1. Retroviruses package two copies of its unspliced RNA
genome into their viral particles. It was reported that
retroviral genomes in the virus particle are non-
covalently dimerized in the 5’ untranslated region (UTR).
Genome selection is mediated by RNA packaging
elements, called -sites, which are typically located
within the 5’UTR. -sites generally overlap with the
dimerization site, implicating that retroviral genome
packaging and genome dimerization are coupled.
Background
Conclusion
Materials and Methods
Introduction
IMPACT OF DYNAMICS OF HIGHLY CONSERVED RETROVIRAL RNA PACKAGING
ELEMENT ON GENOME DIMERIZATION AND NUCLEOCAPSID BINDING
Gerardo Camarena Gomez, Sabrina Ngo, Trevor Mathias, Yasuyuki Miyazaki, Michael Summers
Figure 4| Evidence for two alternate conformations of SL-C.
On the left, portion of the 2D NOESY spectrum obtained for SL-C
hairpin RNA, showing chemical exchange cross peaks that
correlate protons of the three distinct SL-C conformations (shown
at top). Note that a kissing interaction forms when SL-C harbors
a GACG tetraloop. On the right, NOSEY spectrum of SL-C
construct containing U328A/A333U mutations.
1. D’Souza, V. & Summers, M. F. (2005). How
Retroviruses Select Their Genomes. Nature 3, 643-
655.
2. Miyazaki, Y., et al. Structure of a Conserved Retroviral
RNA Packaging Element by NMR Spectroscopy and
Cryo-Electron Tomography. (Unpublished).
Dr. Michael Summers, Dr. Yasuyuki Miyazaki. This work is
supported by NIH-ARRA 3R37HI030917-1951 (M.F.
SUMMERS) and Howard Hughes Medical Institute at
UMBC.
References
Acknowledgements
We have proposed that the conformational change of
the -site induced by dimerization exposes nucleocapsid
(NC)- binding sites. In the Moloney Murine Leukemia
Virus (MoMuLV), the core-encapsidation signal (CES)
was determined as the minimum region required for
genome packaging. CES is composed of stem loops
DIS-1, DIS-2, SL-C, and SL-D. DIS-1 and DIS-2 harbor
self-complementary sequences and can form extended
dimer conformations while SL-C and SL-D harbor a
GACG tetraloop. A stem loop harboring a GACG
tetraloop can interact by forming an intermolecular CG
base pairing at the loop (kissing interaction). GACG
sequences in SL-C and SL-D are highly conserved
among all -retroviruses, implicating its biological
significance. Although it was hypothesized that SL-C and
SL-D promote dimerization via kissing interactions, the
function of SL-C and SL-D remains unclear.
Figure 2|RNA Secondary Structure for Moloney Murine Leukemia Virus
Genome Packaging. a| Representation of the Moloney Murine Leukemia
Virus genome, showing the relative locations of the coding and non-coding
elements. b| Secondary structure predicted for the MoMuLV -site, showing
the location of the dimerization initiation site (DIS), SL-C, and SL-D.
c| Potential dimerization of DIS-2. Base pairing enables the AGCU segment
(green) to form a kissing interaction with another molecule. The GACG
tetraloop (blue) on SL-C and SL-D also interact to form a kissing complex.
Note that the NC-binding UAUCUG segment (red) is sequestered by base
pairing in the hairpin structure and exposed in the dimer.
• NMR data revealed that SL-C forms two
alternate conformations, where one contains a
GACG tetraloop and the other does not.
• CESC-KISS inhibited dimerization of DIS, while
CESC-NOKISS promoted dimerization.
• CESC-KISS showed tight NC-binding while
CESC-NOKISS has significantly weak NC-binding.
• These results collectively suggest that SL-C
and SL-D serve as a scaffold for NC-binding by
forming a tandem stem loop complex rather
than promoting genome dimerization.
H
H
hH
Figure 3| Structure of CD determined by NMR.
a,b| Orthogonal views of the 20 structures calculated with Cyana
showing the degree of convergence and overall asymmetrical
shape of the dimer. The position of the experimentally
unrestrained U319 residue is labeled. c,d| Stereoscopic view
showing the c| structure of the A-minor K-turn and its proximity to
the kissing interface and d| stacking of the A353 and A354
residues between the lower stems of SL-C and SL-C.
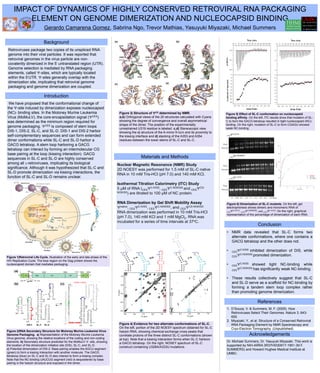
Figure 1|Retroviral Life Cycle. Illustration of the early and late phase of the
HIV Replication Cycle. The blue region on the Gag protein shows the
nucleocapsid domain that mediates packaging.
Figure 5| Effect of SL-C conformation on nucleocapsid
binding affinity. On the left, ITC results show that mutation of SL-
C to form the GACG tetraloop resulted in tight nucleocapsid (NC)-
binding. On the right, mutation of SL-C to form CGAGU showed
weak NC-binding.
Figure 6| Dimerization of SL-C mutants. On the left, gel
electrophoresis shows dimeric and monomeric RNA of
CESNATIVE, CESC-NOKISS, and CESC-KISS. On the right, graphical
representation of the percentage of dimerization of each RNA.
Nuclear Magnetic Resonance (NMR) Study
2D NOESY was performed for 1.5 mM of SL-C native
RNA in 10 mM Tris-HCl (pH 7.0) and 140 mM KCl.
Isothermal Titration Calorimetry (ITC) Study
5 µM of RNA (CESC-KISS, CESC-NOKISS and CESCD-
NOKISS) are titrated to 100 µM of NC protein.
RNA Dimerization by Gel Shift Mobility Assay
native, CESC-KISS, CESC-NOKISS, and CESCD-NOKISS
RNA dimerization was performed in 10 mM Tris-HCl
(pH 7.0), 140 mM KCl and 1 mM MgCl2. RNA was
incubated for a series of time intervals at 37oC.
CESNATIVE
CESC-KISS
CESC-NOKISS
Dimer
Monomer
0 0.25 0.5 1.0 1.5 2.0 3.0 6.0 24 Hour
Dimer
Monomer
Dimer
Monomer