Recommended

More Related Content
Similar to Bee Poster_
Similar to Bee Poster_ (20)
Bee Poster_
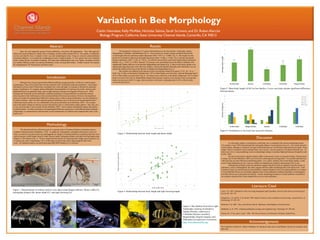
- 1. Abstract Methodology Results Discussion Literature Cited Acknowledgements Introduction Bees, the most important group of insect pollinators, coevolved with angiosperms. They feed upon the pollen and nectar produced by plants, and in exchange, transfer pollen among flowers. The quality of pollination services provided by bees is related to their size and morphological traits. Using an extensive plant-pollinator interaction database, we recorded the morphological traits of approximately 111 bee species that were collected while visiting flowers in northern California. We found that morphological traits were highly correlated, but that bee families differed in their size and the distribution of hair covering their bodies. Further research will explore the morphological “fit” between bee and flower traits. We characterized bees collected as part of a regional study of 10 plant-pollinator communities along a northern California transect (Moldenke, 1976). To update the nomenclature, and gather information on bee size traits, we used eol.org, discoverlife.org, and bugguide.net. Images of female pinned bees containing scale bars were used to measure overall length, thoracic width, thoracic depth, intertegular distance and right forewing length using ImageJ (v1.48) software (Fig. 1). We also categorized hair density by examining these images and scoring bees on a 5 point scale, with 1 representing ≤ 20 % of the body covered in hair, and 5 representing 80-100 % hair cover. All statistical analysis was performed using IBM SPSS Statistics (v21) software. We attempted to characterize 111 species distributed across five bee families: Andrenidae, Apidae, Megachilidae, Colletidae, and Halictidae (Fig. 2). Across species we found a strong correlation between the overall length of the bee and thorax depth (Spearman’s Rho = 0.817, p < 0.0001, Fig. 3). Bee length was also positively correlated to right forewing length (Spearman’s Rho = 0.466, p < 0.001, Fig. 4) and the intertegular distance (Spearman’s Rho = 0.501, p < 0.015). An ANOVA showed that overall body length differed among bee families (F4,69 = 8.857, p ≤ 0.001). Dunnett’s T3 post-hoc tests showed that bees in the family Colletidae were significantly smaller than bees in the families Megachilidae and Apidae (Fig. 5). Bees in the family Apidae were significantly larger than bees in the other four families, whereas the families Halictidae and Andrenidae were not statistically different from bees in the families Colletidae and Megachilidae (Fig. 5). Bee families also differed in the amount of hair covering their bodies (Chi-square 72.471, df = 16, p < 0.001; Fig. 6). Bees in the family Colletidae had ≤ 20 % of their bodies covered in hair, while the Halictidae had 0- 40 % of their bodies covered in hair (Fig. 6). In contrast, most of the bees in the family Apidae had ≥ 60 % of their bodies covered in hair (Fig. 6). Megachilidae displayed the greatest variation in the amount of hair covering their bodies, but most were 40-61 % range, while most bees in the family Andrenidae had 21-40 % of their bodies covered in hair (Fig. 6). As with earlier studies, we found that overall body size is correlated with various morphological traits. For example, Cane (1987) demonstrated that intertegular distance is proportional to bee size. Our results showed a similar pattern, although we detected a stronger relationship between overall body length and thoracic depth. Wing length, which is commonly used to estimate body size in insects, was also positively correlated with body size. As body size evolves, wing morphology must correspondingly change to maintain flight performance (Kingsolver and Koehl, 1994). We also found evidence that bee families differ in the amount of hair covering their bodies. Plumose hairs, a unique trait of bees (Michener, 2007), are involved in collecting and carrying pollen. It is possible then that bees with more hair are more efficient at transferring pollen. If so, plants visited by bees in the family Apidae, would receive better pollination services; however further investigations are needed to demonstrate if this is true. Although the pollinator syndrome hypothesis suggests that floral traits are adapted to specific groups of pollinators, such as hummingbirds, this study suggests that bees differ morphologically at the family level. Such variation in body size and hairiness has the potential to impact pollination, and in turn plant-pollinator coevolution. It is possible that flowers are not broadly adapted to bees as the pollination syndrome hypothesis would suggests, but rather flowers are to particular bee families. Further morphological analyses of plant-pollinator communities are needed to determine how finely adapted flowers are to their pollinators. Cane, J. H. 1987. Estimation of bee size using intertegular span (Apoidea). Journal of the Kansas Entomological Society 60: 145–147. Kingsolver, J. G. and M. A. R. Koehl. 1994. Selective factors in the evolution of insects wings. Annual Review of Entomology 39: 425–451. Michener, C.D. 2007. Bees of the World, 2nd ed. Baltimore, John Hopkins University Press. Moldenke, A. R. 1976. California pollination ecology and vegetation type. Phytologia 34: 305-361. Proctor, M., P. Yeo, and A. Lack. 1996. The Natural History of Pollination. Portland, Timber Press. We would like to thank Dr. Andrew Moldenke for sharing his data with us and Bethany Wynne for assistance with data entry. Biologists have long recognized that different pollinator groups generally visit flowers with divergent morphologies. These observations led to the development of the “pollination syndrome hypothesis,” which states that plants evolved a suite of floral traits, including color, scent and shape, in response to selection by particular groups of pollinators. For example, plants pollinated by hummingbirds evolved long red corolla, whereas moth pollinated flowers are generally white, open at night and emit a strong sweet fragrance (Proctor et al., 1996). Biologist still frequently use the pollination syndrome hypothesis to assign pollinators to plant flora without observing them in the field because of the assumed pollinator-floral trait associations. However, this hypothesis does not consider the morphological differences among species within a group of pollinators, nor the implications for floral evolution. Bees, the most important group of pollinators, have evolved various morphological traits to collect and transfer pollen, yet vary substantially at the genus and family level (Michener, 2007). For example, bees in the family Apidae are densely covered with plumose hairs, to which pollen readily adheres. Bees also vary in size, from less than 2 mm to nearly 40 mm (Michener, 2007). Such variation undoubtedly impacts their ability to transfer pollen. In order to determine if morphological differences among bees could influence plant-pollinator relationships, we measured bees collected from several communities in California and compared their traits at the family level. Variation in Bee Morphology Caitlin Hatmaker, Kelly McAfee, Nicholas Salinas, Sarah Scrivano, and Dr. Ruben Alarcón Biology Program, California State University Channel Islands, Camarillo, CA 93012 Figure 3. Relationship between body length and thorax depth. Figure 4. Relationship between body length and right forewing length. Figure 1. Measurements of Andrena duboisi were taken using ImageJ software: thorax width (A), intertegular distance (B), thorax depth (C) and right forewing (D). Figure 2. Bee families from left to right: Andrenidae (Andrena barbilabris), Apidae (Bombus californicus), Colletidae (Hylaeus mesillae), Megachilidae (Hoplitis fulgida), and Halictidae (Lasioglossum trizonatum). http://www.discoverlife.org/ Figure 5. Mean body length (±SE) for bee families. Lower case letters denote significant differences between means. Figure 6. Distribution of bee body hair across bee families.