1. Neuropsychiatric disorders are pervasive in contemporary society, and their
underlying causes are unclear—actual physiological explanations for many of
these diseases are lacking. Mood and anxiety disorders account for a large
proportion of such diseases and are associated with dysfunctions of the brain’s
corticolimbic system, which is linked to emotional processing and regulation.
The corticolimbic circuit includes the basolateral amygdala (BLA),
hippocampus (HPC), prelimbic cortex (PrL), and infralimbic cortex (IL).
When disrupted, the circuit demonstrates a hypersensitivity to threat that is
evident in mood and anxiety disorders. Electrophysiological research in
rodents has revealed synchronous oscillations in the BLA, HPC, and PFC
during fear recall (Seidenbecher 2003; Pape 2005; Narayanan 2007), though
very few studies have observed neural activity during the initial acquisition of
a fear memory. The current study focuses on the oscillatory interactions
between multiple corticolimbic structures and seeks to understand their
interdependent roles in the fear learning process.
In this study, rats have undergone Pavlovian fear conditioning after being
implanted with 32-channel microwire electrode arrays targeting the BLA,
ventral HPC, PrL and IL. In vivo electrophysiological recording techniques
captured neuronal spikes and local field potentials within these structures
throughout habituation, acquisition, recall, and extinction of the fear memory.
Here, we present the result of behavioral analysis from differential fear
conditioning. In future, LFPs and spikes will be carefully analyzed for
correlations with behavioral outputs including time spent freezing and
individual behavior suppression ratio (BSR).
1. Animals and Pre-training:
Seven individually housed, male, Harlan Sprague-Dawley rats at 8-10 weeks of age were
maintained on a restricted diet at no less than 85% of their normal growth curve. Animals were
trained to nosepoke for a food pellet at random intervals with a mean of 60 seconds. The goal of
this training was to achieve a baseline activity level against which to compare freeze as an
indication of fear.
2. Surgery:
Under isoflurane anesthesia, the rats were implanted with custom 32-channel arrays of 50-
micron tungsten microwires, targeting the basolateral amygdala (BLA), infralimbic cortex (IL),
prelimbic cortex (PrL), and ventral hippocampus (vHPC). Rats were allowed one week for post-
surgical recovery.
Methods
Introduction
Conclusions
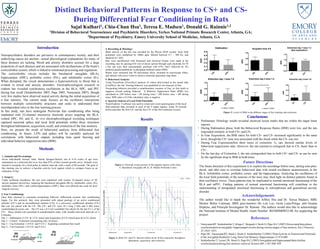
• Preliminary Histology results revealed electrical lesion marks that are within the target brain
regions.
• During Habituation phase, both the Behavioral Response Ratios (BSR) were low, and the rats
responded similarly to both CS+ and CS-
• In Fear Acquisition, the BSR ratios for both CS+ and CS- increased significantly to the same
level, though the CS+ tone was associated with the shock while the CS- was not.
• During Fear Expression(first three tones of extinction 1), rats showed similar levels of
behavioral suppression ratio. However, the rats learned to extinguish fear to CS- faster than to
CS+.
• On the last day of Extinction 2, the rats extinguished fear to both CS+ and CS- as can be seen
by the significant drop in BSR in both tones.
Results
• NarayananRT, Seidenbecher T, Kluge C, Bergado J, Stork O, Pape H-C (2007) Dissociated theta phase
synchronizationin amygdalo- hippocampalcircuits during various stages of fear memory. Eur J Neurosci
25:1823–1831.
• Pape HC, NarayananRT, Smid J, Stork O, Seidenbecher T(2005) ThetaActivity in Neuronsand Networks
of the Amygdala Relatedto Long-Term Fear Memory.Hippocampus 15:874-880
• SeidenbecherT, LaxmiTR, Stork O, Pape H-C (2003)Amygdalar and hippocampaltheta rhythm
synchronizationduring fear memory retrieval.Science (80- ) 301:846–850
3. Fear Conditioning:
Once Rats returned to consistent nosepoking behavior, differential acoustic fear conditioning
began. For this protocol, they were presented with repeat pairings of an active conditioned
stimulus (CS+) and an unconditioned stimulus (US), or a non-active conditioned stimulus (CS-)
that was not paired with the US. The CS+ and CS- were 30 s long 2 kHz and 6 kHz tones,
counterbalanced across rats. The US was a 0.5 mA scrambled foot shock for the last 0.5 s of the
CS+. These stimuli were presented in pseudorandom order, with variable inter-trial intervals of 1-
6 minutes.
Day 1 – Habituation (3 CS+ & 3 CS- tones) and Acquisition (6 CS+/shock pairs & 6 CS- alone)
Day 2 – Context Extinction (nosepoking only, no tones)
Day 3 – Cue Extinction 1 (9 CS+ and 9 CS-) – beginning considered fear recall
Day 4 – Cue Extinction 2 (9 CS+ and 9 CS-)
4. Recording & Histology:
Brain activity of the rats was recorded by the Plexon MAP system: local field
potentials were multiplied by 1000x gain, filtered between 0.7 – 300 Hz, and
digitized at 1 kHz.
Rats were anesthetized with Somnasol and electrical lesions were made at the
recording sites by passing 0.01 mA of direct current through each electrode for 45
s.The rats were then transcardially perfused with 0.9% NaCl followed by 4%
buffered paraformaldehyde in phosphate buffered saline (PBS).
Brains were sectioned into 50 micrometer slices, mounted on microscope slides,
and stained with cresyl violet to observe electrode placement map them.
5. Behavioral Analysis:
Using FreezeScan (CleverSys) analysis of videos time-locked to the neural data
(CinePlex), the rats’ freezing behavior was quantified as one measure of fear.
Nosepoking behavior provided a complimentary measure of fear, as fear tends to
suppress reward seeking behavior. A Behavior Suppression Ratio (BSR) was
calculated as (NP before tone - NP during tone) / (BP before tone + BP during
tone). BSR value is 0 if the calculated value is negative.
Future Directions
The future direction of this experiment is to explore the recordings before tone, during tone-plus-
shock, and after tone to correlate behavioral data with neural oscillation measurements in the
BLA, Infralimbic cortex, prelimbic cortex, and the hippocampus. Analyzing the oscillations of
the local field potentials of the neurons of the mice may shed light on distinct patterns found in
their oscillatory waves. These patterns may be implicated to normal emotional functioning of the
BLA and mPFC. Finding patterns of normal emotional functioning will contribute to the
understanding of deregulated emotional functioning in schizophrenia and generalized anxiety
disorder.
6. Spectral Analysis of Local Field Potentials:
NeuroExplorer 5 software was used to create peri-event spectrograms of the local
field potential data recorded in each of the 4 target regions, using 30 seconds
before and after the first CS+ and the first CS- of the first extinction session.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9
Habituation Acquisition Extinction1 Extinction2
Behavioralsurpressionratio(BSR)
CS- CS+
Distinct Behavioral Patterns in Response to CS+ and CS-
During Differential Fear Conditioning in Rats
Sajal Kulhari1, Chia-Chun Hsu1, Teresa E. Madsen1, Donald G. Rainnie1,2
1Division of Behavioral Neuroscience and Psychiatric Disorders, Yerkes National Primate Research Center, Atlanta, GA;
2Department of Psychiatry, Emory University School of Medicine, Atlanta, GA
C
S-
C
S+
0.0
0.1
0.2
0.3
0.4
0.5
Habituation
BSR
C
S-
C
S+
0.0
0.5
1.0
1.5
Acquition tone 4-6
BSR
C
S-
C
S+
0.0
0.5
1.0
1.5
Extinction day 1 tone 1-3
(fear expression)
BSR
C
S-
C
S+
0.0
0.5
1.0
1.5
Extinction day 1 tone 7-9
BSR
p value= 0.0640
C
S-
C
S+
0.0
0.5
1.0
1.5
Extinction day 2 tone 1-3
BSR
p value= 0.0353
*
C
S-
C
S+
0.0
0.2
0.4
0.6
Extinction day 2 tone 7-9
BSR
Acknowledgements
The author would like to thank the wonderful Jeffrey Hsu and Dr. Teresa Madsen, SIRE
Mentor Brilee Coleman, SIRE peer-mentors Ho Lim Lee, Greta Luna-Priego, and Oceana
Hopkins, and Principle Investigator Dr. Rainnie. The authors would also like to acknowledge
the National Institute of Mental Health, Grant Number: R01MH069852-08, for supporting the
project.
References
Figure 2: Both CS+ and CS- showed similar levels of fear expression throughout
habituation, acquisition, and extinction.
Figure 3: Levels of BSR in the different stages of fear learning and extinction.
No shock
shock
Figure 1: Electrode lesions present in the targeted regions of the brain.
A. Basolateral Amygdala (BLA). B. Medial Prefrontal Cortex.
A. B.