INTRODUCTION
• Each eukaryoticchromosome is made by linear ds DNA molecule.
• Eukaryotes have from 2 – 15 times as many genes as E. coli.
• This many times the amount of DNA is packaged in several chromosomes and each
chromosome is present in 2 (diploid) or more (polyploidy) copies.
• Contour length of E. coli chromosome is about 1.5 mm, while haploid chromosome
complement ( genome) of human contains about 1000 mm DNA (i.e.2000 mm in diploid
per diploid cell).
• This 1000 mm DNA is divided among 23 chromosomes of variable size and shape, each
chromosome containing from15-85 mm of DNA.
• How this 85 mm (i.e., 85,000 µm) of DNA in the largest human chromosome get
condensed into a mitotic metaphase chromosome structure that is about 0.5 µm in
diameter and 10 µm in length???
• This DNA molecule somehow gets packaged into a metaphase structure that is about
0.5 µmin diameter and about 10 µm in length—a condensation of almost 104
-fold in
length from the naked DNA molecule to the metaphase chromosome.
3.
COMPOSITION OF EUKARYOTICCHROMOSOME
• During interphase of diving cells and G0 of non dividing cells, DNA exist as
nucleoprotein complex called Chromatin, less condensed structure.
• While during M-Phase it gets condensed into chromosome.
• Metaphase chromosome stage is highly condensed DNA and so shortest and
thickest.
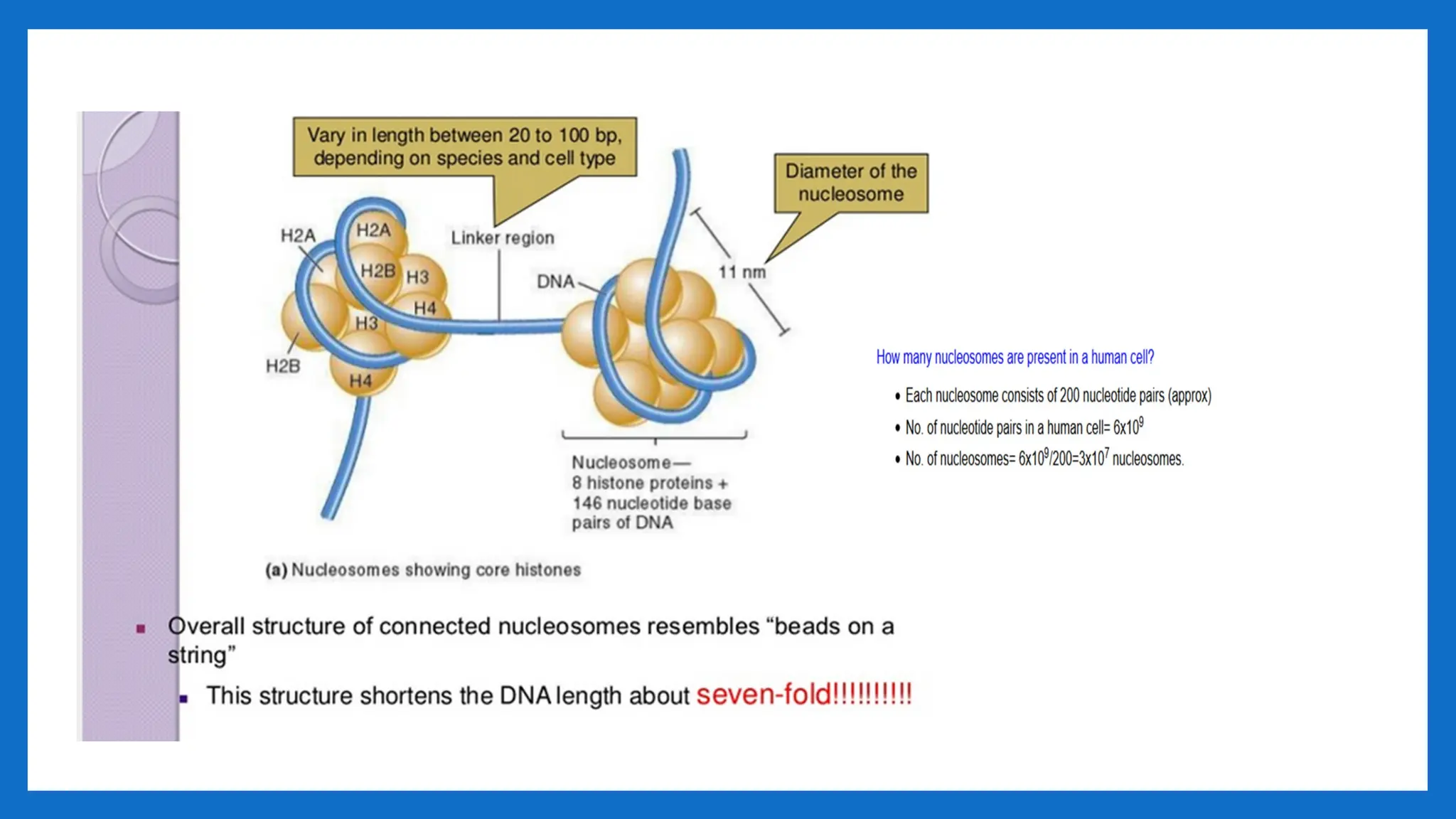
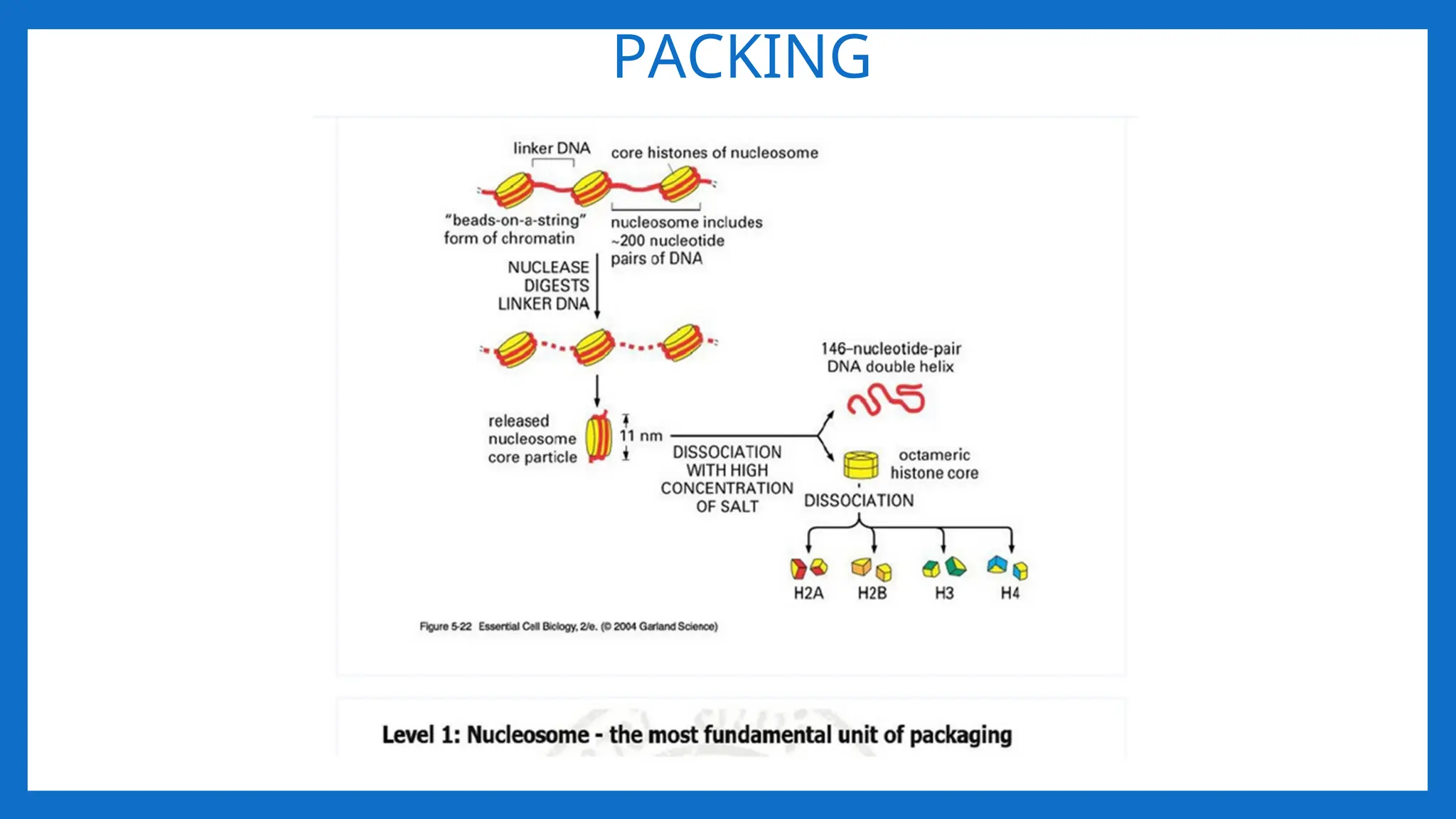
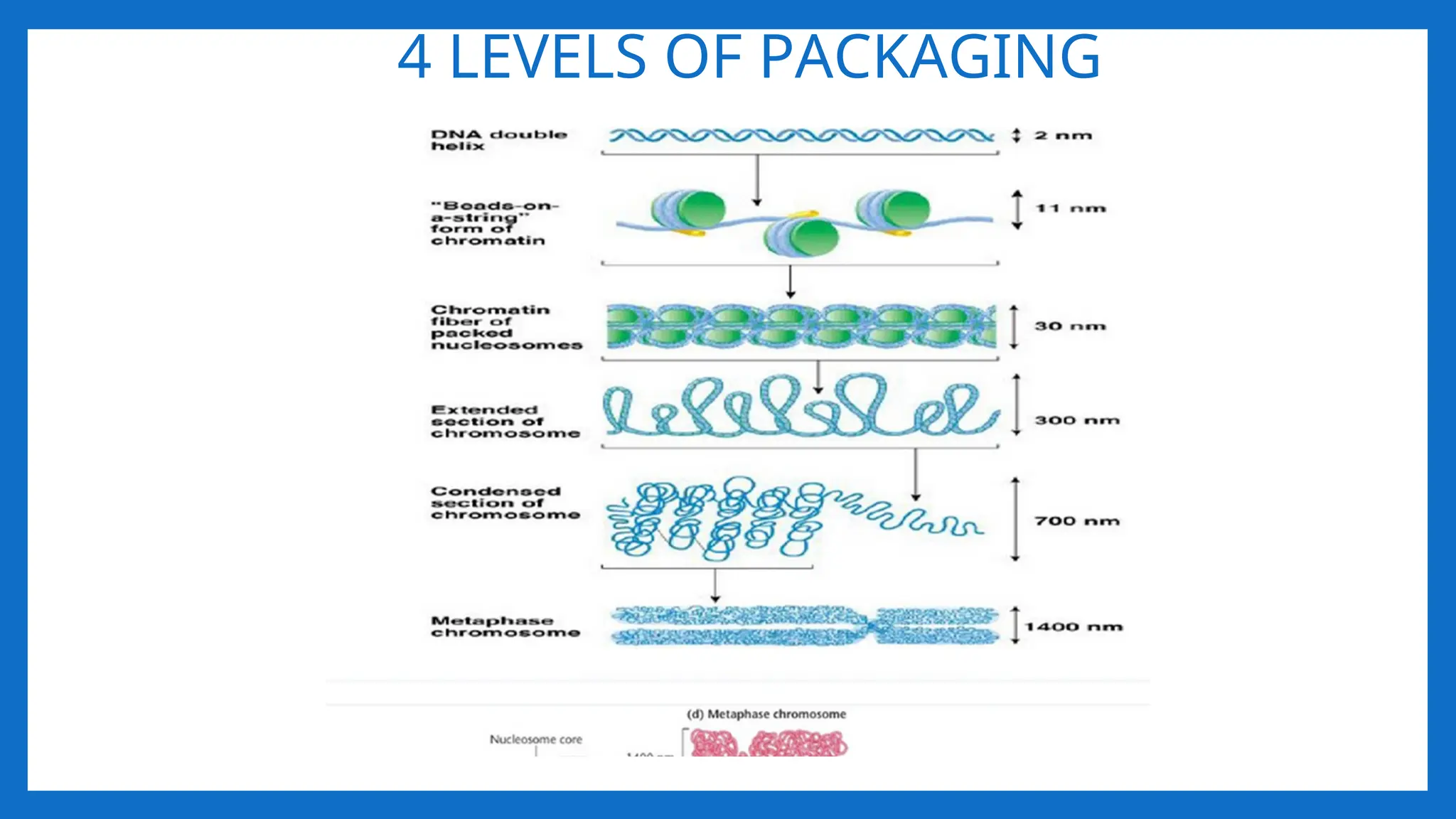
I LEVEL OFPACKAGING 2nm – 11- nm
• Proposed by R.D. Kornberg and J.O. Thomas in 1974.
• According to this theory, Nucleosome was considered as repeating unit (subunit) of
chromatin.
• Adjacent nucleosomes are joined by Linker DNA
• Each nucleosome is associated with one histone H1
• The term nucleosome was given by P. Outdet in 1975
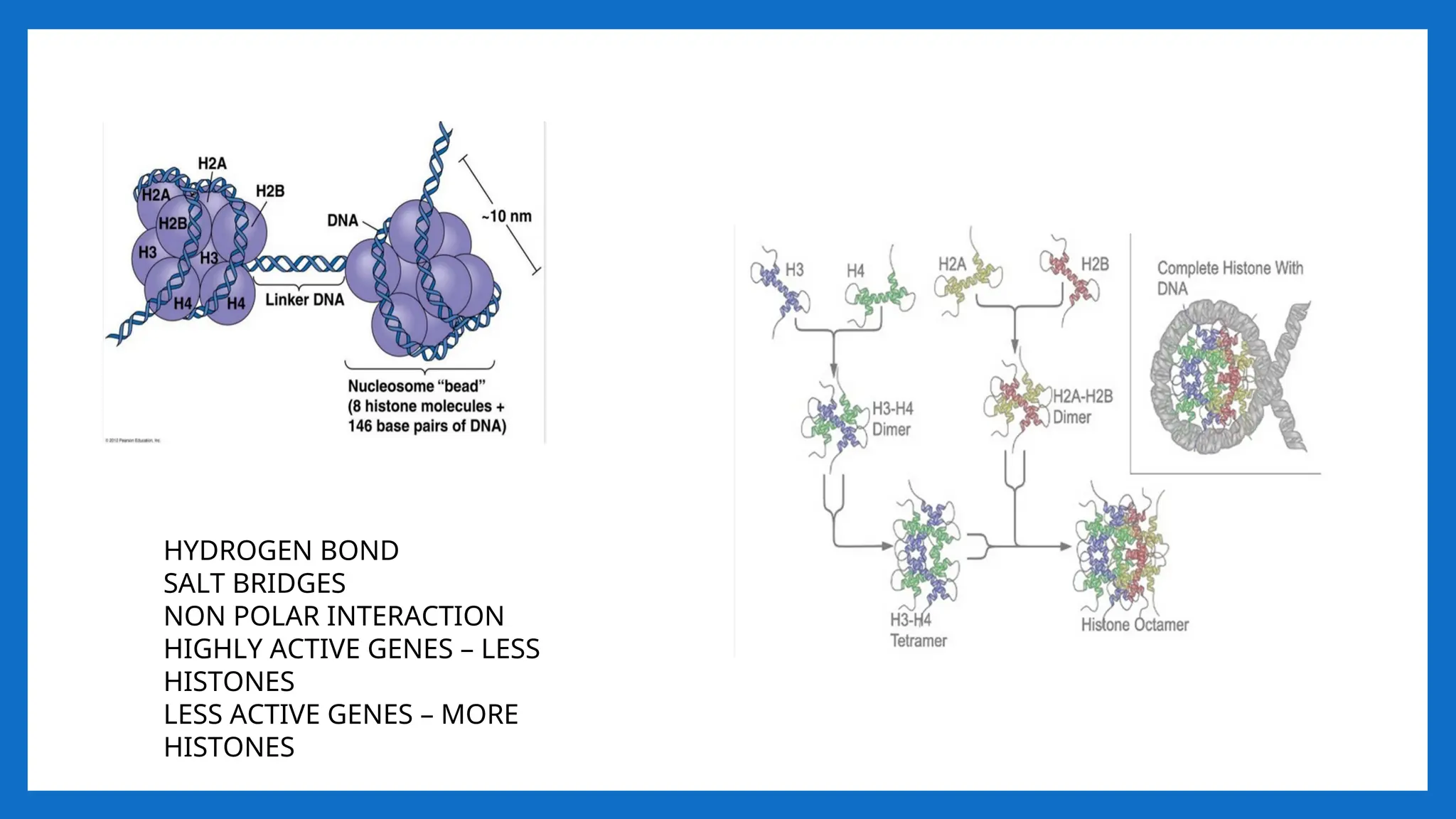
• NUCLEOSOME = 2(H2A H2B, H3, H4) +200 bp DNA + H1
• It is a octamer of histones and 200 bpDNA •
• [H3, H4] tetramer makes a central kernel of nucleosome . (H2A-H2B) dimer are
placed at opposite faces of tetramer.
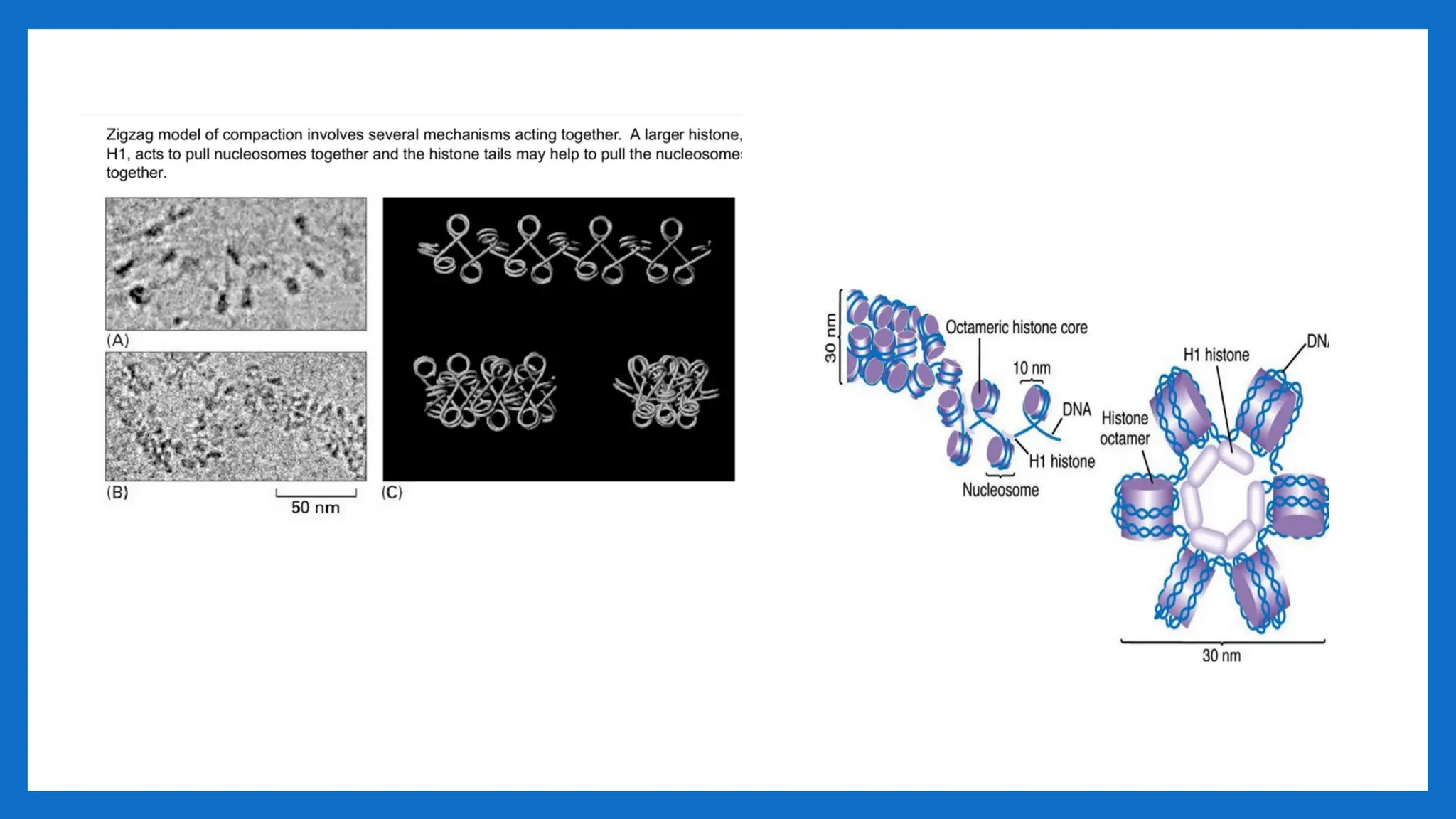
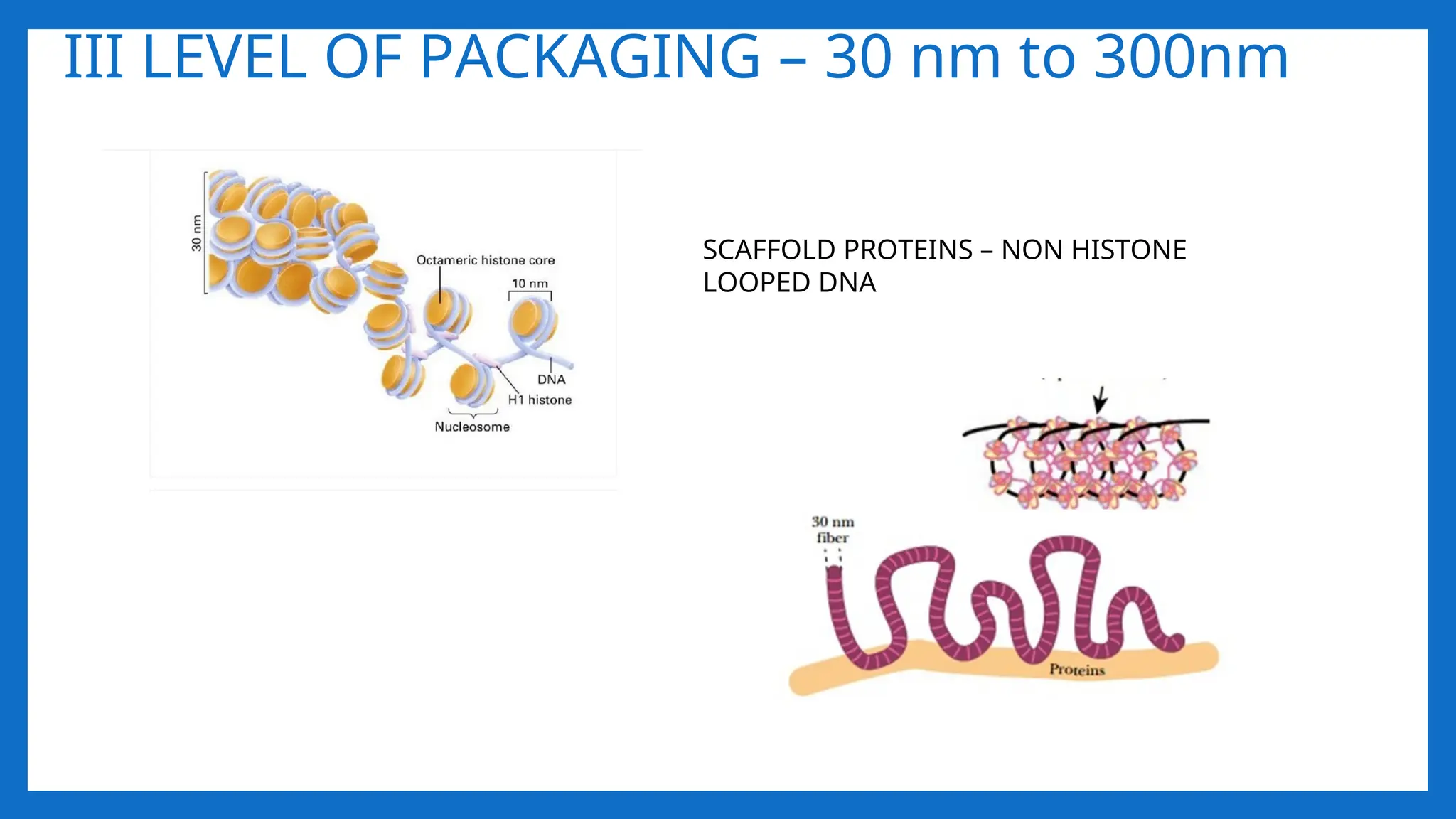
II LEVEL OFPACKAGING 11nm – 30- nm
• Involves further folding or supercoiling of 11nm nucleosome fibre into 30 nm
chromatin fibre, each turn consisting of 6 nucleosmes. • Involves HI • G2 of
interphase.
• As chromosomes enter mitosis the beaded string undergoes higher-order packing.
• The beaded string coils to form the 30-nm chromatin fiber.
• This is second level of packaging
III LEVEL OFPACKAGING – 30 nm to 300nm
SCAFFOLD PROTEINS – NON HISTONE
LOOPED DNA
13.
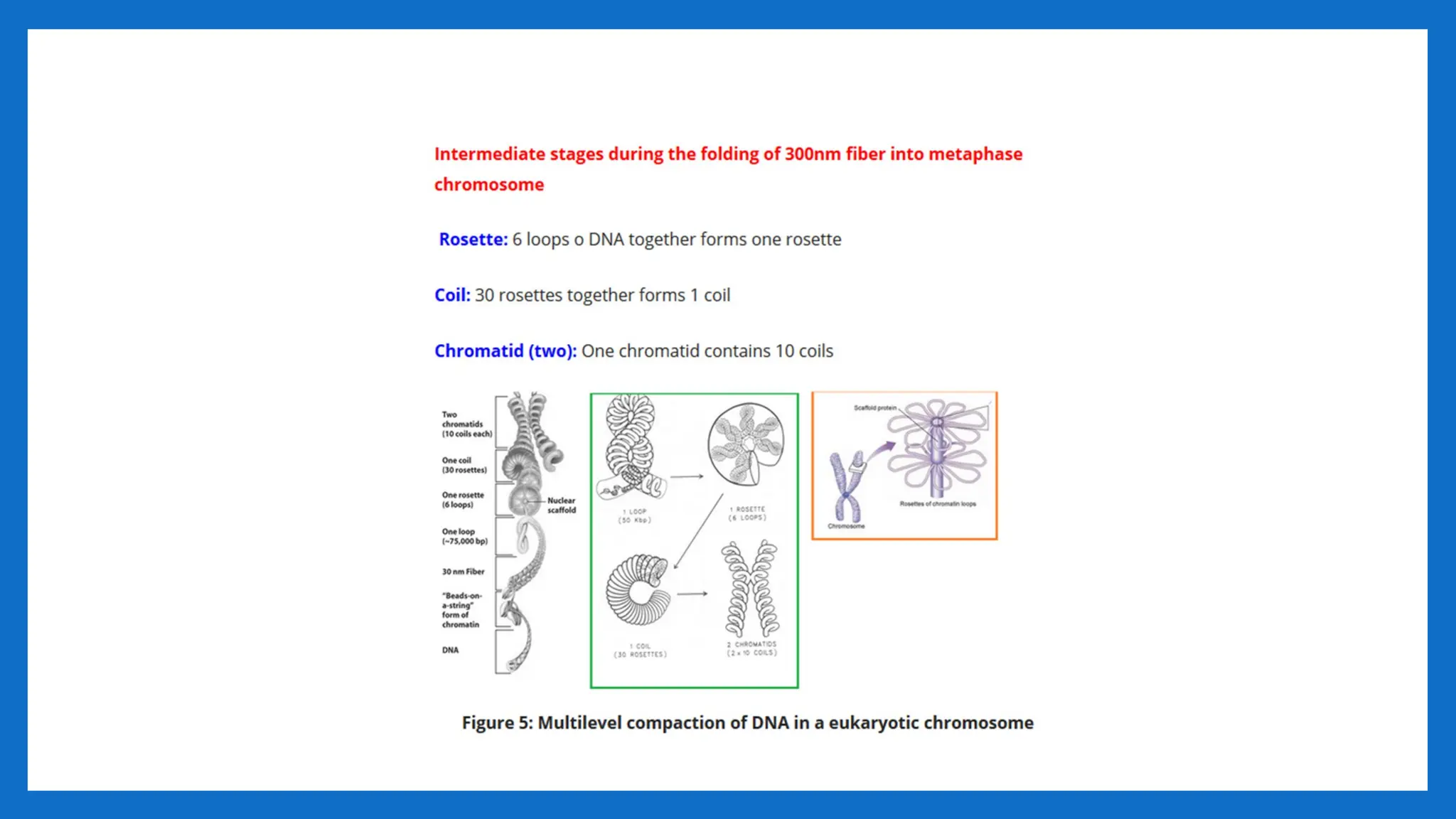
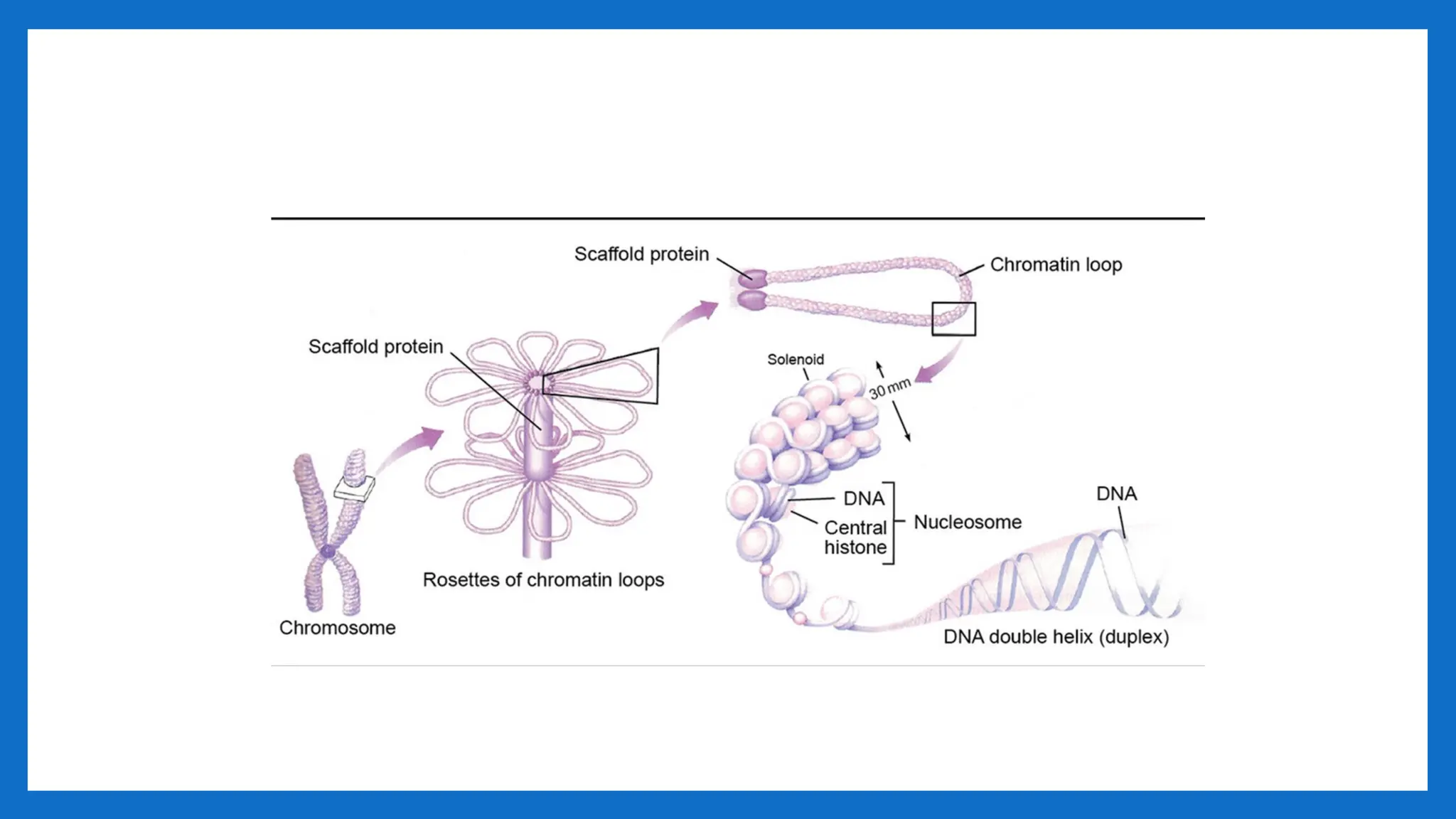
LOOPS, DOMAINS, SCAFFOLDS
•The 30 nm fibres in the form of a solenoid is organized in structures at a higher level
of organization.
• These higher levels of organizations have been examined using metaphase and
interphase chromosomes.
• Folding of the 30-nm fiber form looped domains that attached to a scaffold of non-
histone proteins
• Numerous loops of DNA (30 -90 loops) attached to a protein scaffold. Each loop is a
30nm fiber with 180 -300 nucleosomes, 85 kb of DNA and a length of 10-30 μm
• These loops have actually been observed to emanate from a central scaffold
• Scaffold - The eukaryotic chromosome structure remaining when DNA and histones
have been removed; made from nonhistone proteins. The central framework of a
chromosome to which the DNA solenoid is attached as loops; composed largely of
topoisomerase.
14.
PROTEIN SCAFFOLD
• Thechromatin loop domains are anchored to the matrix by proteins that preferentially bind AT-
rich sequences
• Among the proteins characterized are
• the special AT-rich binding protein 1, SATB1, that is expressed predominantly in thymocytes;
• (2) the 95-kDa attachment region binding protein, (ARBP)
• (3) the A and B lamins of the nuclear envelope,
•
• (4) the nuclear intermediate filament-type protein called NuMA, and
• (5) the hnRNP U protein.
15.
PROTEIN SCAFFOLD contd…
•Most interestingly, topoisomerase II associates with both interphase MARS and
mitotic SARS.
• Topoisomerase II may relieve torsional stress generated by transcription within
individual looped domains during the interphase (as it binds to MARS), and the
enzyme may serve as a structural protein within the chromosome scaffold during
mitosis (as it binds SARS).
16.
Domains on DNA– SARS/ MARS
• In interphase nuclei, a nuclear matrix is found as a filamentous structure on the
interior of nuclear membrane.
•
• Chromatin remains attached to this matrix through matrix attachment regions
(MARs).
• Several MAR sequences have actually been identified. It is believed that the same
sequences of DNA work as MAR in interphase nuclei and as SAR (scaffold
attachment regions) in metaphase chromosomes.
• Both matrix and scaffold are proteinaceous in nature.
17.
• MARS formthe bases of these looped domains and anchor the loops to the mitotic
chromosome scaffolds or to the interphase nuclear matrix.
•
• In mitotic chromosomes, these elements have traditionally been referred to as SARS
for scaffold attachment regions.
• In the case of the interphase matrix, these AT-rich elements are usually called MARS.
18.
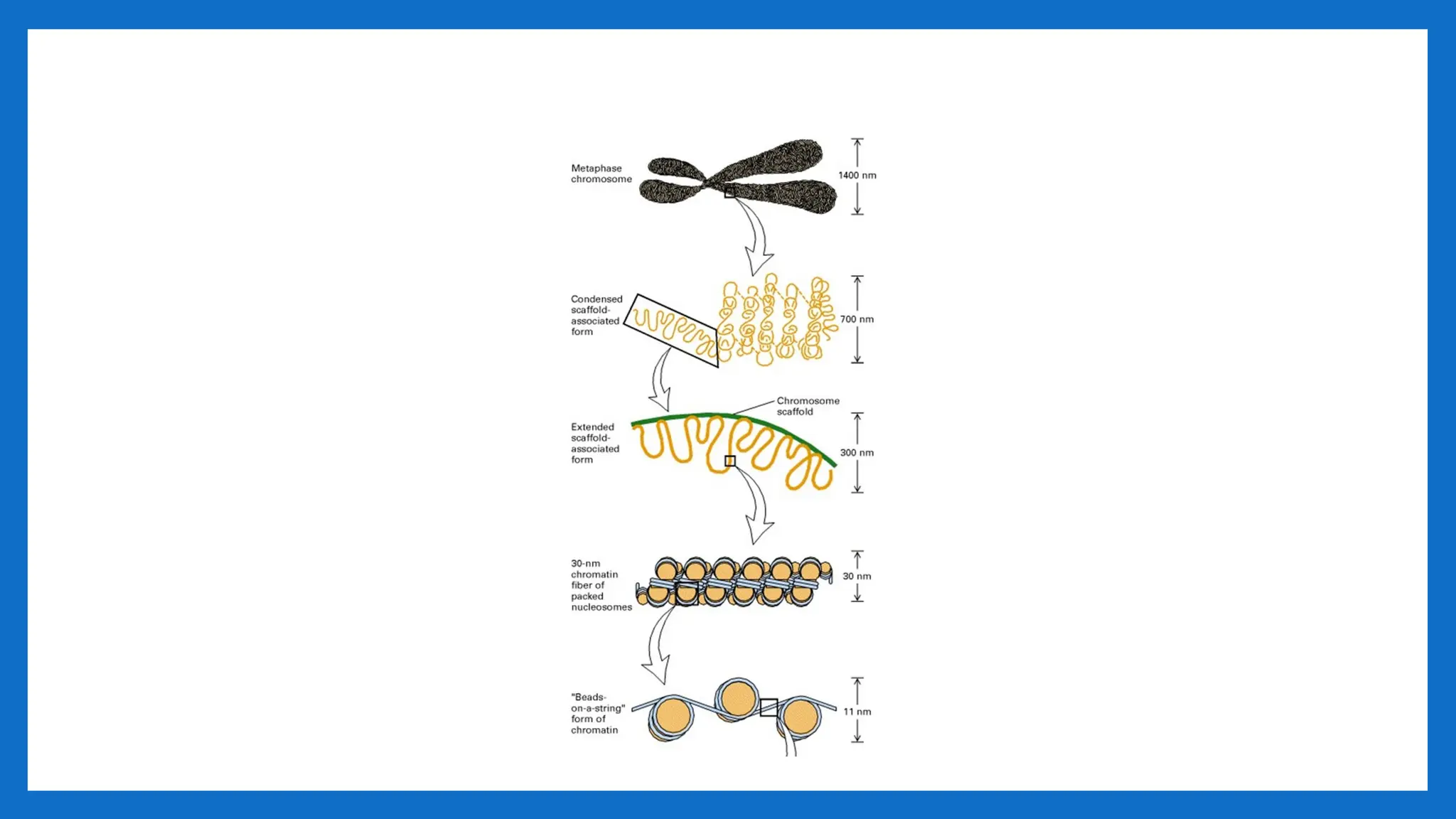
IV LEVEL -300nmto 700nm – 1400nm
• The looped domains (300nm chromatin fiber) coil further during mitosis to form a 700-nm
chromatid (one of the longitudinal subunits of the metaphase chromosome).
• Such tight packing makes the genes on the DNA inactive.
• This structure gives a packaging ratio of ~700
• A pair of sister chromatids comprising a chromosome measures about 1400 nm.
• Fully condensed chromosome is 10,000 fold shorter and 400 – fold thicker than DNA
alone.
• Metaphase chromosomes are the most condensed of normal eukaryotic chromosomes.
• This structure gives a packaging ratio of ~10,000
• The role of these highly condensed chromosomes is to organize and package the giant
DNA molecules of eukaryotic chromosomes into structures that will facilitate their
segregation to daughter nuclei without the DNA molecules of different chromosomes
becoming entangled and, as a result, being broken during the anaphase separation of the
daughter chromosomes.
22.
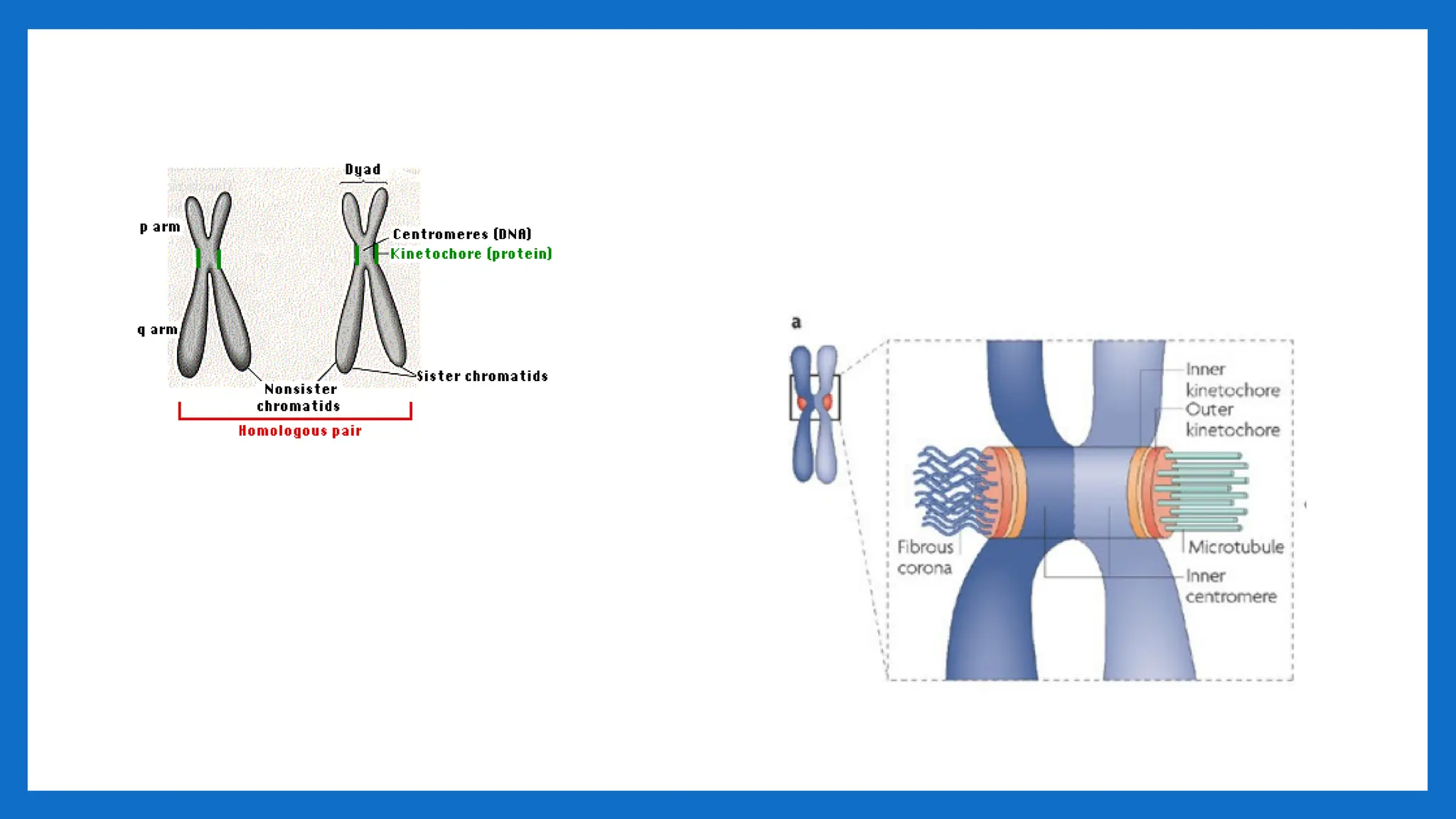
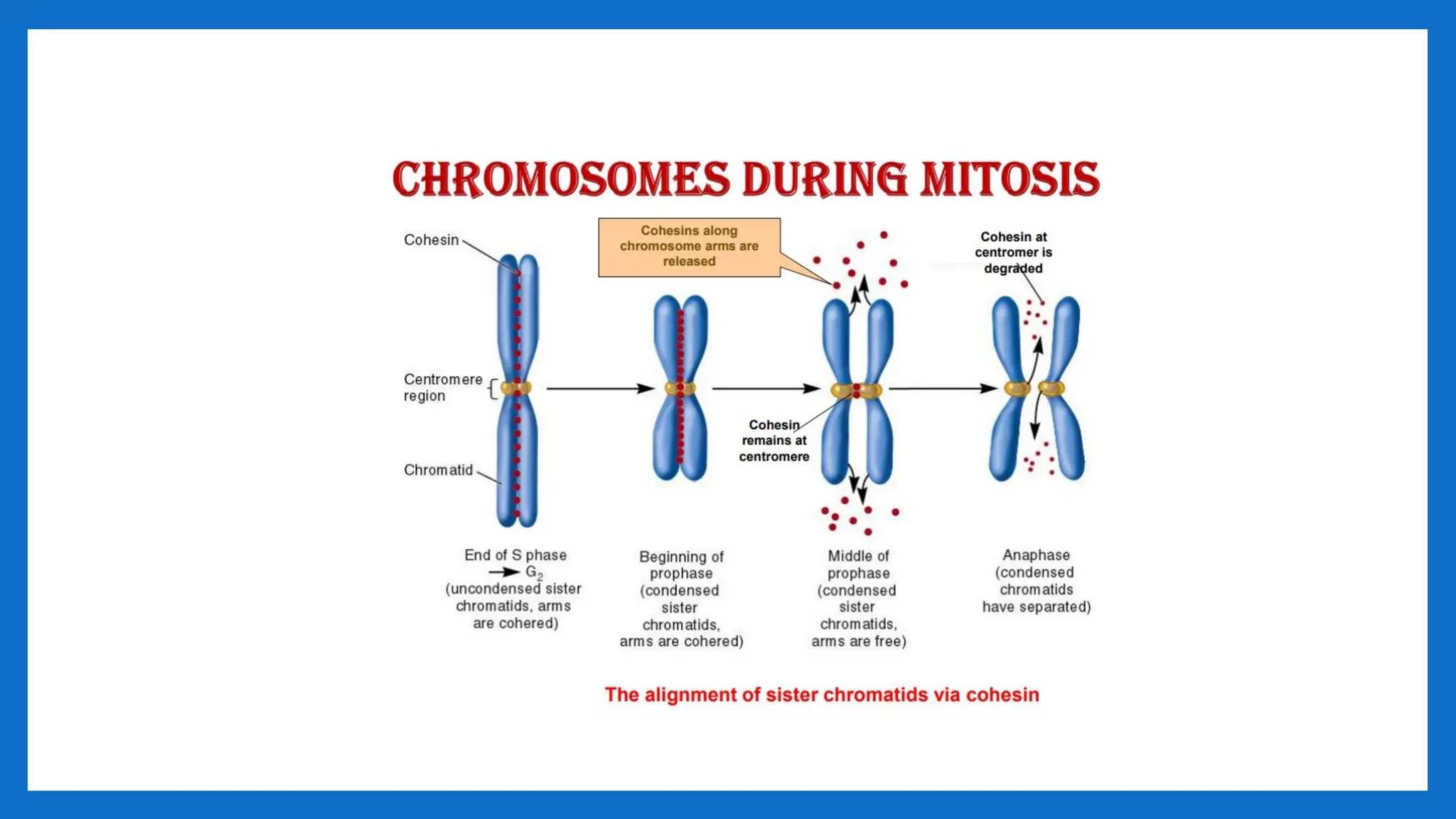
CENTROMERE
• The centromere(centro- + -mere) is the part of a chromosome that links sister
chromatids.
• The word "centromere" is derived from the Greek words centro ("central") and
mere("part")
• During mitosis, spindle fibers attach to the centromere via the kinetochore.
• Centromeres were first defined as genetic loci that direct the behavior of chromosomes.
• The centromere was first described by German biologist Walter Flemming in the 1880s as
the "primary constriction" of the chromosome.
• centromere is a region of specialized chromatin found within each constricted
chromosome that provides the foundation for kinetochore assembly and serves as a site
for sister chromatid attachment
• Errors in centromere or kinetochore function are catastrophic for cells.
• Such errors can lead to aberrant division and chromosomal instability, both of which are
often observed in cancerous cells.
24.
COMPONENT AND FUNCTION
•The centromere usually contains specific types of DNA sequences which are in
higher eukaryotes typically tandem repetitive sequences, often called "satellite
DNA".
• These sequences bind specific proteins called "cen"-Proteins.
• During mitosis the centromeres can be identified in particular during the
metaphase stage as a constriction at the chromosome.
• At this centromeric constriction the two mostly identical halves of the chromosome,
the sister chromatids, are held together until late metaphase.
• During mitotic division, a transient structure called kinetochore is formed on top of
the centromeres.
• The kinetochores are the sites where the spindle fibers attach.
• Kinetochores and the spindle apparatus are responsible for the movement of the
two sister chromatids to opposite poles of dividing cell nucleus during anaphase.
26.
TELOMERS
• Telomeres aredistinctive structures found at the ends of our chromosomes.
• They consist of the same short DNA sequence repeated over and over again.
• In humans the telomere sequence is TTAGGG.
• This sequence is usually repeated about 3,000 times and can reach up to 15,000
base pairs in length.
27.
• These endsequences are protected by telomeric proteins TRF1 and TRF2
• While replicating DNA, the eukaryotic DNA replication enzymes cannot replicate the
sequences present at the ends of the chromosomes.
• Hence, these sequences and the information they carry may get lost.
• Telomeres "cap" the end-sequences and themselves get lost in the process of DNA
replication.
28.
TELOMERE AND CELLDIVISION
• Two main factors contribute to telomere shortening during cell division :
• The “end replication problem” during DNA replication: Accounts for the loss of about
20 base pairs? per cell division.
• Oxidative stress: Accounts for the loss of between 50-100 base pairs per cell division.
The amount of oxidative stress in the body is thought to be affected by lifestyle
factors such as diet, smoking and stress.
• When the telomere becomes too short, the chromosome reaches a 'critical length'
and can no longer be replicated.
• This 'critical length' triggers the cell to die by a process called apoptosis?, also
known as programmed cell death.
29.
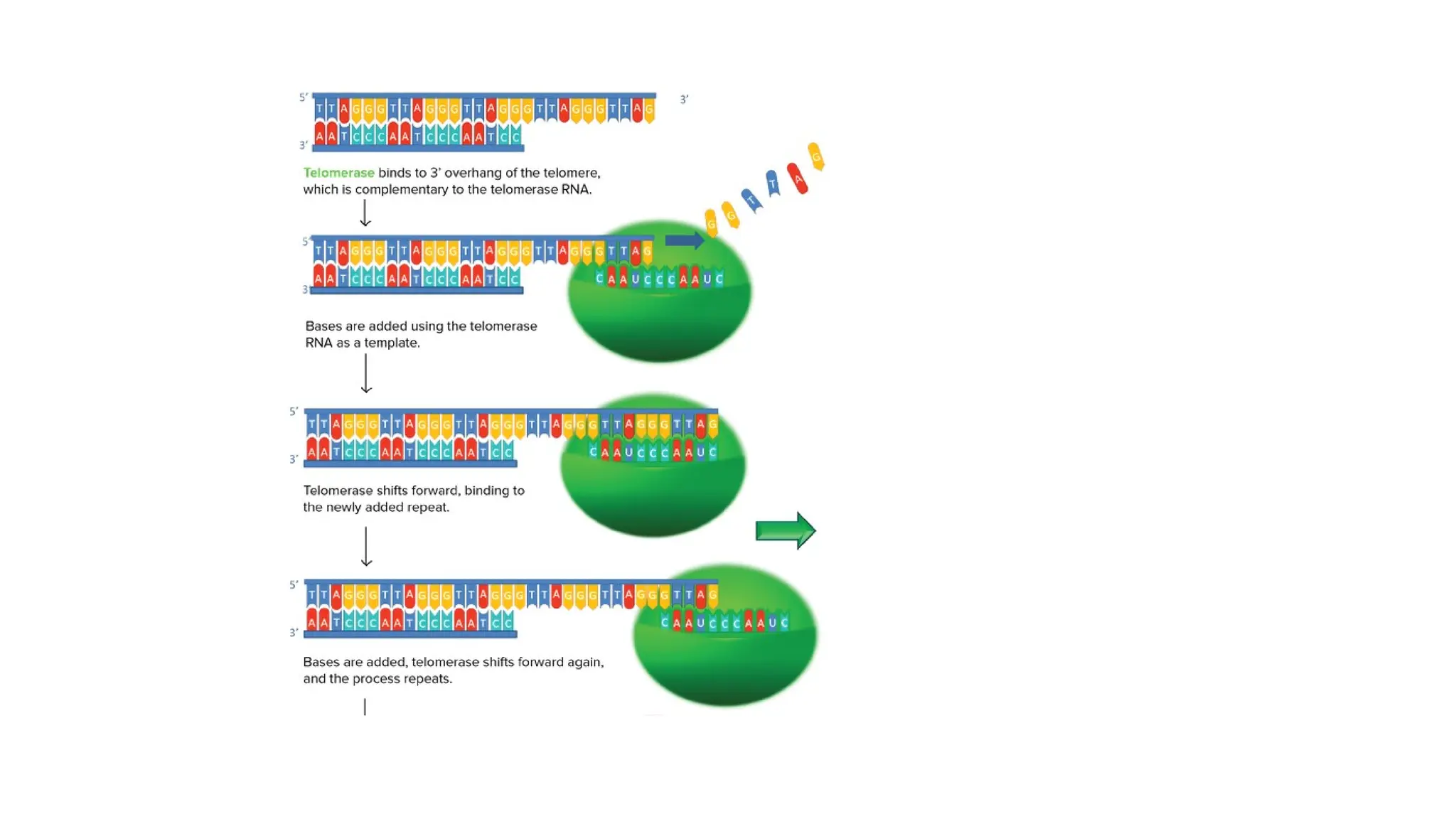
How telomere lengthsare maintained
Telomeric sequence is maintained by a special enzyme called
telomerase.
• Specialized reverse transcriptase.
• Carries its own template RNA, which is complementary to
telomeric repeat sequences.
30.
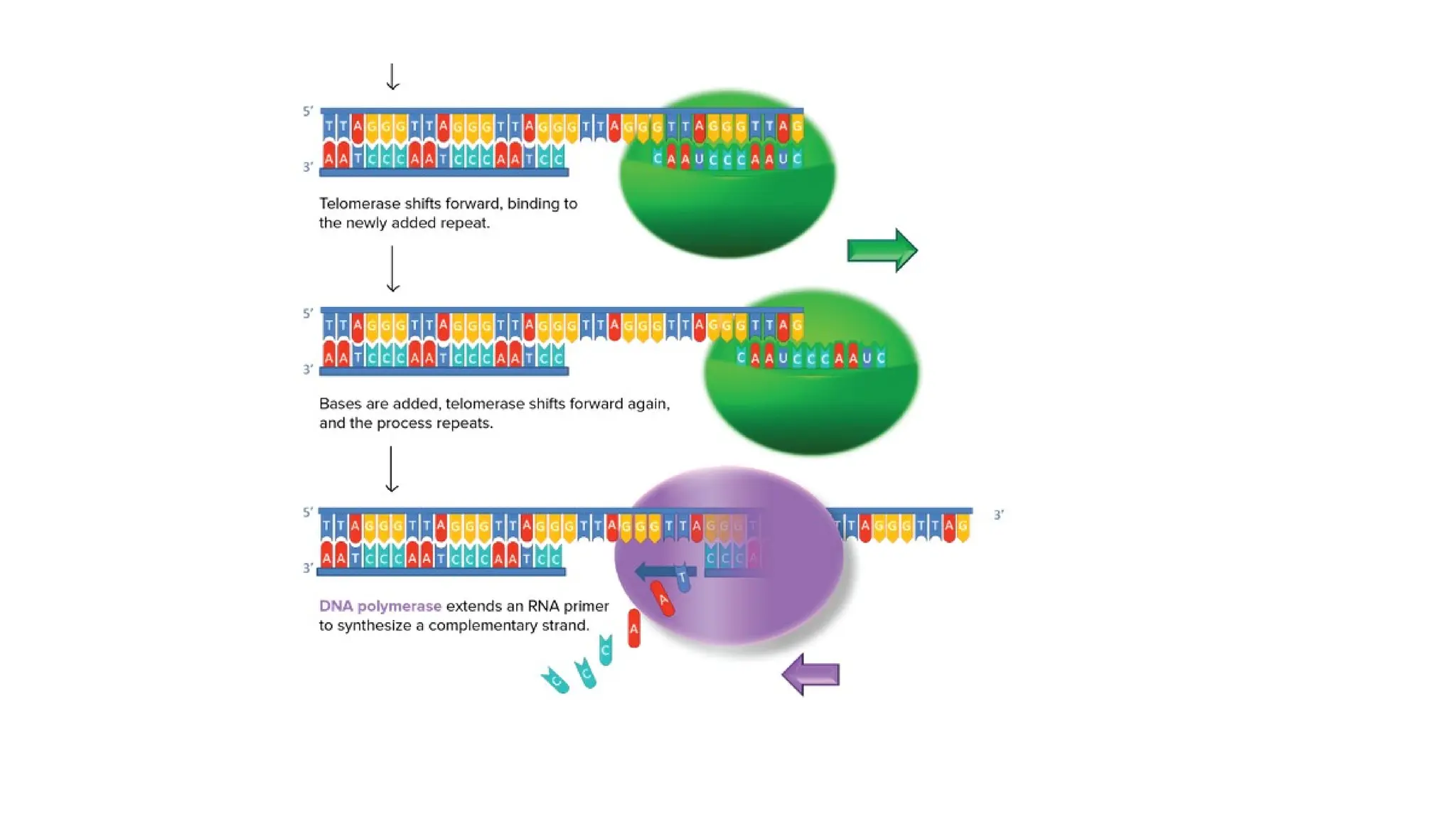
TELOMERASE
• Telomerase isan RNA-dependent DNA polymerase, meaning an enzyme that can
make DNA using RNA as a template.
• The enzyme binds to a special RNA molecule that contains a sequence
complementary to the telomeric repeat.
• It extends (adds nucleotides to) the overhanging strand of the telomere DNA using
this complementary RNA as a template.
• When the overhang is long enough, a matching strand can be made by the normal
DNA replication machinery (that is, using an RNA primer and DNA polymerase),
producing double-stranded DNA.
• The primer may not be positioned right at the chromosome end and cannot be
replaced with DNA, so an overhang will still be present. However, the overall length
of the telomere will be greater.
33.
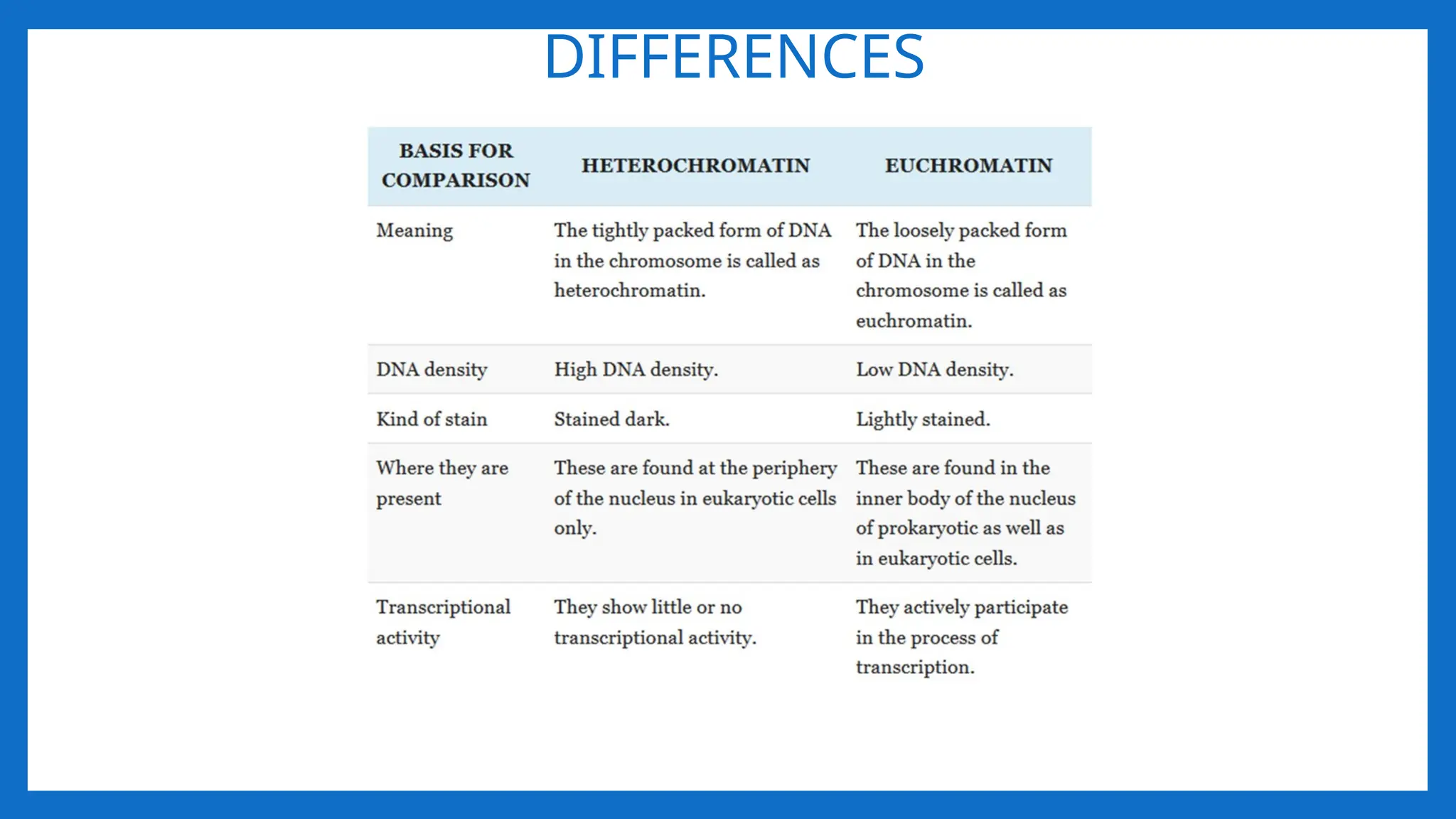
EUCHROMATIN AND HETEROCHROMATIN
•Interphase chromosomes have areas that remain highly condensed,
heterochromatin, and less compacted areas, euchromatin.
• The term Heterochromatin and Euchromatin was coined by Emil Heitz in 1928.
• Heterochromatin and Euchromatin are the parts of the chromatin.
• DNA protein complex found in the eukaryotes.
• These were take part in the protection of DNA inside the nucleus.
34.
HETEROCHROMATIN
• The regionsof the chromosome that appear relatively condensed and stained
deeply with DNA specific strains.
• It is tightly packed form of DNA.
• There are two types of heterochromatin, Constitutive heterochromatin and
Facultative heterochromatin.
• Both of the constitutive heterochromatin and facultative heterochromatin play a
role in the expression of genes.
• Transcriptionally inactive.
• Facultative heterochromatin is the result of genes that are silenced through a
mechanism such as Histone methylation or siRNA through RNAi.
• Constitutive heterochromatin is usually repetitive and forms structural functions
such as centromeres or telomeres.
35.
EUCHROMATIN
• Euchromatin isthe lightly packed form of chromatin that is rich in gene
concentration.
• It is often under active transcription.
• Euchromatin comprises the most active portion of the genome within the nucleus,
92% of the human genome is euchromatic.
• The structure of Euchromatin is reminiscent of an unfolded set of beads represent
Nucleosomes, Nucleosomes consist of eight proteins known as Histones, with
approximately 147 base pairs of DNA wound around them.
• In Euchromatin the wrapping is loose so that the raw DNA may be accessed.
• The basic structure of Euchromatin is an elongated, open 10nm micro fibril, as
noted by electron microscopy.
• Euchromatin participates in the active transcription of DNA to mRNA products.



![I LEVEL OF PACKAGING 2nm – 11- nm
• Proposed by R.D. Kornberg and J.O. Thomas in 1974.
• According to this theory, Nucleosome was considered as repeating unit (subunit) of
chromatin.
• Adjacent nucleosomes are joined by Linker DNA
• Each nucleosome is associated with one histone H1
• The term nucleosome was given by P. Outdet in 1975
• NUCLEOSOME = 2(H2A H2B, H3, H4) +200 bp DNA + H1
• It is a octamer of histones and 200 bpDNA •
• [H3, H4] tetramer makes a central kernel of nucleosome . (H2A-H2B) dimer are
placed at opposite faces of tetramer.](https://image.slidesharecdn.com/organisationofeukaryoticchromososme-251112065113-9c549511/75/ORGANISATION-OF-EUKARYOTIC-CHROMOSOSME-ppt-5-2048.jpg)