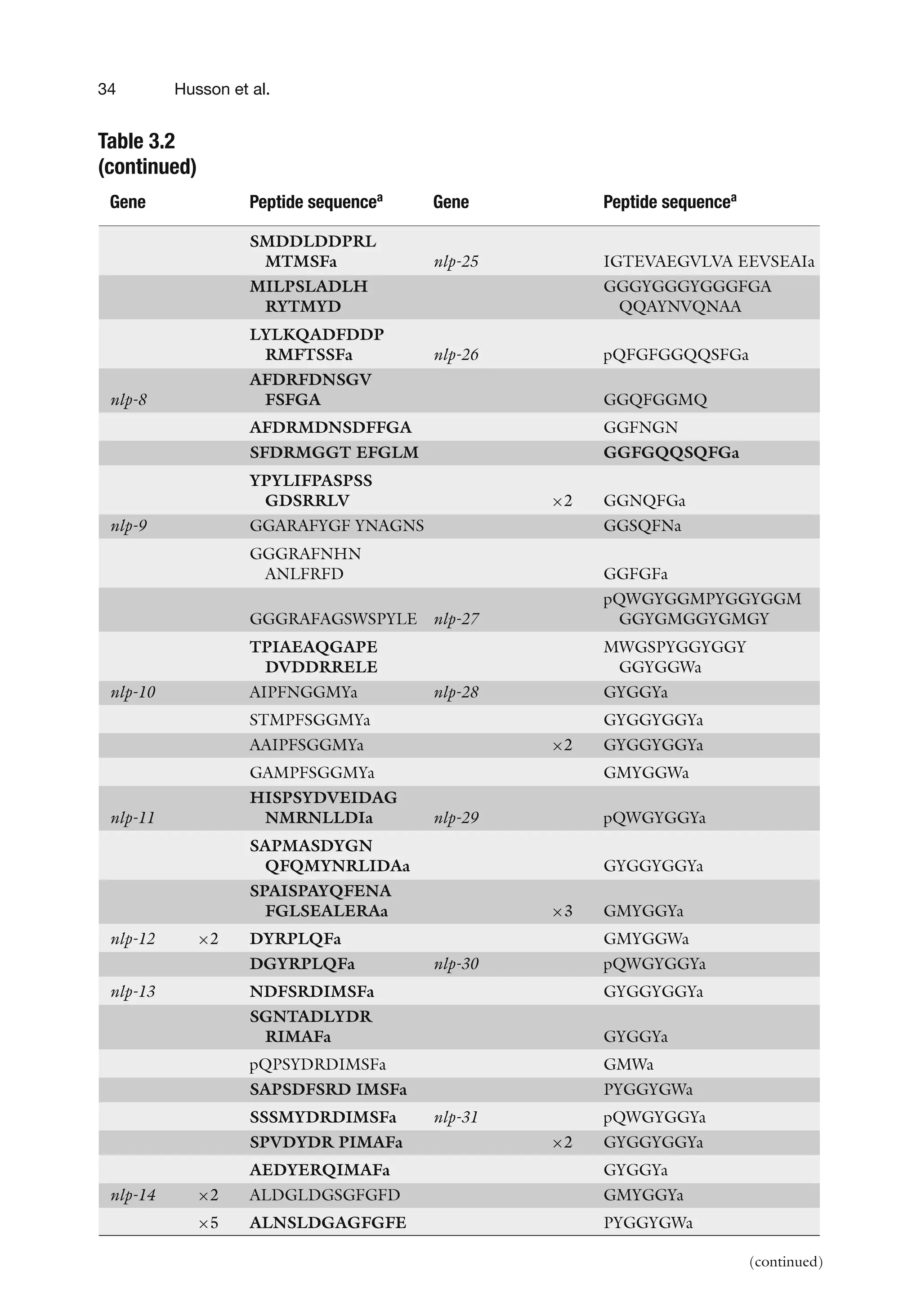
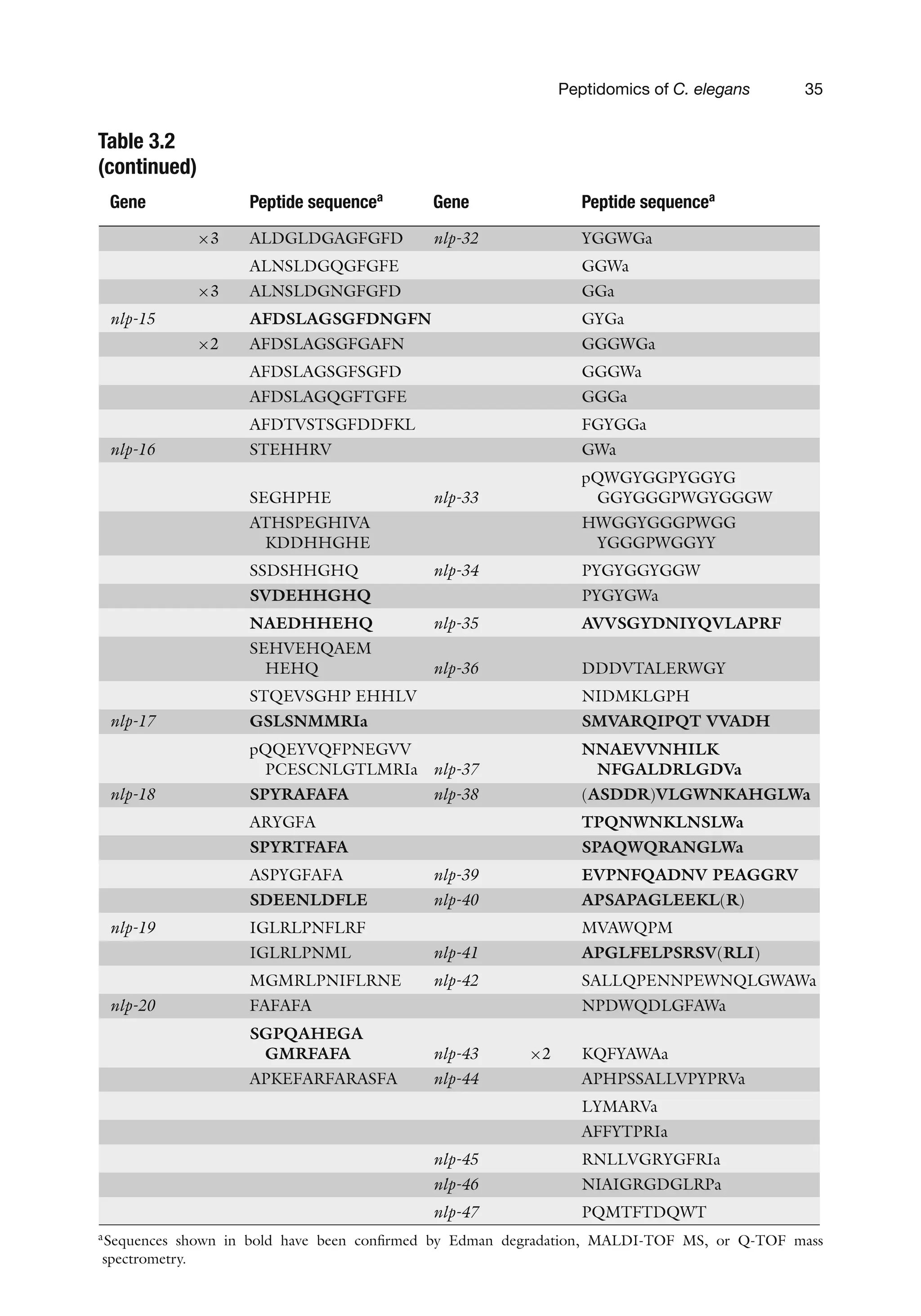








![+ + –
+ –
+
+
–
Spec. 97: 58. Jl. 14, ’06. 2040w.
Olmsted, Stanley, Nonchalante. †$1.25. Holt.
Student life, especially the life of two American students in a
German university town, is cleverly handled in this story, and the
nonchalant heroine, with musical aspirations, is well suited to her
surroundings. The book presents a phase, a passing episode,
interesting and amusing, but superficial in that it deals with that
frivolous side of things which is so typical of student days. The
cafés, the theatres, the bleak boarding houses are well drawn, and
poor Fraulein Mittelini’s tragic struggle for fame is really worthy of
sympathy.
“The author has succeeded ... in giving [the heroine] some
genuine fascination. The style is too obviously imitative of that of
Mr. James.”
Critic. 48: 476. My. ’06. 50w.
“The grip of the book is the grip of Miss Bilton—but it is
entertaining even when she is off the stage.”
N. Y. Times. 11: 287. My. 5, ’06. 540w.
N. Y. Times. 11: 388. Je. 16, ’06. 160w.
R. of Rs. 33: 758. Je. ’06. 100w.
Oman, Charles William Chadwick. Inaugural lecture on the
study of history delivered on Wednesday, Feb. 7, 1906. *35c.
Oxford.
In this lecture on the teaching and study of history the Chichele
professor “perceives the great virtues of the tutorial system. He
recognizes a fact which is often overlooked by zealous reformers,
that no system of teaching can flourish which does not meet the
wants of the learners; and this general truth is in a particular sense
applicable to the universities of England.... The fact ‘that must be](https://image.slidesharecdn.com/34348-250226191938-6b836bd8/75/Peptidomics-Methods-and-Protocols-1st-Edition-Mikhail-Soloviev-Auth-58-2048.jpg)









![+
+
+ +
+
+
+
+ –
+
Reviewed by Erl B. Hulbert.
Am. J. Theol. 10: 362. Ap. ’06. 90w.
“The task has been performed with equal loyalty and ability, and
the book is every way a fitting memorial of a man of great gifts,
high character, and broad influence.”
Critic. 48: 285. Mr. ’06. 60w.
“Mr. Osborn’s biography, in a word, is a worthy memorial of a
great Christian and a great American, and a book which should
enlarge the horizon and stimulate to a higher life all into whose
hands it falls.”
Lit. D. 32: 492. Mr. 31, ’06. 430w.
“The only criticism to be brought against this biography is that
the index is extravagant in its dimensions.”
N. Y. Times. 11: 14. Ja. 13, ’06. 520w.
Outlook. 81: 1039. D. 23, ’05. 160w.
R. of Rs. 33: 253. F. ’06. 90w.
Osbourne, Lloyd. Motormaniacs. [+]75c. Bobbs.
“Pretty little stories they are too, when we are permitted to pause
and enjoy them, and the motormaniacs are always entertaining
and capital company to the end of the run.”
Acad. 71: 399. O. 20, ’06. 110w.
“The dialogue is comic, and the narrative runs with a swing and
zest which are valuable aids to easy reading.”
Ath. 1906, 2: 545. N. 3. 100w.
“The book is full of humour and energy.”
Spec. 97: 497. O. 6, ’06. 110w.](https://image.slidesharecdn.com/34348-250226191938-6b836bd8/75/Peptidomics-Methods-and-Protocols-1st-Edition-Mikhail-Soloviev-Auth-68-2048.jpg)




























































