Cerebellum
• Integrates massivesensory and
other inputs from many regions
of the brain and spinal cord
• smoothly coordinates ongoing
movements and to participate in
motor planning
• has no direct connections to
lower motor neurons, but instead
exerts its influence through
connections to motor systems of
the cortex and brainstem
3.
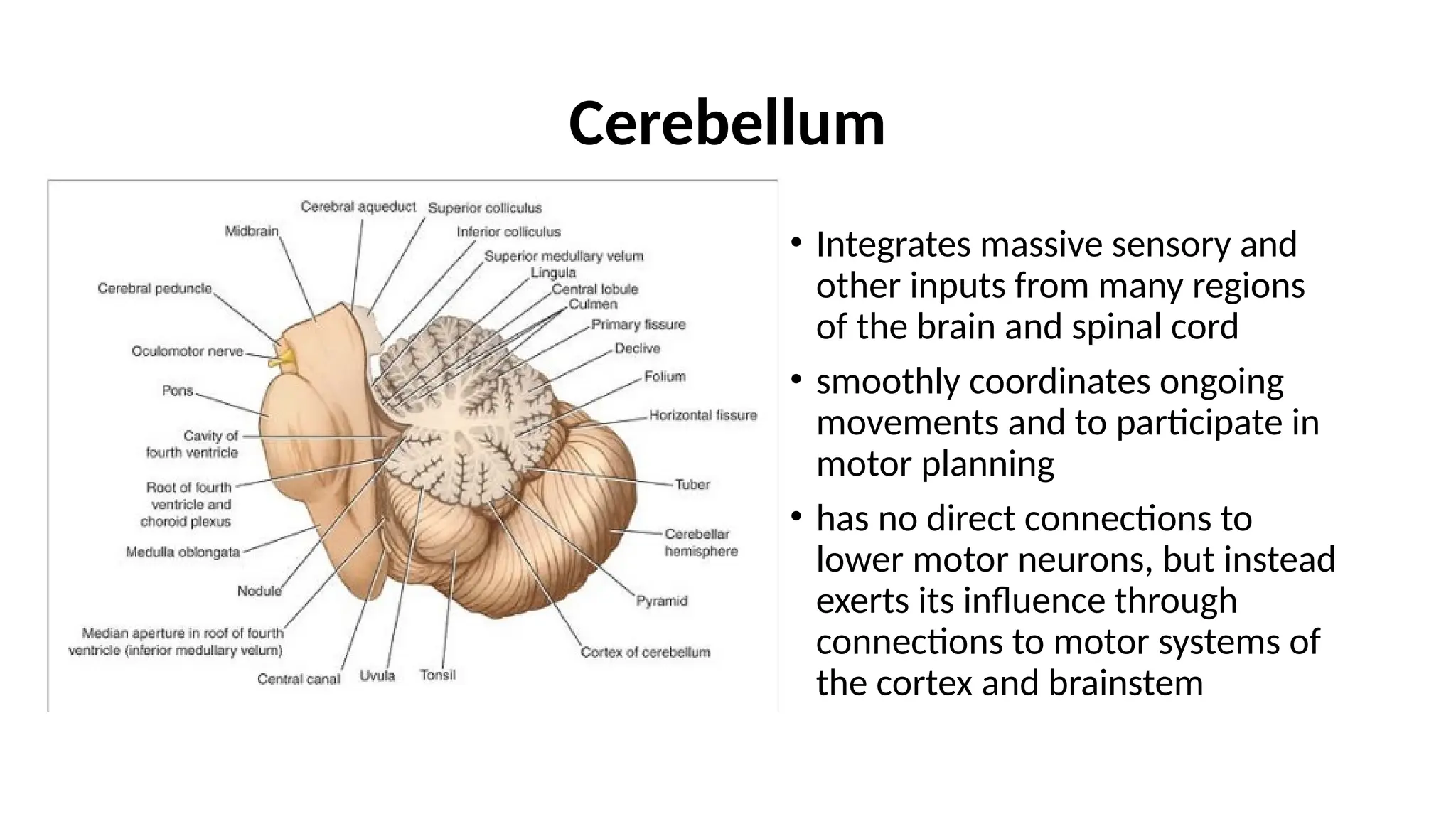
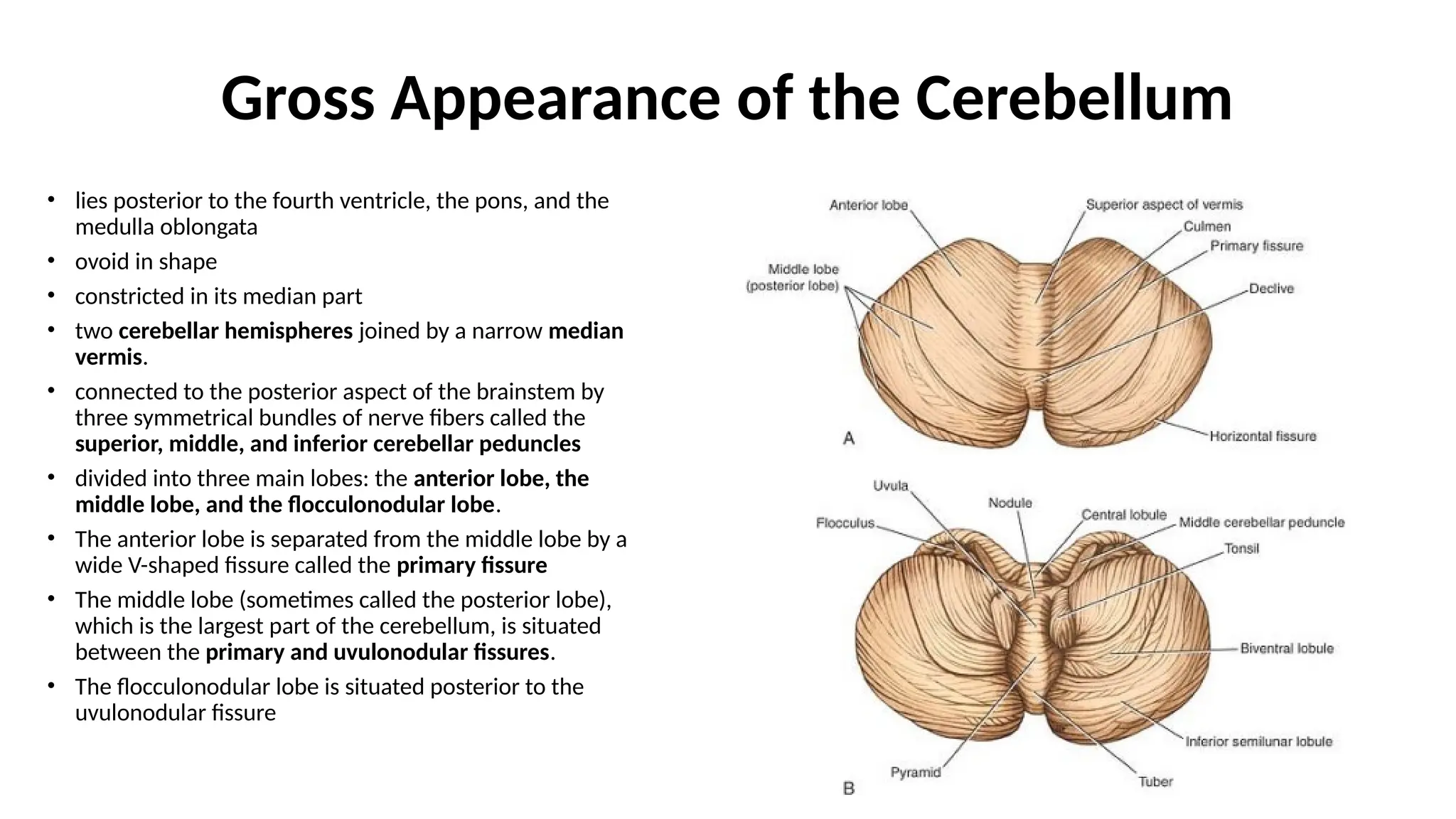
Gross Appearance ofthe Cerebellum
• lies posterior to the fourth ventricle, the pons, and the
medulla oblongata
• ovoid in shape
• constricted in its median part
• two cerebellar hemispheres joined by a narrow median
vermis.
• connected to the posterior aspect of the brainstem by
three symmetrical bundles of nerve fibers called the
superior, middle, and inferior cerebellar peduncles
• divided into three main lobes: the anterior lobe, the
middle lobe, and the flocculonodular lobe.
• The anterior lobe is separated from the middle lobe by a
wide V-shaped fissure called the primary fissure
• The middle lobe (sometimes called the posterior lobe),
which is the largest part of the cerebellum, is situated
between the primary and uvulonodular fissures.
• The flocculonodular lobe is situated posterior to the
uvulonodular fissure
4.
Gross Appearance ofthe Cerebellum
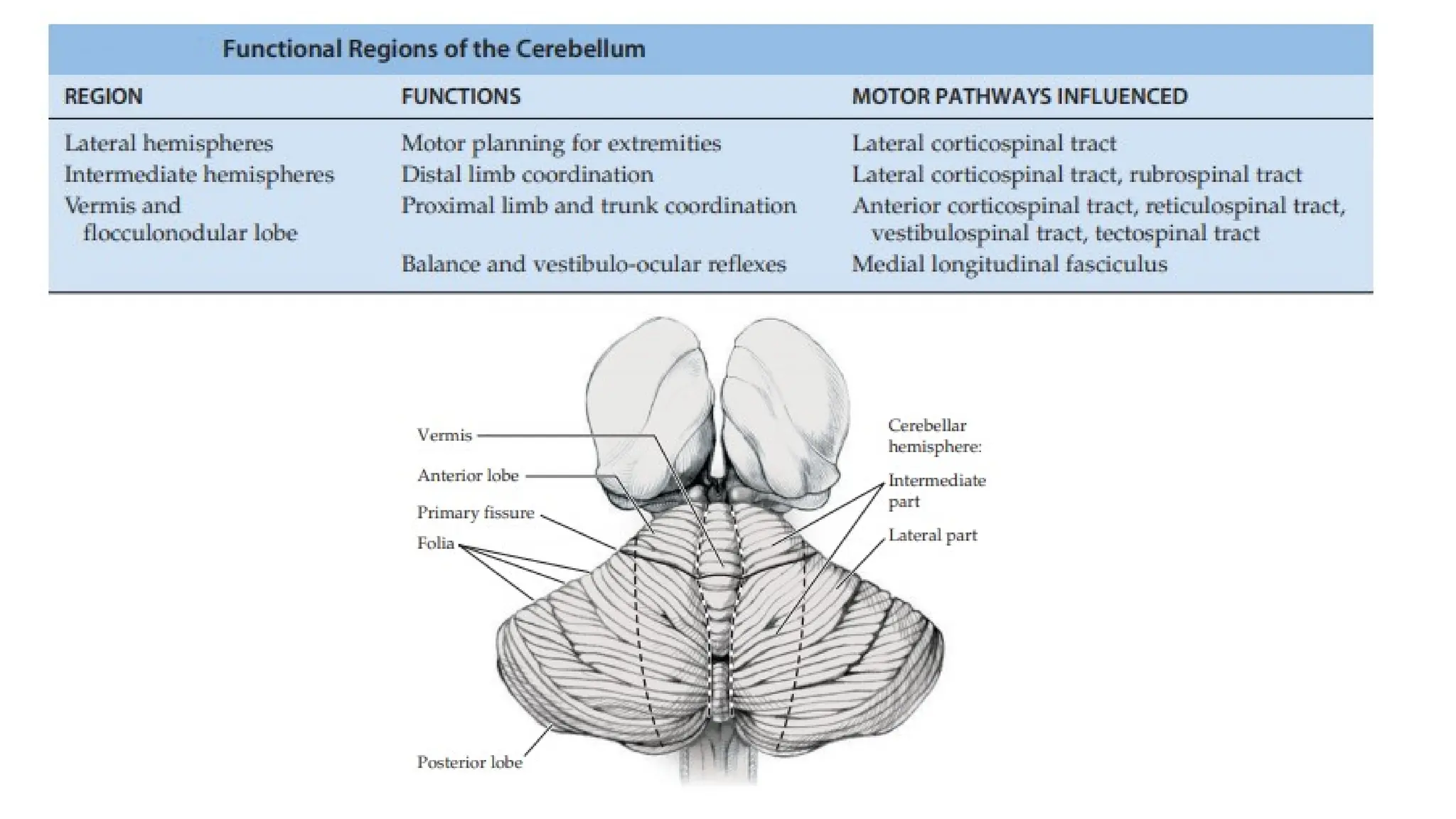
The cerebellum is organized into different regions with specialized functions:
• The inferior vermis and flocculonodular lobes regulate balance and eye
movements through interactions with the vestibular circuitry. These regions,
together with other parts of the vermis, are involved in control of the
medial motor systems (proximal trunk muscles).
• Intermediate cerebellar regions control the lateral motor systems (distal
appendicular muscles).
• Finally, large regions of the most lateral cerebellar hemispheres are
important in motor planning - with the planning of sequential movements
of the entire body and is involved with the conscious assessment of
movement errors.
6.
Ataxia
• Cerebellar lesionstypically result in a characteristic type of irregular
uncoordinated movement called ataxia.
1. Ataxia is ipsilateral to the side of a cerebellar lesion.
2. Midline lesions of the cerebellar vermis or flocculonodular lobes
mainly cause unsteady gait (truncal ataxia) and eye movement
abnormalities, which are often accompanied by intense vertigo,
nausea, and vomiting.
3. Lesions lateral to the cerebellar vermis mainly cause ataxia of the
limbs (appendicular ataxia).
7.
Structure of theCerebellum
• an outer covering of gray matter called the cortex
• inner white matter
• intracerebellar nuclei - masses of gray matter - embedded in the
white matter of each hemisphere
8.
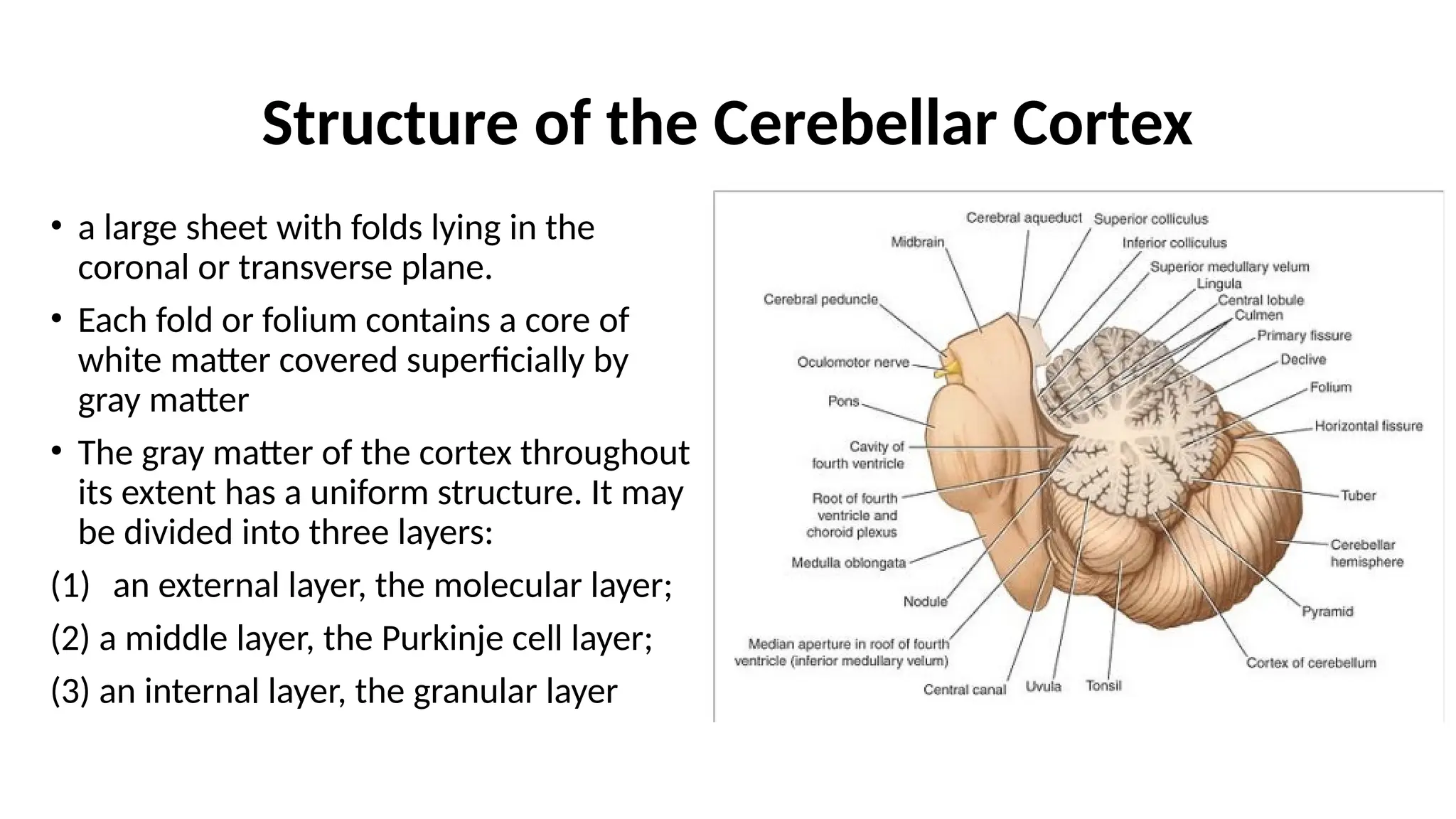
Structure of theCerebellar Cortex
• a large sheet with folds lying in the
coronal or transverse plane.
• Each fold or folium contains a core of
white matter covered superficially by
gray matter
• The gray matter of the cortex throughout
its extent has a uniform structure. It may
be divided into three layers:
(1) an external layer, the molecular layer;
(2) a middle layer, the Purkinje cell layer;
(3) an internal layer, the granular layer
9.
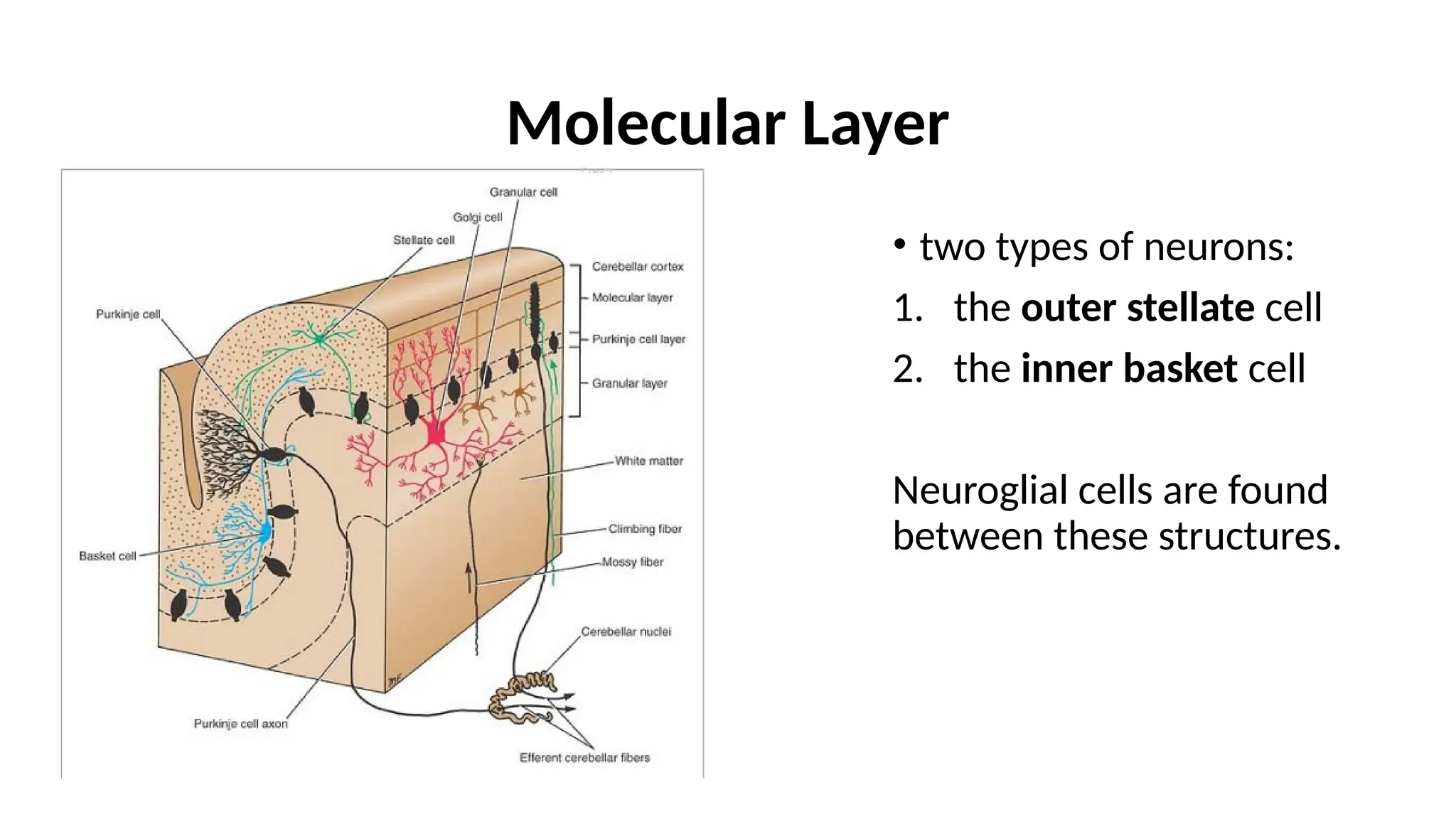
Molecular Layer
• twotypes of neurons:
1. the outer stellate cell
2. the inner basket cell
Neuroglial cells are found
between these structures.
10.
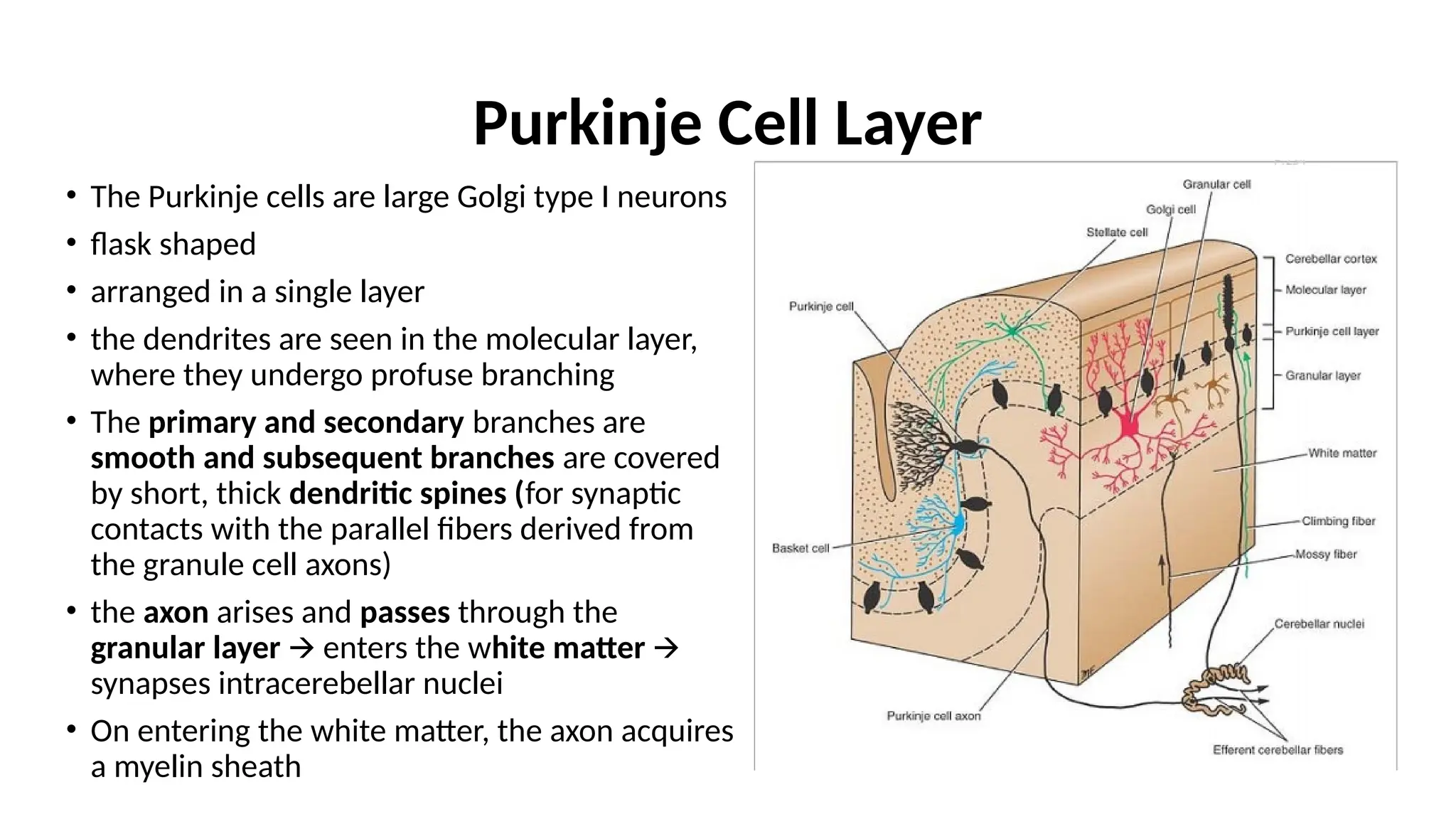
Purkinje Cell Layer
•The Purkinje cells are large Golgi type I neurons
• flask shaped
• arranged in a single layer
• the dendrites are seen in the molecular layer,
where they undergo profuse branching
• The primary and secondary branches are
smooth and subsequent branches are covered
by short, thick dendritic spines (for synaptic
contacts with the parallel fibers derived from
the granule cell axons)
• the axon arises and passes through the
granular layer enters the w
🡪 hite matter 🡪
synapses intracerebellar nuclei
• On entering the white matter, the axon acquires
a myelin sheath
11.
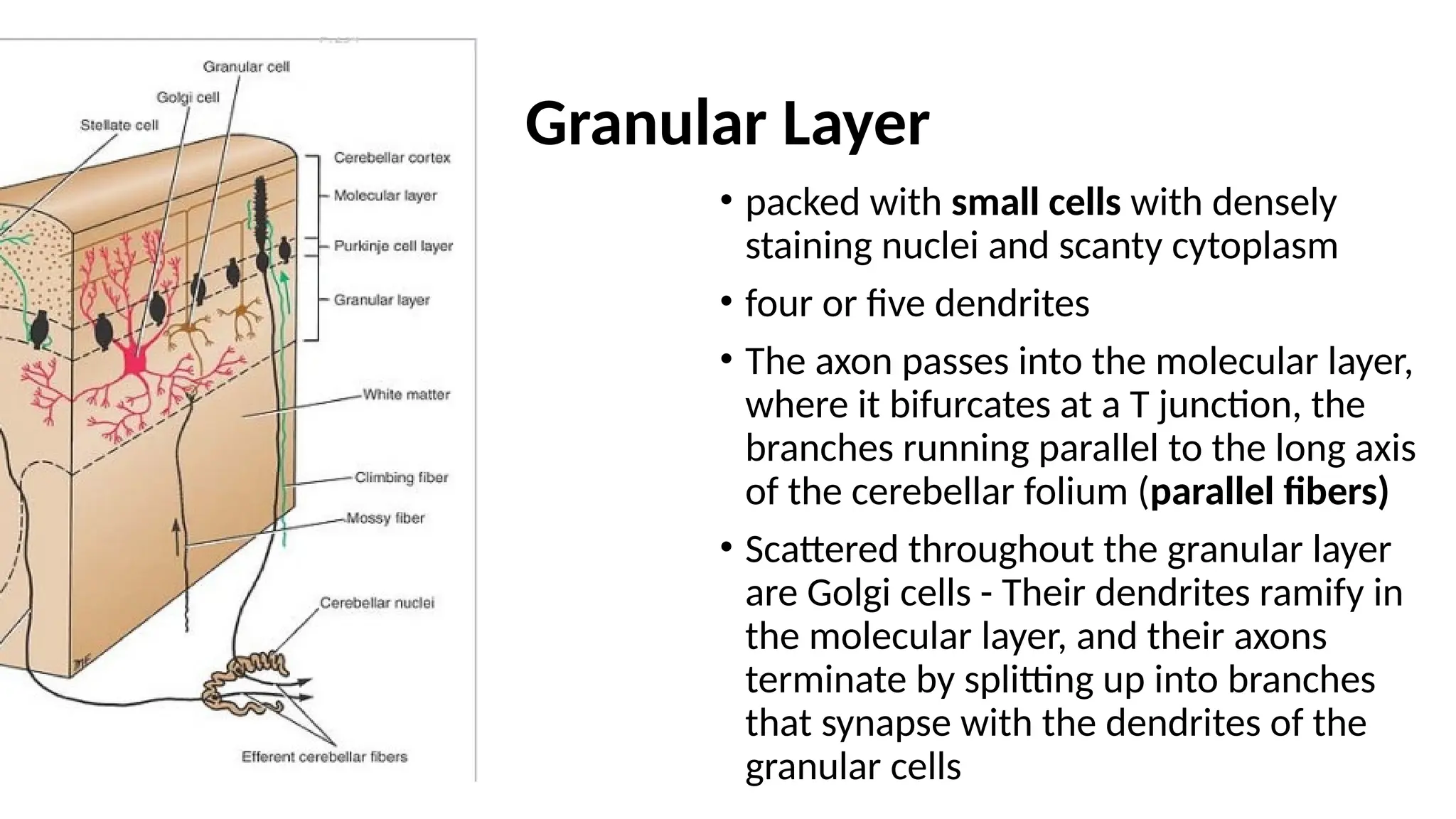
Granular Layer
• packedwith small cells with densely
staining nuclei and scanty cytoplasm
• four or five dendrites
• The axon passes into the molecular layer,
where it bifurcates at a T junction, the
branches running parallel to the long axis
of the cerebellar folium (parallel fibers)
• Scattered throughout the granular layer
are Golgi cells - Their dendrites ramify in
the molecular layer, and their axons
terminate by splitting up into branches
that synapse with the dendrites of the
granular cells
12.
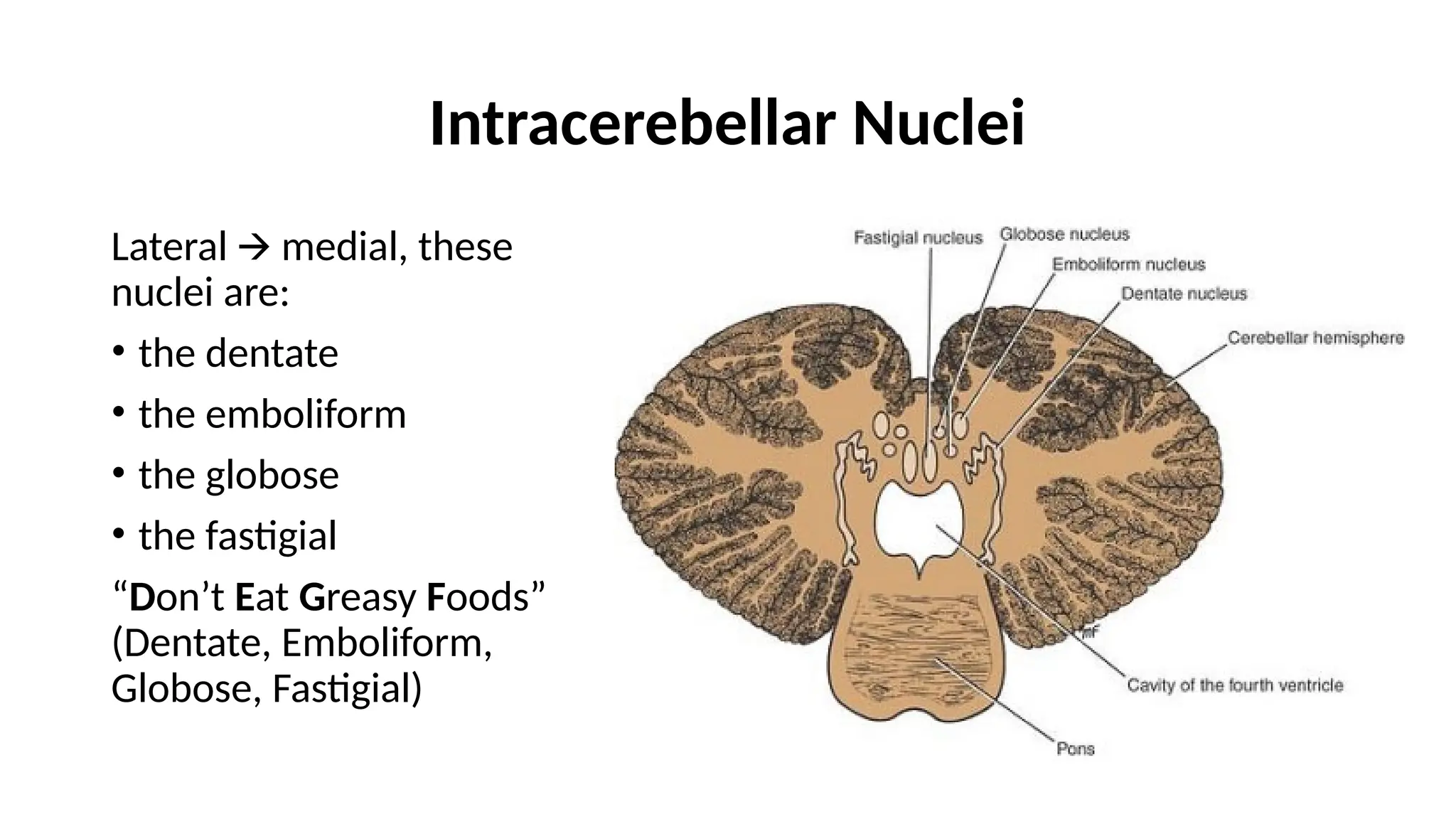
Intracerebellar Nuclei
Lateral medial,these
🡪
nuclei are:
• the dentate
• the emboliform
• the globose
• the fastigial
“Don’t Eat Greasy Foods”
(Dentate, Emboliform,
Globose, Fastigial)
13.
Intracerebellar Nuclei
• Thedentate nucleus is the largest of the cerebellar nuclei. It has the shape of a crumpled bag with
the opening facing medially. The interior of the bag is filled with white matter made up of efferent
fibers that leave the nucleus through the opening to form a large part of the superior cerebellar
peduncle.
• The emboliform nucleus is ovoid and is situated medial to the dentate nucleus.
• The globose nucleus consists of one or more rounded cell groups that lie medial to the emboliform
nucleus.
• The fastigial nucleus lies near the midline in the vermis and close to the roof of the fourth ventricle; it
is larger than the globose nucleus.
NOTE!!! The intracerebellar nuclei are composed of large, multipolar neurons with simple branching
dendrites.
Fibers from the dentate, emboliform, and globose nuclei leave the cerebellum through the superior
cerebellar peduncle.
Fibers from the fastigial nucleus leave through the inferior cerebellar peduncle.
14.
Intracerebellar Nuclei
• Thedentate nuclei projections from the
🡨 lateral cerebellar
hemispheres.
• The emboliform and globose nuclei are together called the
interposed nuclei🡨 intermediate part of the cerebellar hemispheres
• NOTE! experimental recordings have shown the dentate nucleus to be
active just before voluntary movements, while the interposed nuclei
are active during and in relation to the movement.
• The fastigial nuclei the vermis and a small input from the
🡨
flocculonodular lobe.
15.
White Matter
• madeup of three groups of fibers:
(1) Intrinsic
(2) Afferent
(3) efferent.
The intrinsic fibers do not leave the cerebellum but connect different regions of the organ.
Some interconnect folia of the cerebellar cortex and vermis on the same side; others connect
the two cerebellar hemispheres together.
The afferent fibers form the greater part of the white matter and proceed to the cerebellar
cortex. They enter the cerebellum mainly through the inferior and middle cerebellar peduncles.
The efferent fibers constitute the output of the cerebellum and commence as the axons of the
Purkinje cells of the cerebellar cortex. The great majority of the Purkinje cell axons pass to and
synapse with the neurons of the cerebellar nuclei (fastigial, globose, emboliform, and dentate).
16.
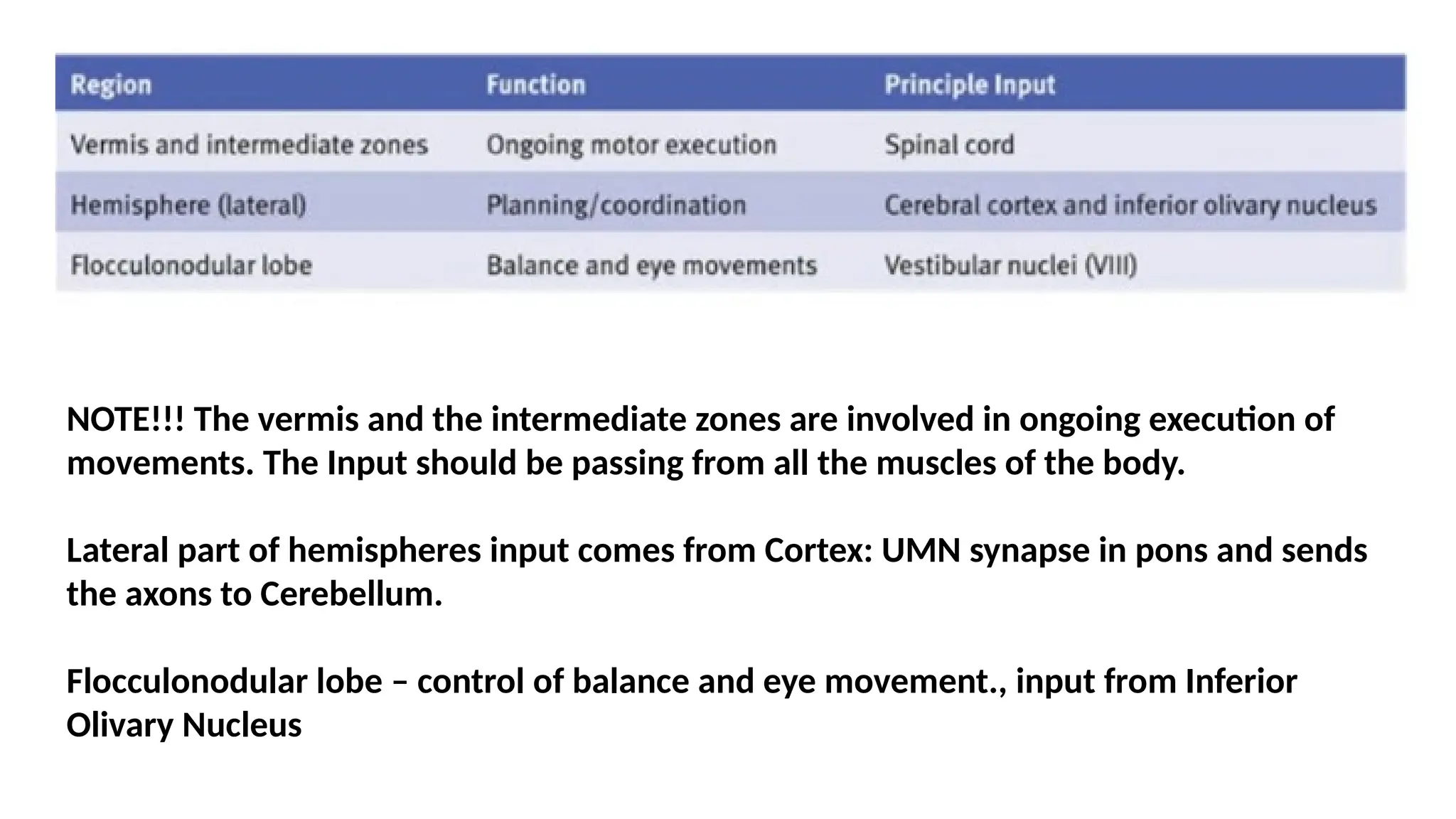
NOTE!!! The vermisand the intermediate zones are involved in ongoing execution of
movements. The Input should be passing from all the muscles of the body.
Lateral part of hemispheres input comes from Cortex: UMN synapse in pons and sends
the axons to Cerebellum.
Flocculonodular lobe – control of balance and eye movement., input from Inferior
Olivary Nucleus
Cerebellar Cortical Mechanisms
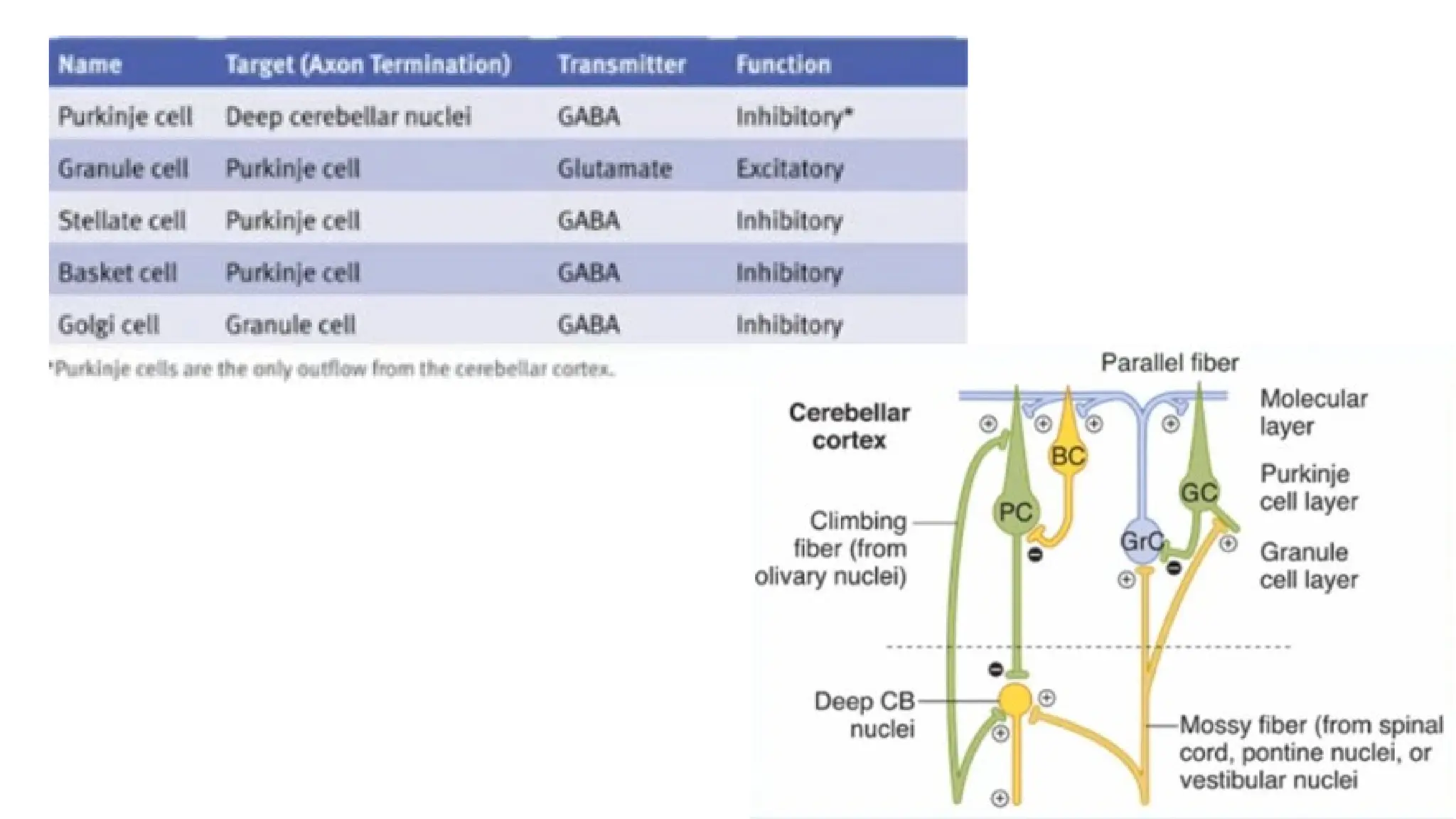
NOTE!!!The climbing and the mossy fibers constitute the two main lines of input to the cortex and are
excitatory to the Purkinje cells
Entire output of the Purkinje cells are INHIBITORY THE MAIN FUNCTION TO SMOOTHEN THE
🡪
MOVEMENT
Inferior Olivary Nuclei Climbing Fibers (ERRORs of Movement) Excitation of Purkinje cells Developing
🡪 🡪
the motor learning
(A single Purkinje neuron makes synaptic contact with only one climbing fiber. However, one climbing fiber
makes contact with 1 to 10 Purkinje neurons. )
Spinal Cord, Pontine nuclei, Vestibular nuclei Mossy Fibers (General condition of the body/ movement)
🡪
(A single mossy fiber may stimulate thousands of Purkinje cells through the granule cells)
20.
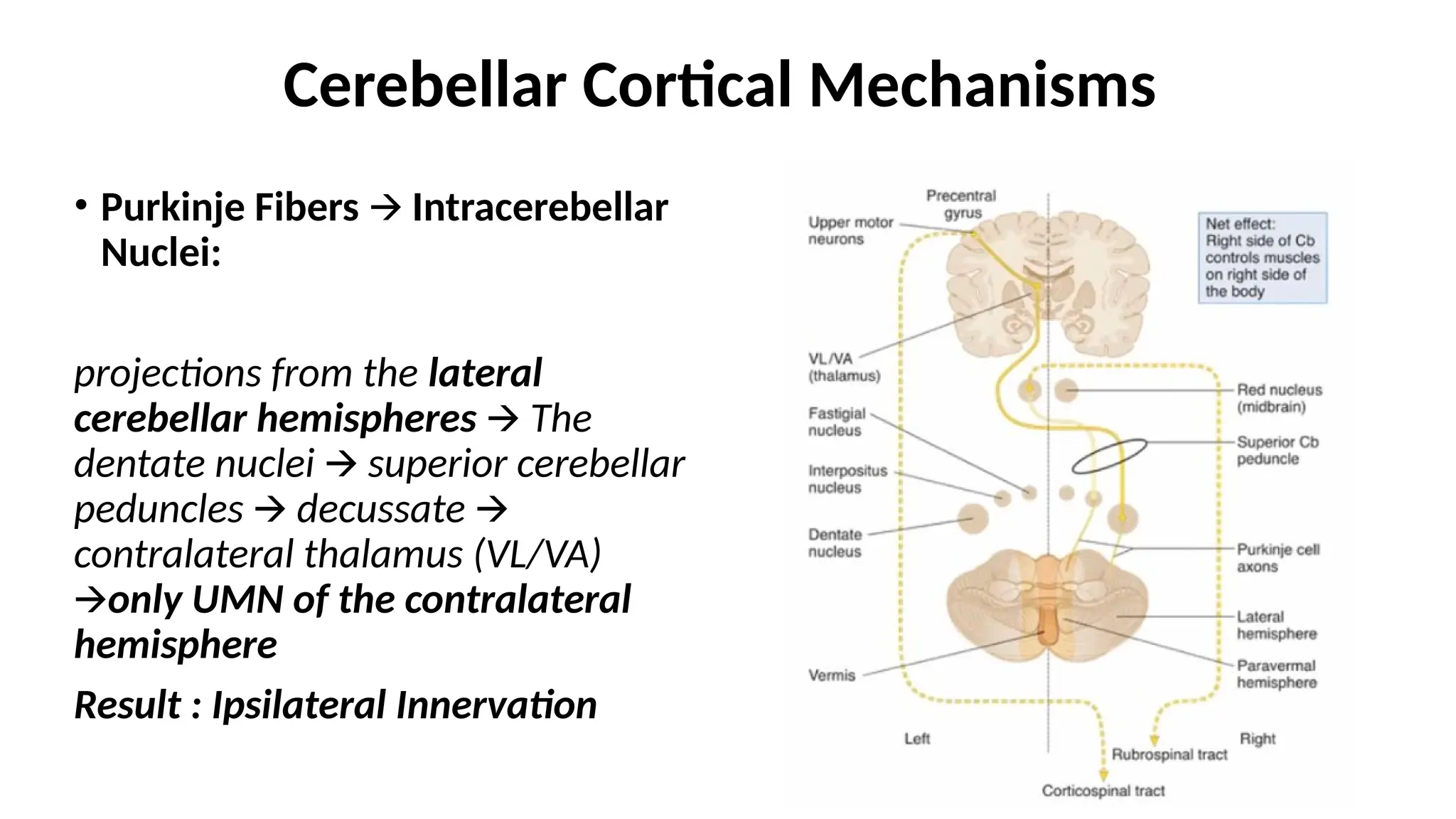
Cerebellar Cortical Mechanisms
•Purkinje Fibers Intracerebellar
🡪
Nuclei:
projections from the lateral
cerebellar hemispheres 🡪 The
dentate nuclei superior cerebellar
🡪
peduncles decussate
🡪 🡪
contralateral thalamus (VL/VA)
🡪only UMN of the contralateral
hemisphere
Result : Ipsilateral Innervation
21.
Input Pathways tothe Cerebellum
• Corticopontocerebellar pathway, which originates in the cerebral
motor and premotor cortices and also in the cerebral somatosensory
cortex. It passes by way of the pontile nuclei and pontocerebellar
tracts mainly to the lateral divisions of the cerebellar hemispheres on
the opposite side of the brain from the cerebral areas.
• Olivocerebellar tract, which passes from the inferior olive to all parts
of the cerebellum and is excited in the olive by fibers from the
cerebral motor cortex, basal ganglia, widespread areas of the reticular
formation, and spinal cord;
22.
Input Pathways tothe Cerebellum
• Vestibulocerebellar fibers, some of which originate in the vestibular
apparatus itself and others from the brain stem vestibular nuclei—
almost all of these terminate in the flocculonodular lobe and fastigial
nucleus of the cerebellum
• Reticulocerebellar fibers, which originate in different portions of the
brain stem reticular formation and terminate in the midline cerebellar
areas (mainly in the vermis).
23.
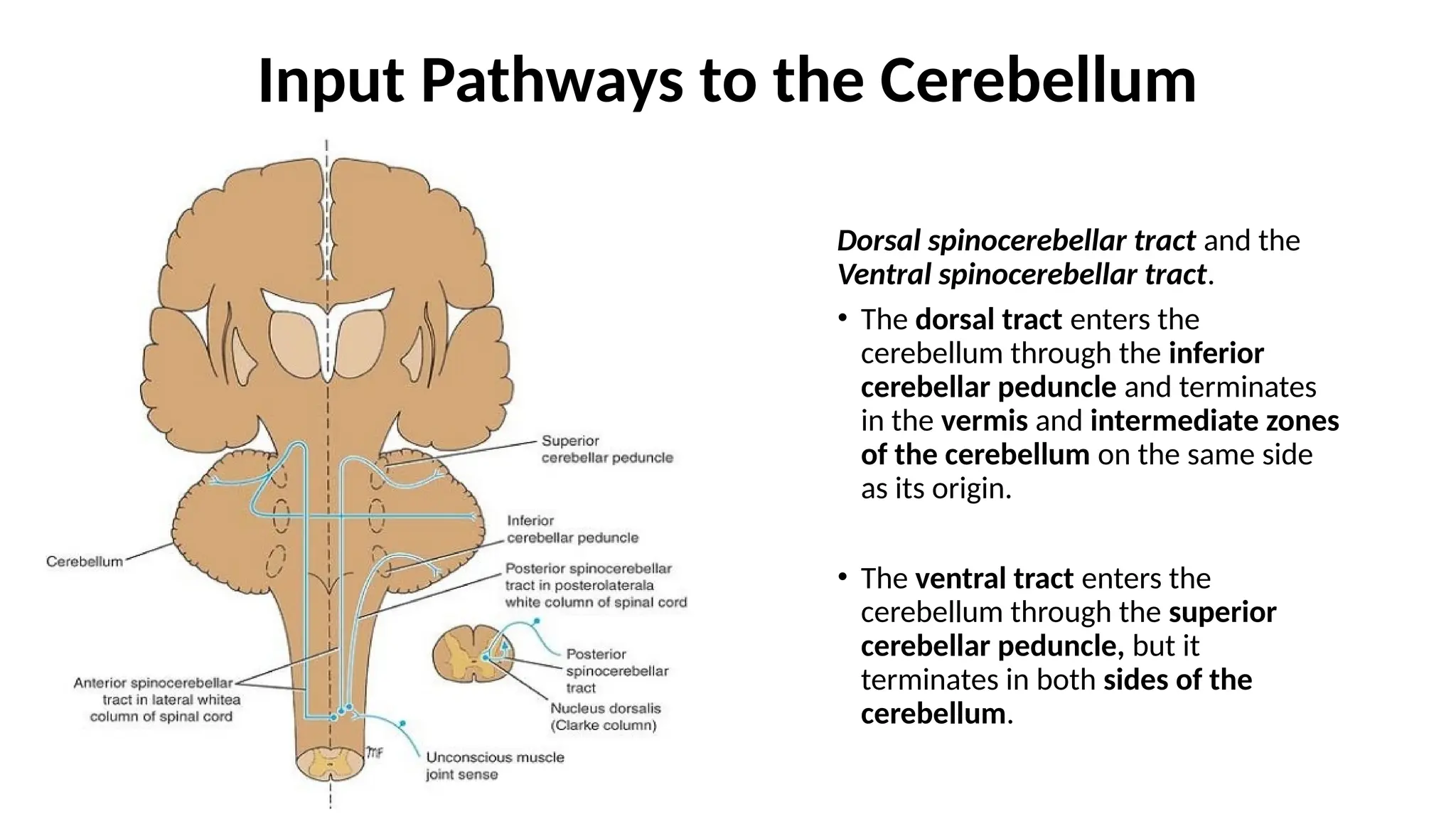
Input Pathways tothe Cerebellum
Dorsal spinocerebellar tract and the
Ventral spinocerebellar tract.
• The dorsal tract enters the
cerebellum through the inferior
cerebellar peduncle and terminates
in the vermis and intermediate zones
of the cerebellum on the same side
as its origin.
• The ventral tract enters the
cerebellum through the superior
cerebellar peduncle, but it
terminates in both sides of the
cerebellum.
24.
Input Pathways tothe Cerebellum
• The signals transmitted in the dorsal spinocerebellar tracts come
mainly from the muscle spindles and to a lesser extent from other
somatic receptors throughout the body, such as Golgi tendon organs,
large tactile receptors of the skin, and joint receptors.
• All these signals apprise the cerebellum of the momentary status of
(1) muscle contraction, (2) degree of tension on the muscle tendons,
(3) positions and rates of movement of the parts of the body, and (4)
forces acting on the surfaces of the body.
25.
Input Pathways tothe Cerebellum
• The ventral spinocerebellar tracts receive much less information from
the peripheral receptors.
• Instead, they are excited mainly by motor signals arriving in the
anterior horns of the spinal cord from
(1) the brain through the corticospinal and rubrospinal tracts and
(2) the internal motor pattern generators in the cord itself.
Thus, this ventral fiber pathway tells the cerebellum which motor
signals have arrived at the anterior horns; this feedback is called the
efference copy of the anterior horn motor drive
26.
• The spinocerebellarpathways can transmit impulses at velocities up
to 120 m/sec, which is the most rapid conduction in any pathway in
the central nervous system.
• This extremely rapid conduction is important for instantaneous
apprisal of the cerebellum of changes in peripheral muscle actions
27.
Output Signals fromthe Cerebellum
A pathway that originates in the midline structures of the cerebellum
(the vermis) and then passes through the fastigial nuclei into the
medullary and pontile regions of the brain stem.
This circuit functions in close association with the equilibrium
apparatus and brain stem vestibular nuclei to control equilibrium, as
well as in association with the reticular formation of the brain stem to
control the postural attitudes of the body.
28.
Output Signals fromthe Cerebellum
A pathway that originates in (1) the intermediate zone of the cerebellar
hemisphere and then passes through (2) the interposed nucleus to (3)
the ventrolateral and ventroanterior nuclei of the thalamus and then to
(4) the cerebral cortex, to (5) several midline structures of the thalamus
and then to (6) the basal ganglia and (7) the red nucleus and reticular
formation of the upper portion of the brain stem. This complex circuit
helps to coordinate mainly the reciprocal contractions of agonist and
antagonist muscles in the peripheral portions of the limbs, especially in
the hands, fingers, and thumbs.