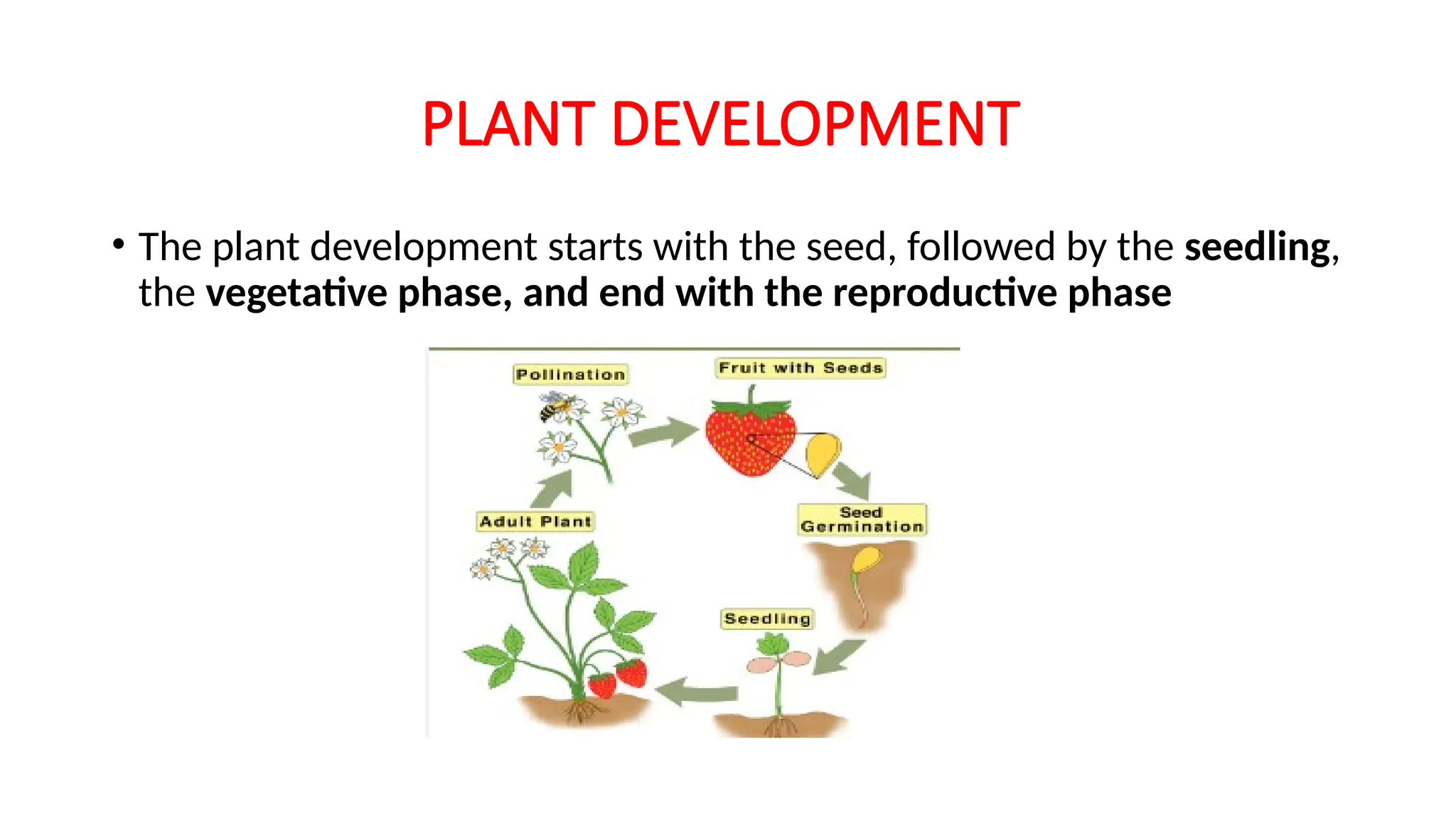
PLANT DEVELOPMENT
• Theplant development starts with the seed, followed by the seedling,
the vegetative phase, and end with the reproductive phase
2.
Seed Development
• Delouche(1971) defined seed development and maturation as a
process comprising of a series of morphological, physical,
physiological and biochemical changes that occur from ovule
fertilization to the time when seeds become physiologically
independent of the parent plant.
3.
Phases of seeddevelopment
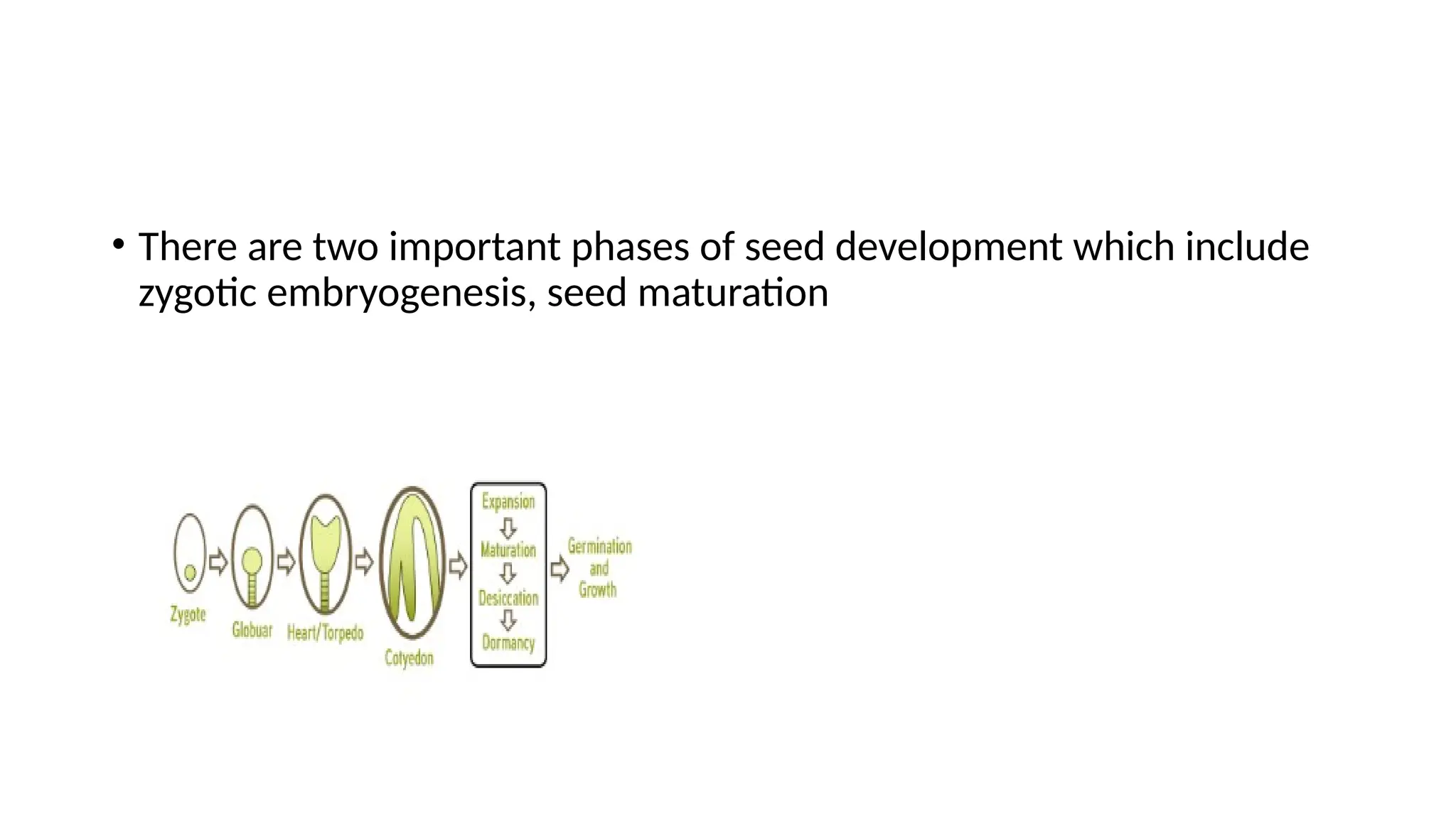
• The phases of seed development can be broadly categorized into two phases,
• A morphogenesis phase constituting phases from endosperm development,
cell divisions, and embryo and cotyledon differentiation and
• A maturation phase involving embryo growth at the expense of endosperm,
desiccation of the seed and accumulation of seed storage materials
• Seed development programs in major dicot seed crops such as soybean and
canola are similar to that in Arabidopsis
• Further, early stages of seed development in monocots are broadly similar to
dicots.
• However, later stages of seed development vary in monocots
4.
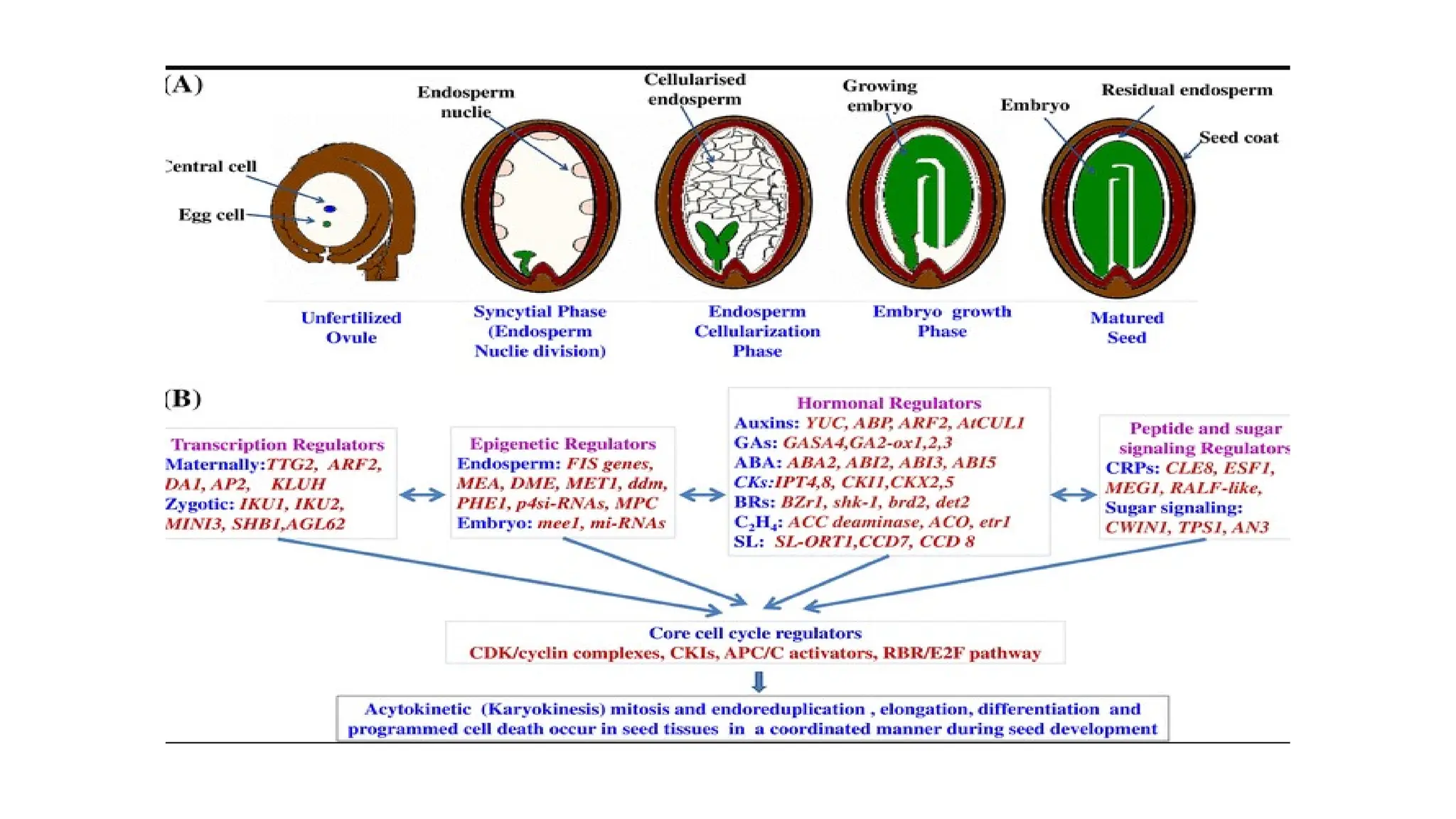
A morphogenesis phase
•Seed development begins with fertilization of both egg cell and central
cell to form the diploid embryo and triploid endosperm
• Followed by syncytial phase in which endosperm nuclei divide rapidly
without cell division and create seed cavity.
• In cellularization phase, rapidly divided endosperm nuclei are
cellularized and finally in embryo growth phase embryo grows at the
expense of endosperm to fill the seed cavity.
• A mature seed contains a single layer of endosperm and integuments
become seed coat.
6.
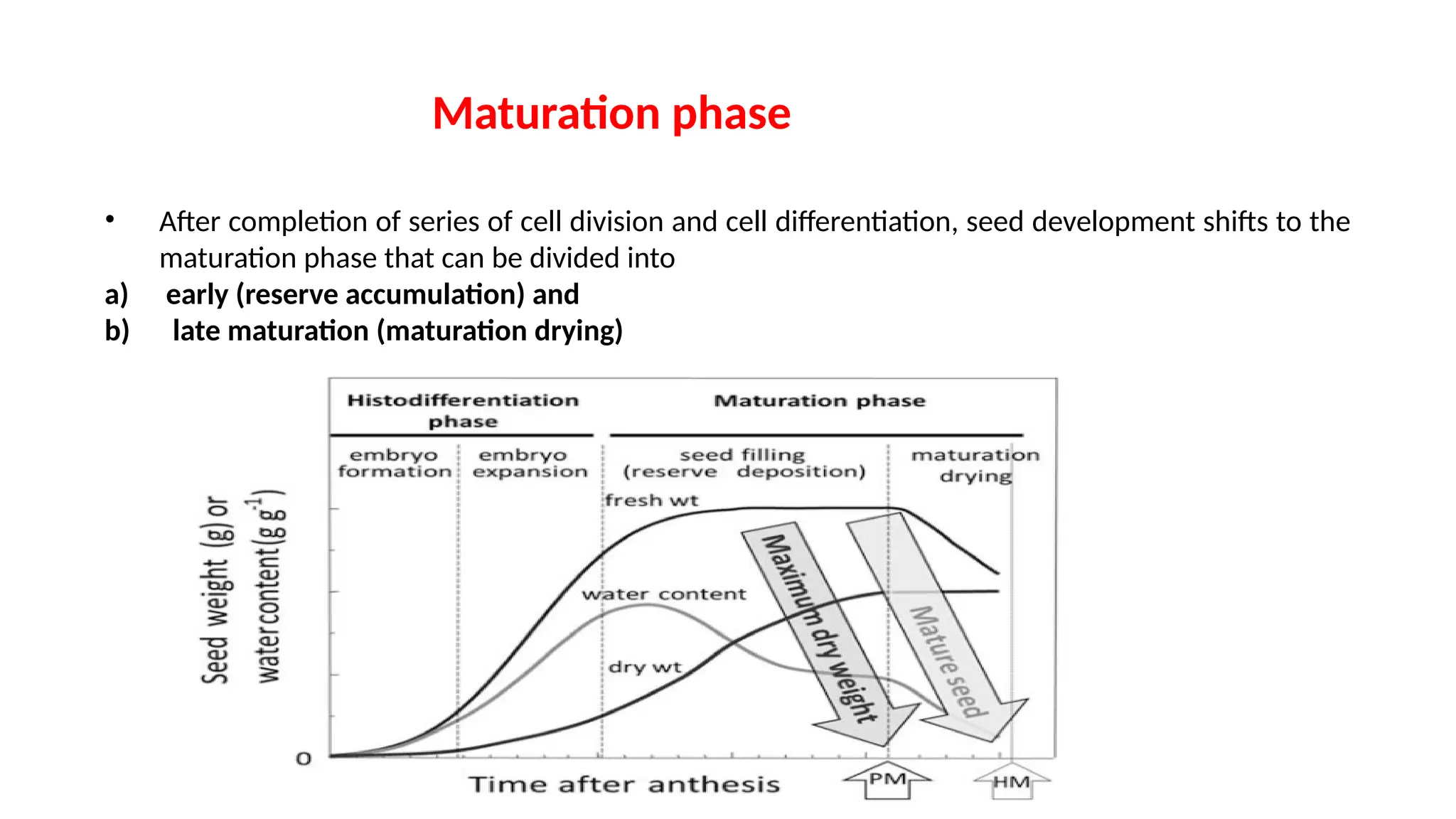
• After completionof series of cell division and cell differentiation, seed development shifts to the
maturation phase that can be divided into
a) early (reserve accumulation) and
b) late maturation (maturation drying)
Maturation phase
7.
Maturation Phase
• Seedmoisture content increases during the initial part of
development after fertilization and later begins to decline until
equilibrium is established with environmental factors
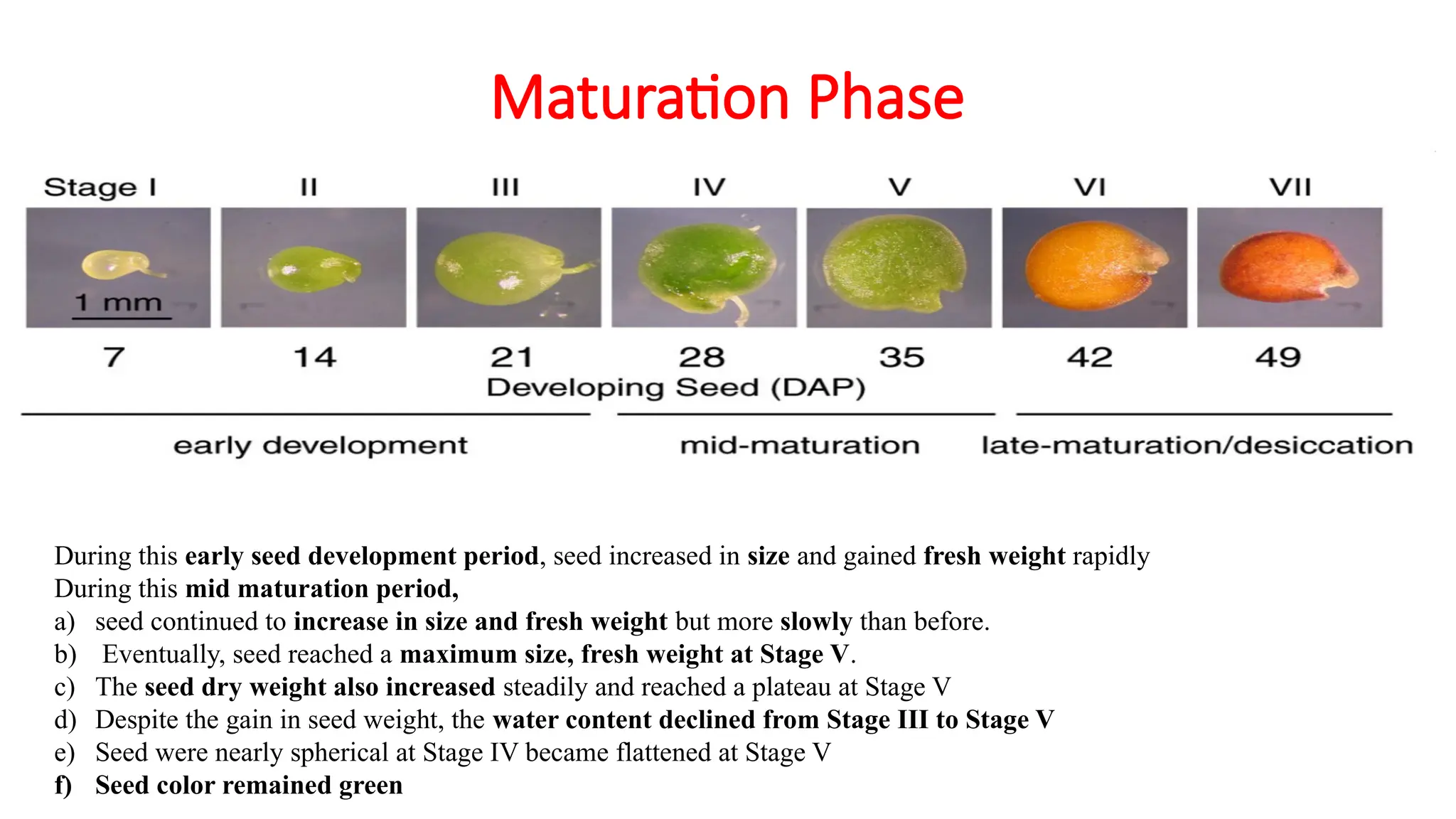
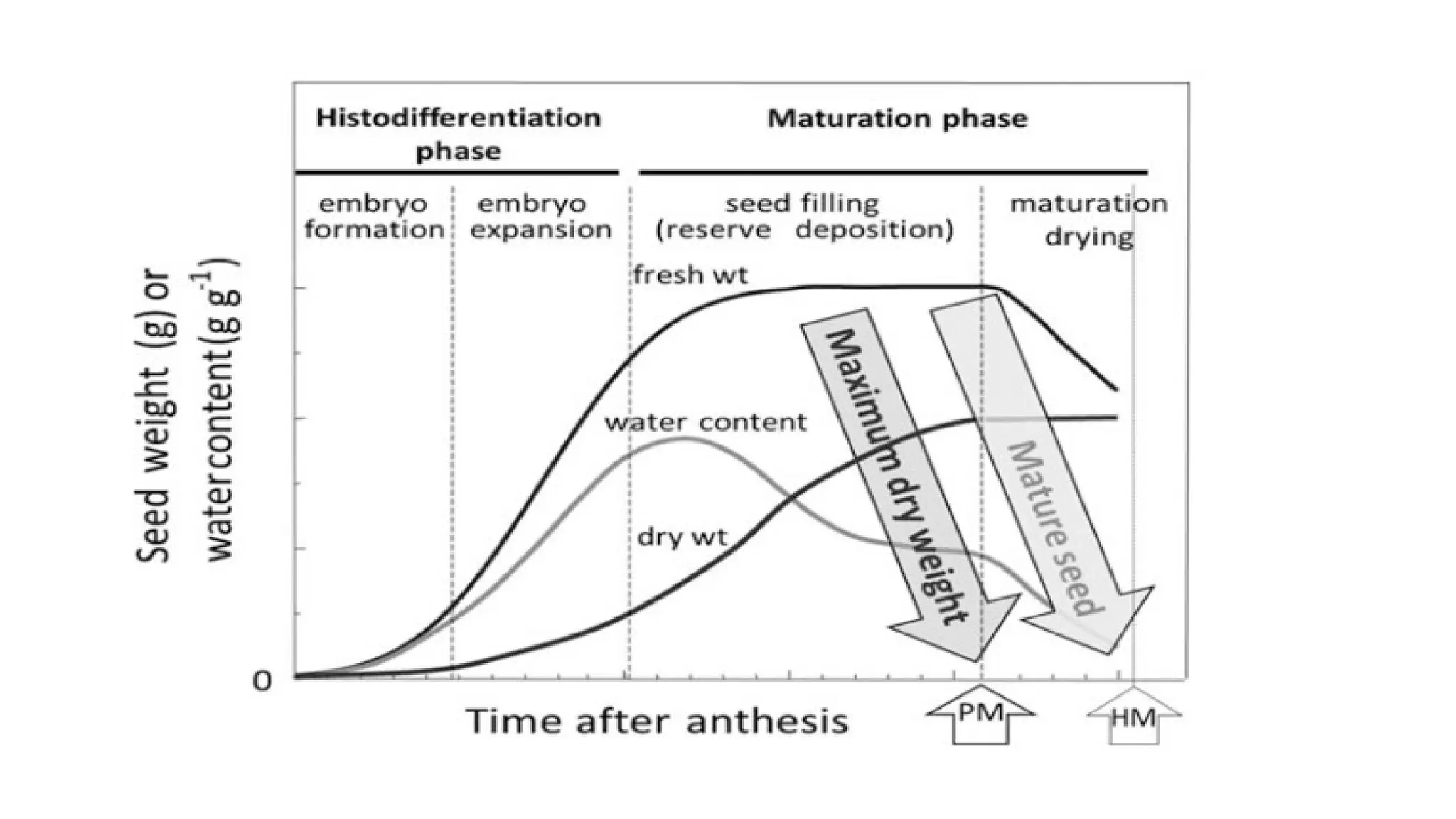
During this early seed development period, seed increased in size and gained fresh weight rapidly
During this mid maturation period,
a) seed continued to increase in size and fresh weight but more slowly than before.
b) Eventually, seed reached a maximum size, fresh weight at Stage V.
c) The seed dry weight also increased steadily and reached a plateau at Stage V
d) Despite the gain in seed weight, the water content declined from Stage III to Stage V
e) Seed were nearly spherical at Stage IV became flattened at Stage V
f) Seed color remained green
8.
• During thislate-maturation/desiccation period,
• seed had constant dry weight but water content dropped dramatically from Stage
V to Stage VII
• They were reduced in size and remained flattened.
• The color of seed was orange–brown at Stage VI and dark orange–brown at
Stage VII.
• During early maturation, the seed can acquire desiccation tolerance.
• Desiccation tolerance is defined as the ability of a living entity to deal with
extreme moisture loss to levels below 0.1 g water per gram dry weight, or drying
to relative humidity below 50%, and subsequent re-hydration without
accumulation of lethal damage
10.
• Physiological maturitywhen the dry weight reaches its maximum and that after
this stage, the flow of nutrients to seeds from the mother plant generally ceases.
• The late maturation phase is also often called the maturation drying phase.
• A significant decline in seed moisture content occurs at the end of maturation,
whereas the acquisition of desiccation tolerance begins during mid and late
maturation
• At the same time, massive structural and physiological changes occur within the seed,
• Structural changes: accumulation of reserve food, seed coat development structural
dormancy (impermiable seed coat) longevity & viability.
• A strong reduction in metabolic activity and a transition to a quiescent (temporary
inactivity & reversible) and frequently dormant state (temporary suspension due
to avoid stresses) at the end of late maturation.
Maximum dry weight
11.
• Based onthe desiccation tolerance, seeds can be classified into
a) orthodox and
b) recalcitrant types
Recalcitrant types
• Recalcitrant seeds usually do not show this quiescent state .( cant
withstand dessication) – no maturation drying phase, storage
limitations, short term plantings
• Recalcitrant seeds are sensitive to dehydration and desiccation leads to
damage and loss of viability
• Eg: rubber, avacado, Cacao
12.
Orthodox Types
• Thematuration in the orthodox seeds is accompanied with a water
loss up to 5–10% w/w, which allows them to sustain unfavourable
environmental conditions, such as extremely high and low
temperatures and drought.
• In orthodox seeds, the mechanisms behind the onset of desiccation
tolerance are activated at the early stages of maturation
• Later on, desiccation tolerance is lost during germination, at the
moment of radicle emergence
13.
How dessication toleranceis acquired?
Desiccation tolerance is acquired by seeds through accumulation of an
array of small molecules and proteins
maintain the structural integrity of critical cellular organelles,
membranes and
proteins
• They can persist during the dry state and resume their biological
functions upon hydration.
14.
• The embryoaccumulates specific molecules that are associated with
the cells’ ability to tolerate extreme water stress viz.
a) low molecular weight antioxidants,
b) oligosaccharides such as
c) raffinose,
d) stachyose,
e) late embryogenesis abundant proteins (LEAs) and
f) heat shock proteins (HSPs).
15.
• Further, structuralchanges occur at the cellular level such as folding
of cell walls, condensation of chromatin and dismantling of
thylakoids in chloroplasts
• These physiological and structural changes reduce metabolic activity
while mitigating the mechanical stress of cell shrinkage during
dehydration (maturation drying process).
• Changes at this stage correspond with a gradual increase in seed
longevity
16.
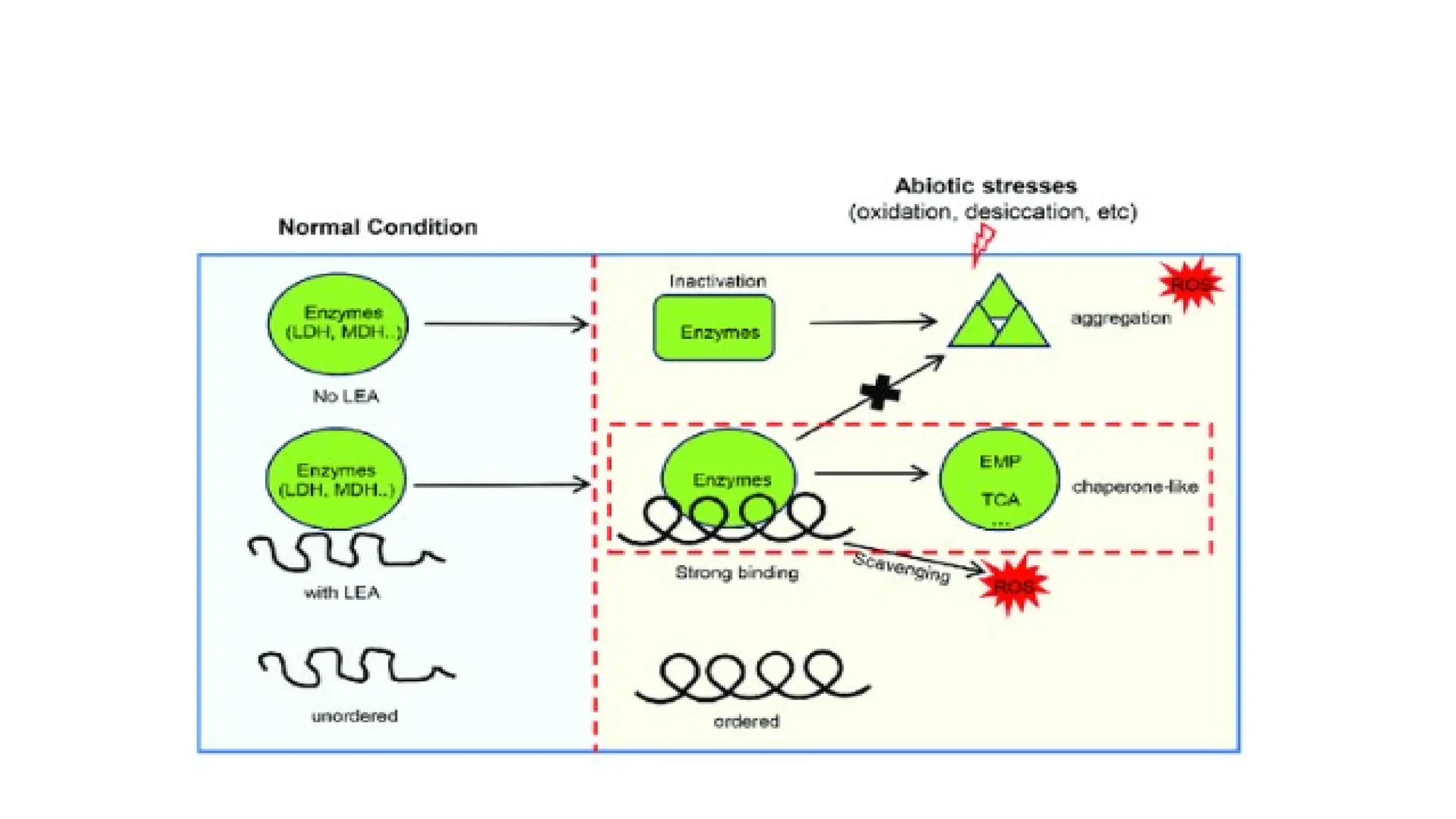
late embryogenesis abundantproteins (LEAs)
• LEA proteins have relative high content of glycine, alanine, glutamate,
lysine, arginine and threonine, while low amounts of cysteine and
tryptophan residues.
• Due to this primary nature, LEA proteins are stable in a broad
temperature range.
• During cell dehydration, LEA proteins act as chaperons, i.e., involved in
structural stabilization of denatured proteins and promote their
refolding through intensive hydrogen bond formation.
• LEA proteins are also responsible for sequestration of ionic
compounds, accumulating during cell dehydration, and protection of
membrane proteins and enzymes from the deleterious effects
18.
• Stresses usuallycause protein dysfunction.
• Maintaining proteins in their functional conformations and
preventing the aggregation of non-native proteins are particularly
important for cell survival under stress.
• Chaperones are responsible for protein folding, assembly,
translocation and degradation in many normal cellular processes,
stabilize proteins and membranes, and can assist in protein refolding
under stress conditions.
• They can play a crucial role in protecting plants against stress by re-
establishing normal protein conformation and thus cellular
homeostasis
19.
• Other structuraladaptations that occur during this stage are chromatin
compaction and nuclear size reduction, which are reversed during
germination.
• Furthermore, metabolic activity is reduced and chlorophyll is degraded
towards the end of seed maturation thereby minimizing the production of
reactive oxygen species (ROS).
• Non-reducing sugars fill the free volume between large molecules, created
during dehydration and the dehydrated cytoplasm forms a glassy matrix
with very low molecular mobility.
• To protect the seed against oxidative damage, which cannot be repaired by
enzyme activity under dry conditions.
• Seeds accumulate during seed maturation many antioxidants, such as
ascorbate, glutathione, polyols, tocopherols, quinones, flavonoids and
phenolics
20.
Group II LEAproteins
• Group II LEA proteins accumulate highly in plant embryos during the late stages
of seed development as an aid to embryo maturation under desiccation.
• In plant vegetative tissues, group II LEA proteins are rarely detected and are
limited to young parts of plants, especially those that exhibit excessive cell
division and cell elongation, for example, at the root tips, in expanding stems, and
in petioles.
• Some group II LEA proteins are found in mature seeds.
• Localized:
all the segments of the embryo and endosperm of mature seeds.
• accumulates in developing cotyledons during mid-to-late embryogenesis and in
seedlings during dehydration stress
21.
• It comprisesabout 2% of the proteins in mature cotyledons.
• The carrot group II LEA gene, ECP40, is distributed in the zygotic
embryos and endosperm of mature seeds
• In seed physiology, DHNs or group II LEA proteins are considered to
be responsible for the persistence and longevity of seeds.
• Plant seeds are of special interest for investigating the proteins from
the group II LEA family, since they are relatively abundant during seed
maturation stages and in response to any external stimulus causing
dehydration to the seeds .
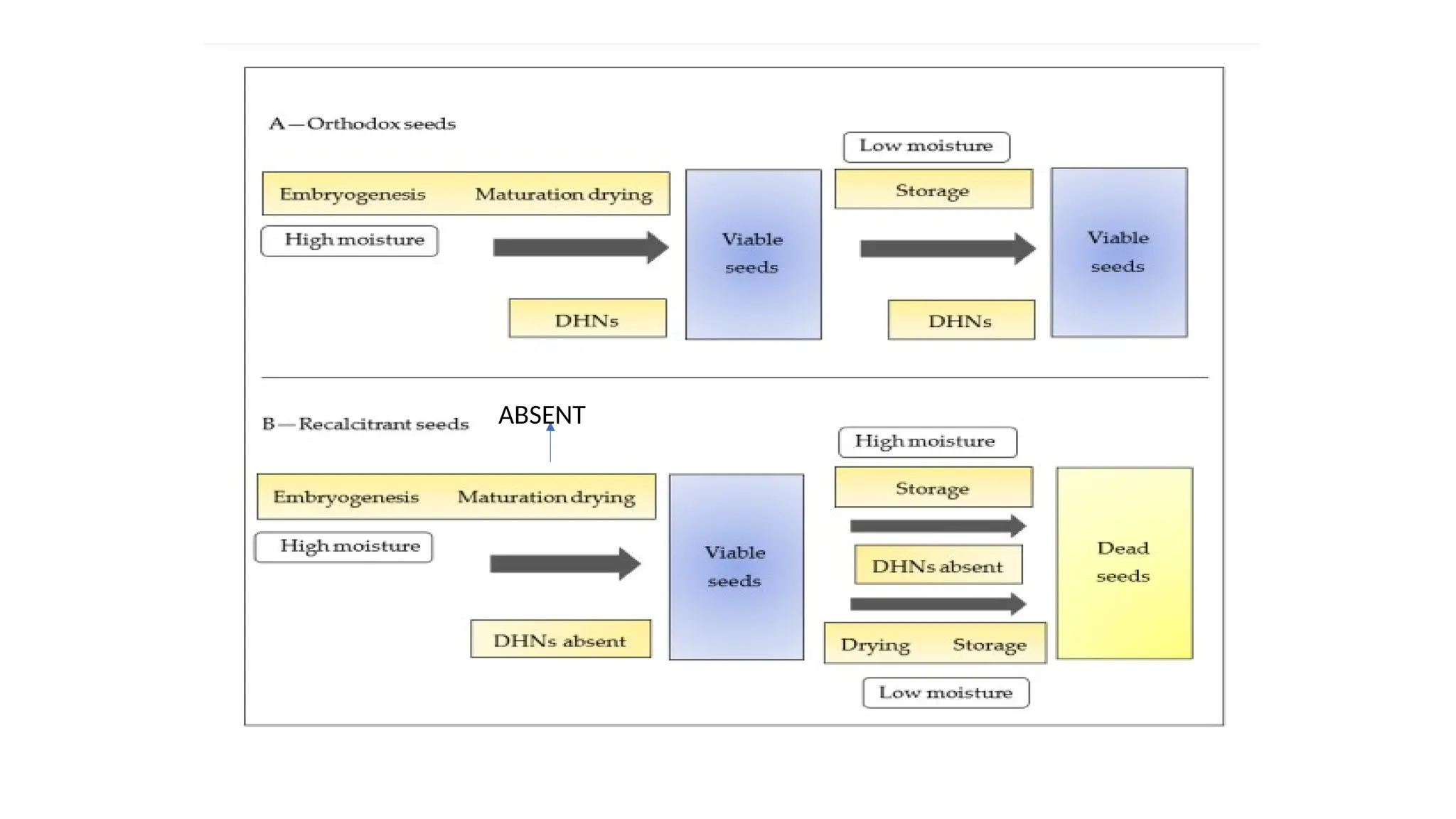
• Seeds are classified as recalcitrant or orthodox based on their storage
behaviors
22.
• Recalcitrant seedsdo not go through maturation drying and drop with
a relatively high content of moisture
• Seed recalcitrance is a major issue for the natural production of plant
species that causes a serious problem in seed conservation and storage
• In recalcitrant seeds, a positive correlation was found between the seed
moisture content and the germination rate
• These seeds cannot be maintained and stored in conventional freezers
due to their low survivability after drying and freezing at −20 ◦C.
• The absence of resistance in recalcitrant seed drying was attributed to
the lack of DHNs
23.
• Orthodox seeds,on the other hand, go through maturation drying
and are dropped from plants at a low content of moisture
• These seeds have the potential to be dried to an internal seed water
content of less than 12% and can be maintained, stored, and survived
at freezing temperatures
• DHNs are synthesized in orthodox seeds, which are accumulated
during the final stages of maturation and during seed desiccation
• It has been suggested that, in orthodox seeds, DHNs favor their
tolerance towards moisture loss and osmotic stress during the stage
of seed maturation
24.
• In orthodoxseeds, as a response to the maturation drying, DHNs are
synthesized, whereas recalcitrant seeds do not synthesize DHNs
because of the absence of any maturation drying
• It is because of DHNs that orthodox seeds retain their viability during
storage whereas recalcitrant seeds become unviable because of the
absence of DHNs during maturation drying and storage conditions.
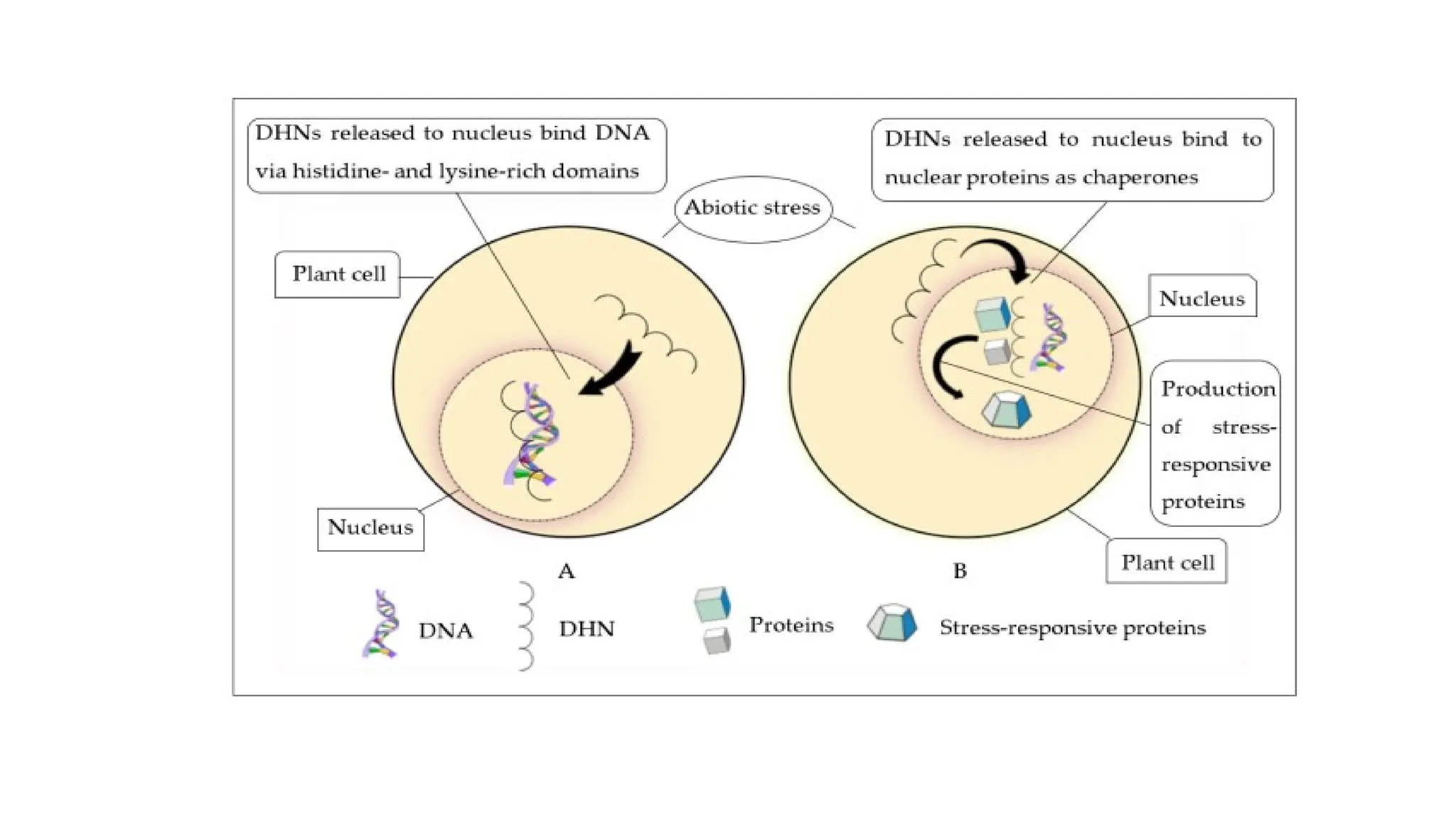
Functional mechanism
• Aschaperones and bind to DNA and other protein biomolecules by shielding
them and thereby preserve the functions of these proteins during the stress.
• The ability to bind to DNA and protect it from immoderate ROS such as H2O.
• DHNs preserve the activities of lactate dehydrogenase (LDH) and malate
dehydrogenase (MDH) under freezing and thawing stress damage.
• Such stabilization of cell structures and organelles was evident through the
overexpression of DHN genes in transgenic tomatoes, which improved the
relative water content (RWC) and lowered the rate of water loss in the
tomato
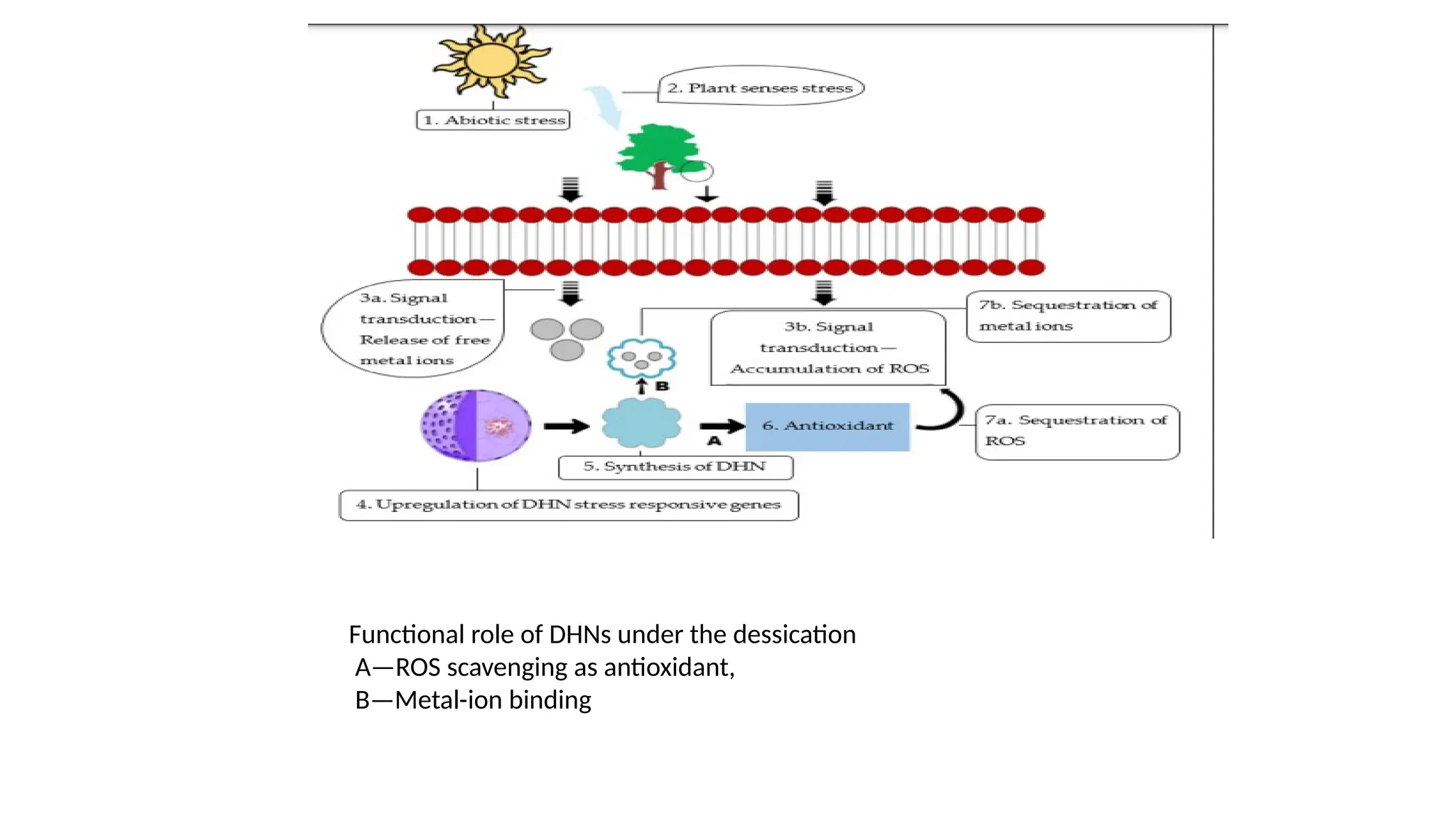
28.
• Under thedessication, the plant senses the stress and signals its
organelles for the presence of stress through the release of free metal
ions or through the accumulation of ROS.
• After the signal transduction, DHN genes are upregulated within the
nucleus, and DHNs are synthesized for the stress tolerance
mechanism.
• Both phenomena can occur within plant cells based on the signal
transduction pathway.
• (A) DHNs act as antioxidants and scavenge ROS that accumulate
within the plant cells.
• (B) DHNs, through their property of metal-ion binding, also scavenge
the free metal-ion radicals that arise within plant cells under abiotic
stress.
29.
Functional role ofDHNs under the dessication
A—ROS scavenging as antioxidant,
B—Metal-ion binding
30.
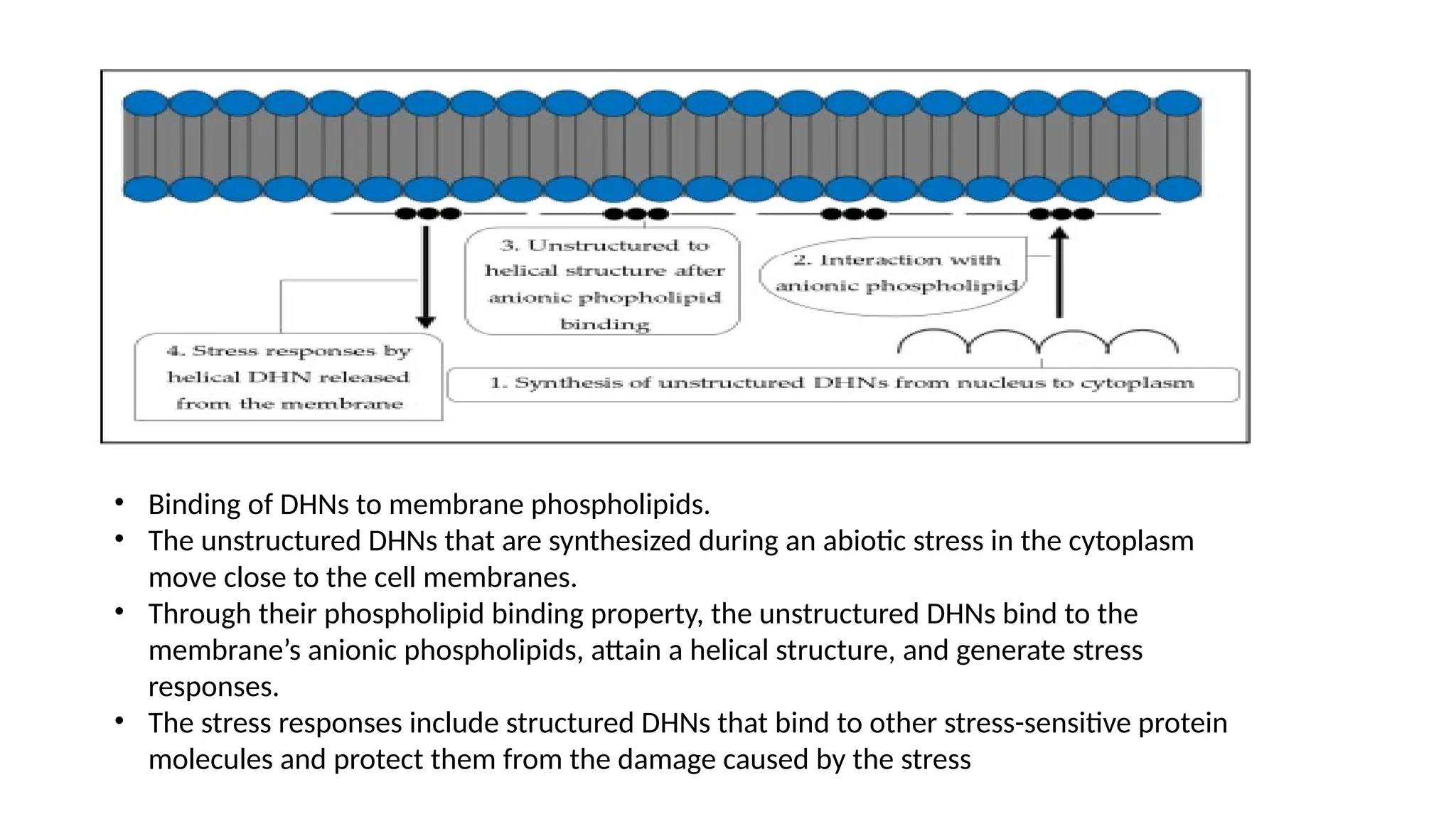
• Binding ofDHNs to membrane phospholipids.
• The unstructured DHNs that are synthesized during an abiotic stress in the cytoplasm
move close to the cell membranes.
• Through their phospholipid binding property, the unstructured DHNs bind to the
membrane’s anionic phospholipids, attain a helical structure, and generate stress
responses.
• The stress responses include structured DHNs that bind to other stress-sensitive protein
molecules and protect them from the damage caused by the stress
31.
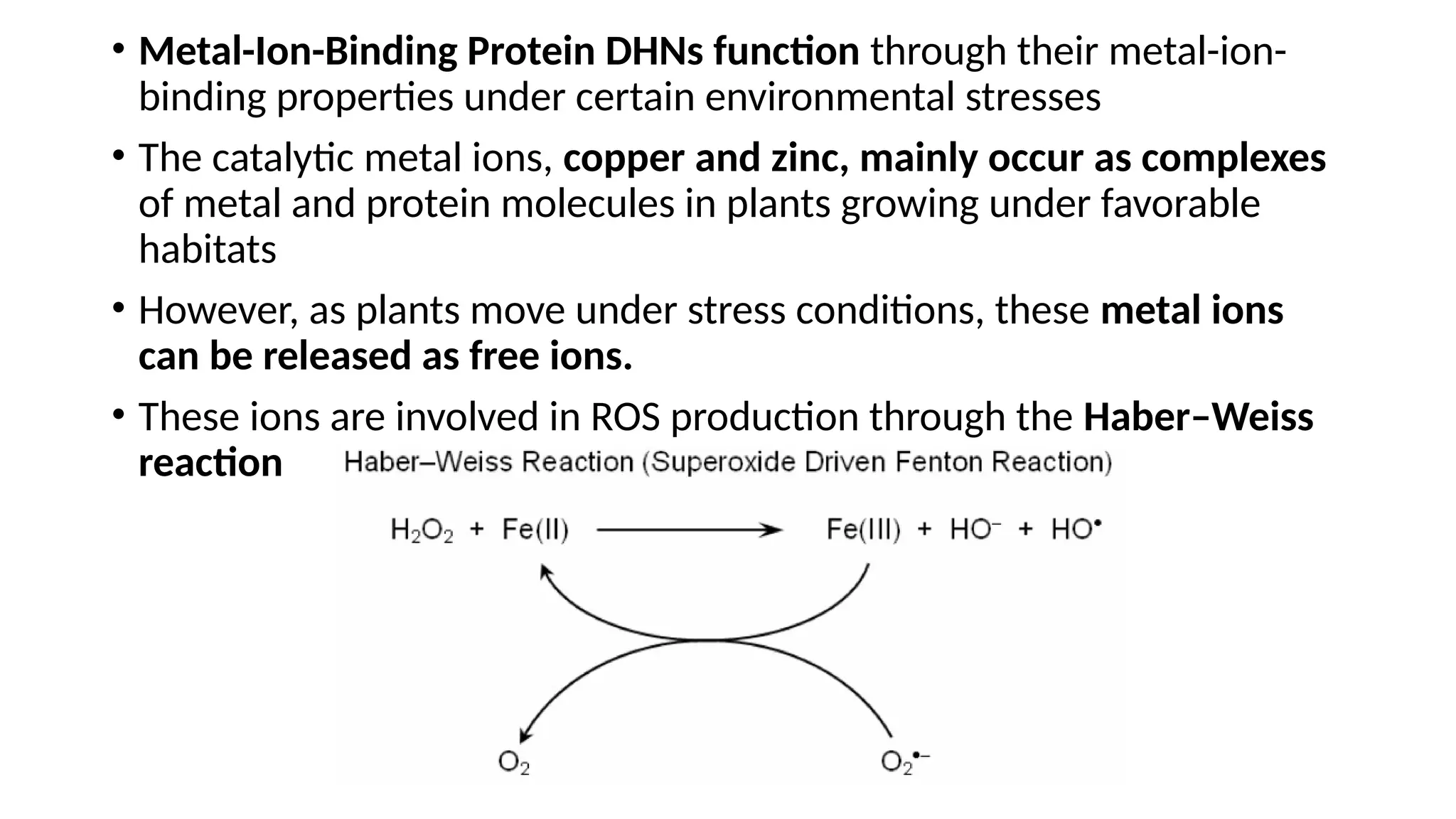
• Metal-Ion-Binding ProteinDHNs function through their metal-ion-
binding properties under certain environmental stresses
• The catalytic metal ions, copper and zinc, mainly occur as complexes
of metal and protein molecules in plants growing under favorable
habitats
• However, as plants move under stress conditions, these metal ions
can be released as free ions.
• These ions are involved in ROS production through the Haber–Weiss
reaction
32.
• Metal ionsare a common target for a number of DHNs.
• Abiotic stresses, such as water stress, result in the release of metal ions from
the membranes and organelles and increase the concentration of free
metals in the intracellular spaces
• It has been hypothesized that DHNs bind to these free metal ions and
decrease the damage they cause
• The binding of DHNs to metal ions has been reported in Arabidopsis thaliana
and citrus DHNs, AtHIRD11 and CuCOR15, which are able to bind to iron and
cobalt over magnesium and calcium and prevent the release of free ions
• It has also been found that CuCOR15 acts as a radical scavenger that
reduces the metal toxicity in plants under drought stress
• Moreover, an ion transport protein (ITP), KS-DHN, from Ricinus communis
was indicated as an active transporter of metal ions within plants.
33.
• Phospholipid-Binding ProteinDHNs tend to bind to phospholipids because of their rich K-segments
and histidine motifs
• Their binding to phospholipids triggers the accumulation of a crucial stress signaling phospholipid,
phosphatidic acid
• The concentration of PA in an inflated plasma membrane is very low, about 1%, but increases under
drought stress
• The increase in PA concentration is due to low water content within cells or release of ABA
• The presence of basic amino acids such as arginine and lysine in DHNs enables them to bind to
anionic phospholipids
• The interaction between dehydrins and membranes changes certain membrane properties, such as
water content and temperature within cells
• DHNs bind to charged lipids by the occurrence of electrostatic interactions .
• Some DHNs gain their helicity structure through binding with acidic phospholipids
• This enables them to bind to other biomolecules within the cytoplasm and protect them from stress
• As DHNs bind particularly to acidic phospholipids, it can be postulated that DHNs may interact with
membranes of the cell at specific regions
34.
Hormonal Regulation ofSeed Development
and Maturation
• Plant hormones are signal molecules that are produced in the plant and are active
at very low concentrations.
• The hormones abscisic acid (ABA), gibberellins (GAs), auxin (IAA), cytokinins,
ethylene and brassinosteroids regulate cellular processes in targeted cells, which
may or may not be the cells in which they are synthesize.
• Measurements of endogenous hormone concentration have suggested the high
transient expression of cytokinins, GAs and IAA during the early phase of seed
development
• In studies with tomato, GAs were found essential to produce fertile pollen, but
pollination of a GA-deficient (female) mutant with mutant pollen, obtained upon
spraying the male plant with GAs, resulted in the development of normal-looking
healthy seeds that only needed GAs for germination
35.
• Cytokinins havealso been implicated in promoting suspensor
function, but may be even more significant in promoting endosperm
growth and grain filling via promotion of cell division (Bewley et al.
2013).
• In contrast, during early embryogenesis, auxins play a major role in
establishing the embryonic body plan via effects on apical-basal
polarity or pattern formation (transition of embryo from globular to
heart shape and cotyledon separation at later stages) and vascular
development
36.
• During maturation,seeds of most species acquire the capability to endure
desiccation.
• The maturation phase begins when the embryo and endosperm have
accomplished the morphogenesis and patterning stages (Wobus and Weber 1999).
• This phase is categorized by a growth arrest, followed by the synthesis and
accumulation of reserves, whose degradation upon germination will provide
nutrients to the growing seedling before the photosynthetic capacity is fully
acquired (Baud et al. 2002)
. Early and mid-phases of maturation are controlled by the action of ABA, initially
synthesized in the maternal tissues and later on, although to a lower extent, in the
embryo and endosperm (Nambara and Marion-Poll 2003).
Seed maturation coincides with an increase in seed ABA content; consistent with
the fact that ABA induces expression of a cyclin-dependent kinase inhibitor (ICK1)
that could lead to cell cycle arres
37.
• In thelater stage, a decline in ABA level occurs and synthesis of LEA
proteins follows, which is characteristic to the late maturation phase.
• Maturation is not always an obligatory process, if ABA effects are
eliminated by removing the embryo from the seed would lead to
development of seedlings (Berger 2003).
• But due to their low vigour, planting these immature seeds in the
field will not result in the development of a healthy seedling.
38.
Role of ABAbiosynthesis genes in seed
development
• Maternal ABA plays a significant role in embryo development and seed
maturation.
• ABA is also de novo synthesized in embryo and testa during embryo
development, as well as accumulates during seed maturation,
• Facilitates late seed maturation processes,
• Synthesis of storage proteins to prevent seed abortion,
• Induces primary dormancy and allows successful germination as well as a
successive seedling enterprise
• So, de novo synthesis of active ABA plays a more important role in seed
development and later germination
39.
• Active ABAis synthesized through an indirect pathway from
xanthophylls (e.g., zeaxanthin, violaxanthin, and neoxanthin).
• Three types of genes are responsible for the successive steps of ABA
biosynthesis such as ZEAXANTHIN EPOXIDATION (ZEP), OXIDATIVE
CLEAVAGE OF 9-CIS-EPOXYCAROTENOIDS (NCED), and ABSCISIC
ALDEHYDE OXIDATION (AAO).
• The ZEP/ABA gene was firstly identified in Arabidopsis thaliana and
Nicotiana plumbaginifolia.
40.
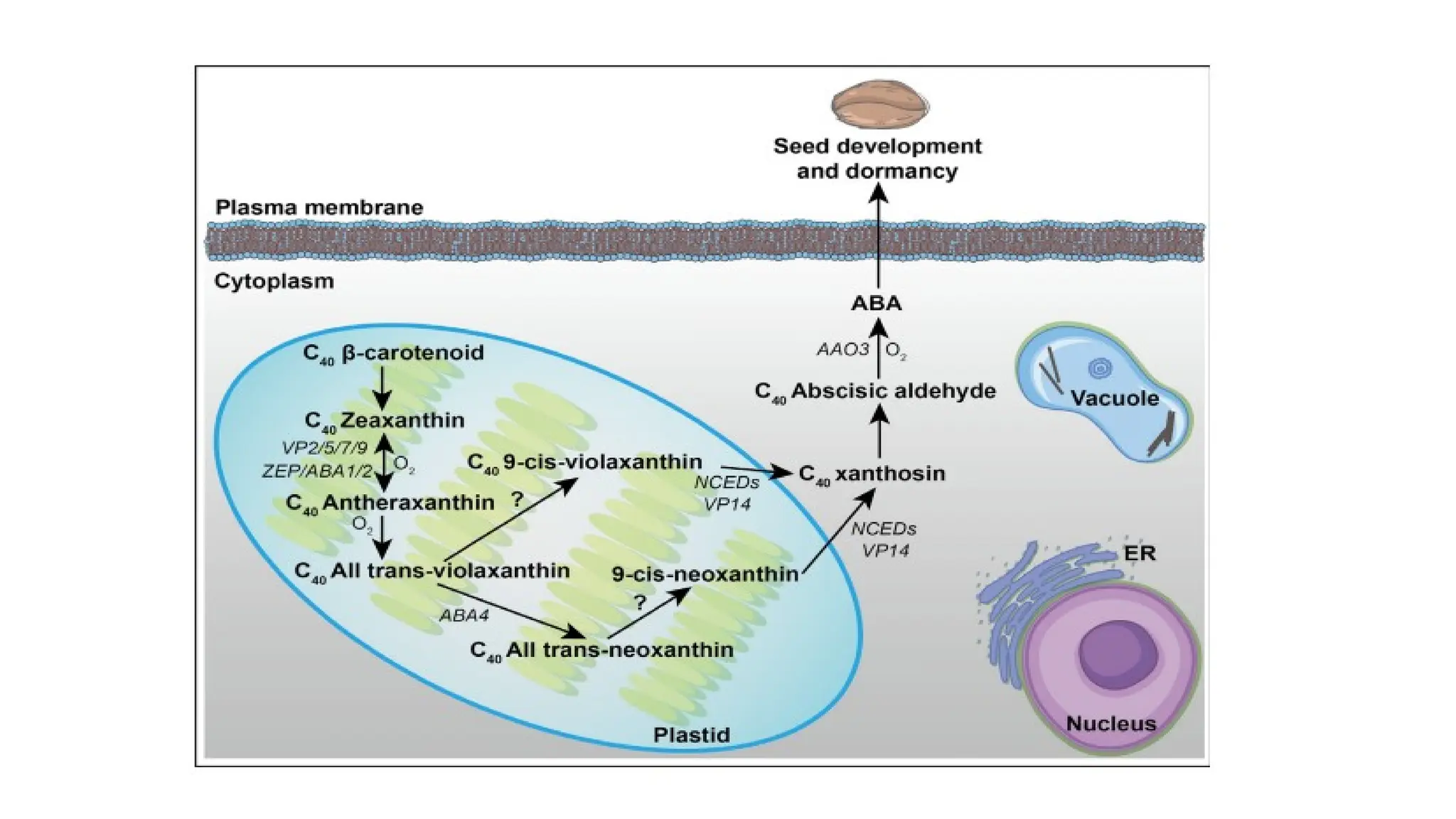
• Regulation ofseed development and dormancy by ABA biosynthesis through the carotenoid
pathway started from b-carotene (C40).
• The complete ABA synthesis process takes place in plastids and cytoplasm where ZEAXANTHIN
EPOXIDASE (VPs, ZEP, ABA1/2) converts zeaxanthin into antheraxanthin and all trans-violaxanthin.
• ABA4 catalyzes the conversion from all-trans-violoxanthin to the all-trans-neoxanthin.
• The conversion of xanthoxin from 90 -cis-neoxanthin and 90 -cis-violaxanthin is exerted by VP14 and
NCEDs (NINE-CIS-EPOXYCAROTENOID DIOXYGENASE), among which the NCEDs display different
subcellular localization of plastid or cytoplasm.
• The oxidation of abscisic aldehyde by AAO3 (ABSCISIC ALDEHYDE OXIDASE3) is responsible for the
conversion from abscisic aldehyde into ABA, which in turn induces and maintains seed dormancy..
42.
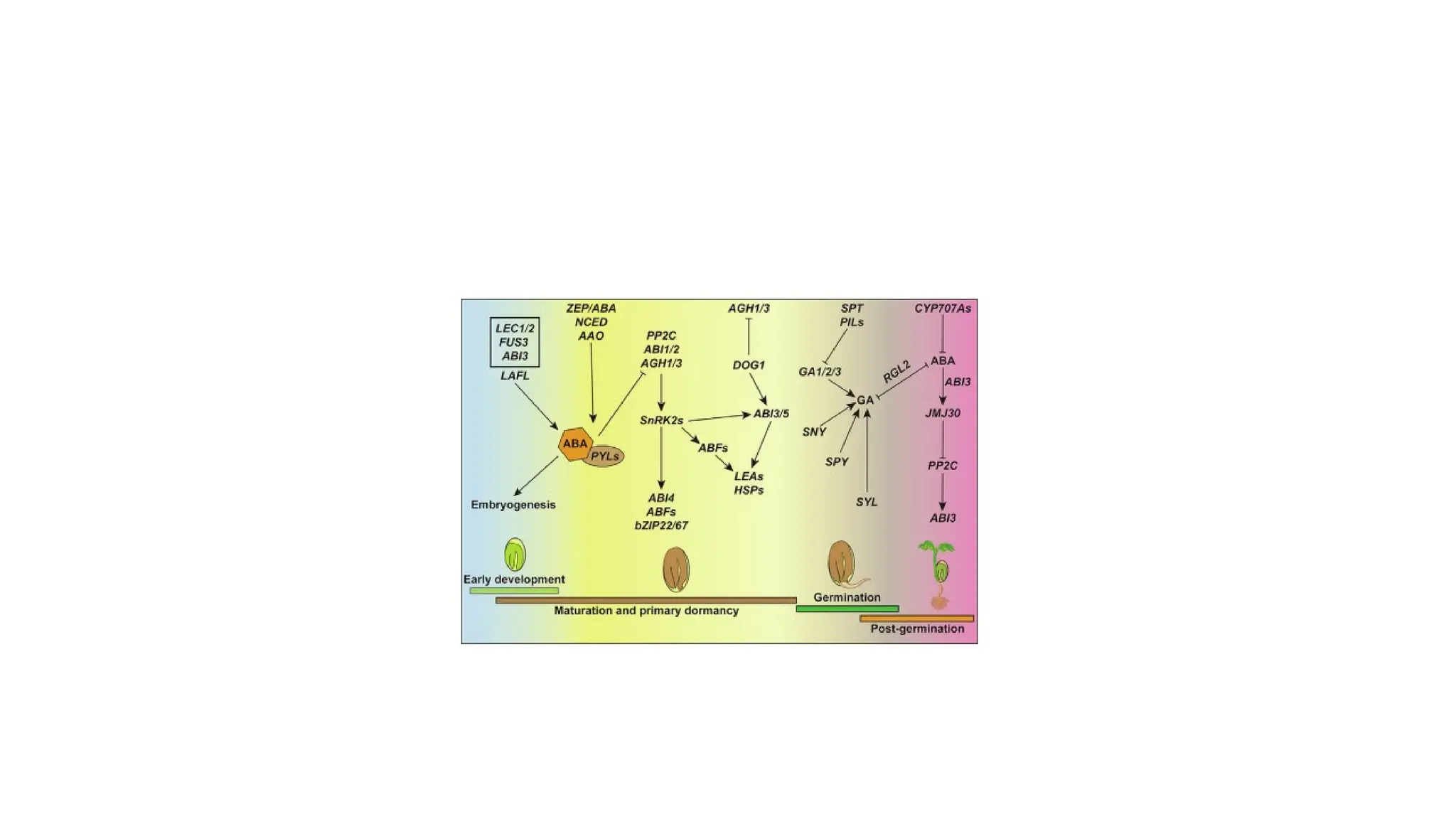
Role of ABAsignaling components in different
seed developmental stages
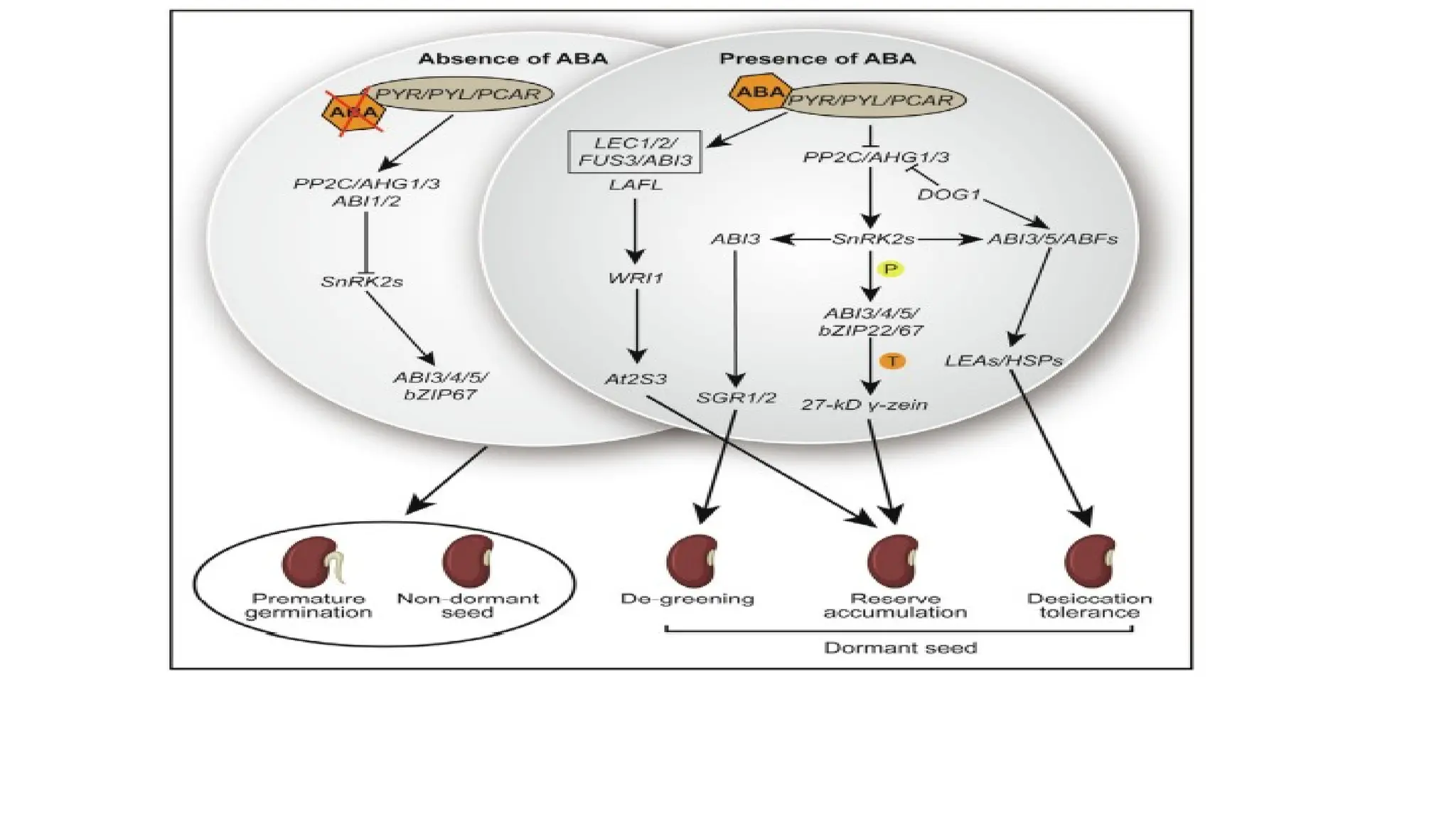
• The ABA signaling pathway is involved in seed development.
• In the absence of ABA: Receptors PYLs release and activate protein phosphatase 2C (PP2C) such
as ABI1/2 and AGH1/3.
• Downstream SNF1-RELATED PROTEIN KINASE subfamily (SnRK2s) genes are inactivated by active
PP2C which leads to premature germination and the nondormant seed through repression of lots
of transcription factors such as ABI1/2/3/4/5 and bZIP67.
• presence of ABA: Receptors PYR/PYL/RCAR bind ABA and PP2C together to inhibit the activity of
PP2C, which release the activity of SnRK2s
• downstream transcription factors such as ABI3 by protein phosphorylation, then regulate
downstream genes SGR1/2 function to mediate seed de-greening process.
43.
• Additionally, theactive LAFL (ABI3, FUS3, LEC1, and LEC2) network by ABA along with WRI1
regulates the At2S3 gene; an active bZIP22 function downstream of SnRK2s to promote
gene transcription of 27-kD c-zein for protein reserve accumulation in the seed.
• Along with seed de-greening and storage product accumulation, SnRK2s function upstream
of ABI3/5 and ABFs to regulate LEAs and HSPs that are pivotal for desiccation tolerance. In
other branches, DOG1 also plays a role upstream of ABI3/5/ABFs as well as functions as a
repressor of PP2Cs (AHG1/3) to involve seed desiccation tolerance acquirement.
• ABA signaling components (SnRK2s, ABI3, ABI4, ABI5, ZmbZIP22, bZIP67, and ABFs) are
involved in storage product accumulation, de-greening, and desiccation tolerance with
different function pathways to provide a mature and dormant seed.
• Letters ‘‘P” and ‘‘T” in the color circles indicate the two manners of protein
phosphorylation and gene transcription regulation, respectively. Activated and repressive
effects are shown by arrows and bars, respectively
45.
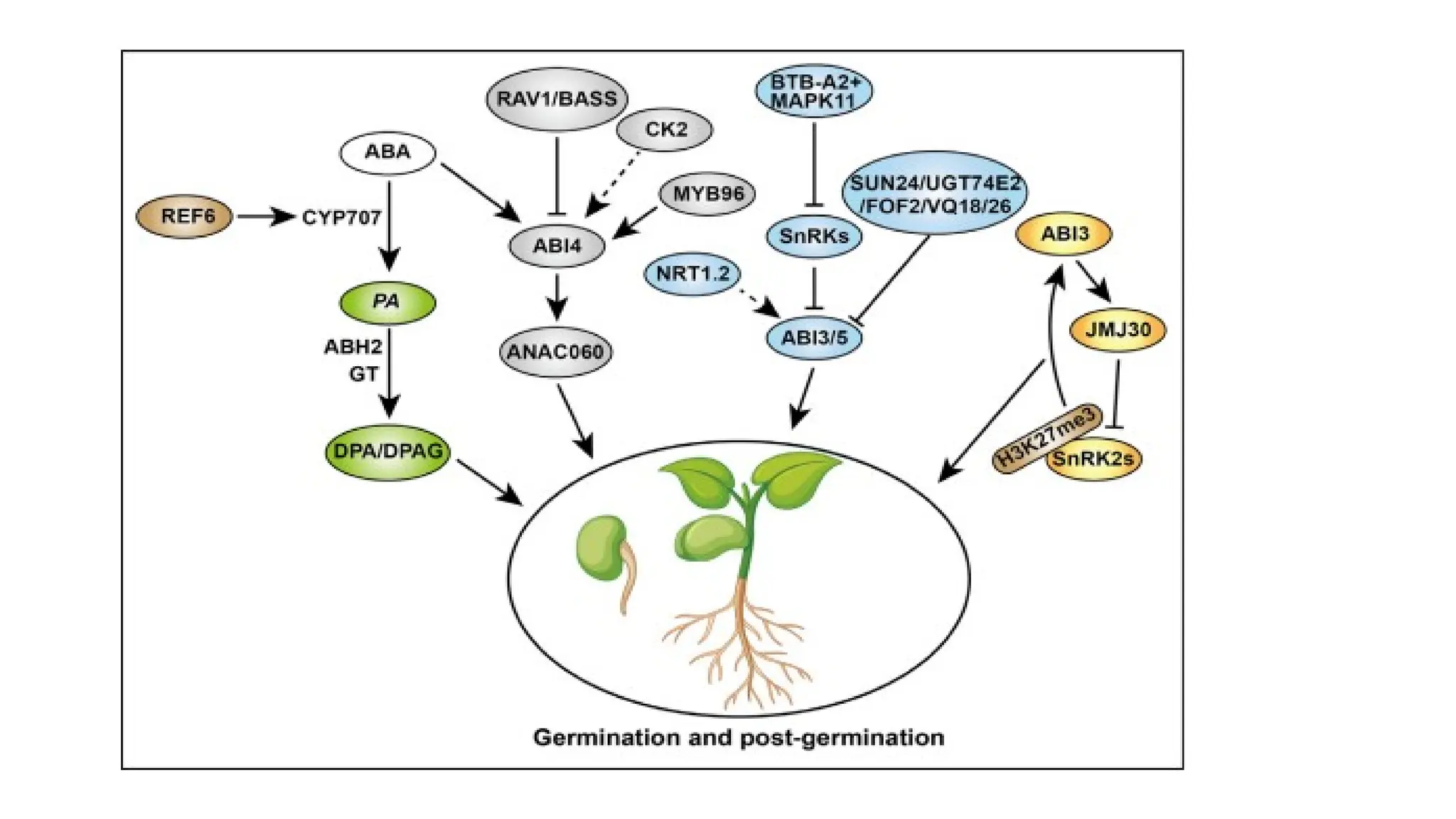
• The functionof ABA in seed germination and seedling establishment.
• Seed completes germination successfully through degradation of
active ABA into PA (phaseic acid) and DPA (dihydrophaseic acid)/DPAG
with CYP707As regulated by REF6 and phaseic acid reductase (ABH2
and GT) respectively.
• During germination and seedling establishment, the core ABA
signaling component SnRK2s and downstream ABI3/4/5 are activated
or repressed by many factors directly or indirectly to promote seed
germination and seedling establishment.
47.
Ethylene
• The ethylenepathway studies in relation to seed development and maturation
are extremely limited. I
• n plant tissues, ethylene affects chlorophyll metabolism (Matilla 2000).
• Because chlorophyll loss is triggered during the final stages of embryogenesis
(during acquisition of seed vigour), this process may be affected by ethylene.
• Mustard and canola seeds produce significant amounts of ethylene during
embryogenesis, specifically in the early pre-desiccation stages (Child et al.
1998).
• Hence the role of ethylene can be attributed as minor during seed
development and maturation and may be associated with the embryo de-
greening process.
48.
Heat shock proteins
•Higher plants are unable to cope up with the extended exposure to temperatures
above 45 °C
• cellular homeostasis
• The loss of biological activity of proteins upon high temperature stress may be
due to aggregation and/or protein misfolding
• The stress-induced accumulation of aggregated and mis-folded proteins is
irreversible and deleterious to the cell functioning.
• To balance the homeostasis of cellular proteins under heat stress, plant cell
upregulates several heat inducible genes, commonly referred as “heat shock
genes” (HSGs)
• chaperone activity
49.
Heat Shock proteins
•Hsps are broadly divided into two major families i.e.,
low and large molecular weight Hsps
sizes
Hsp100/Clp, Hsp90, Hsp70, Hsp60/chaperonin and sHsps
50.
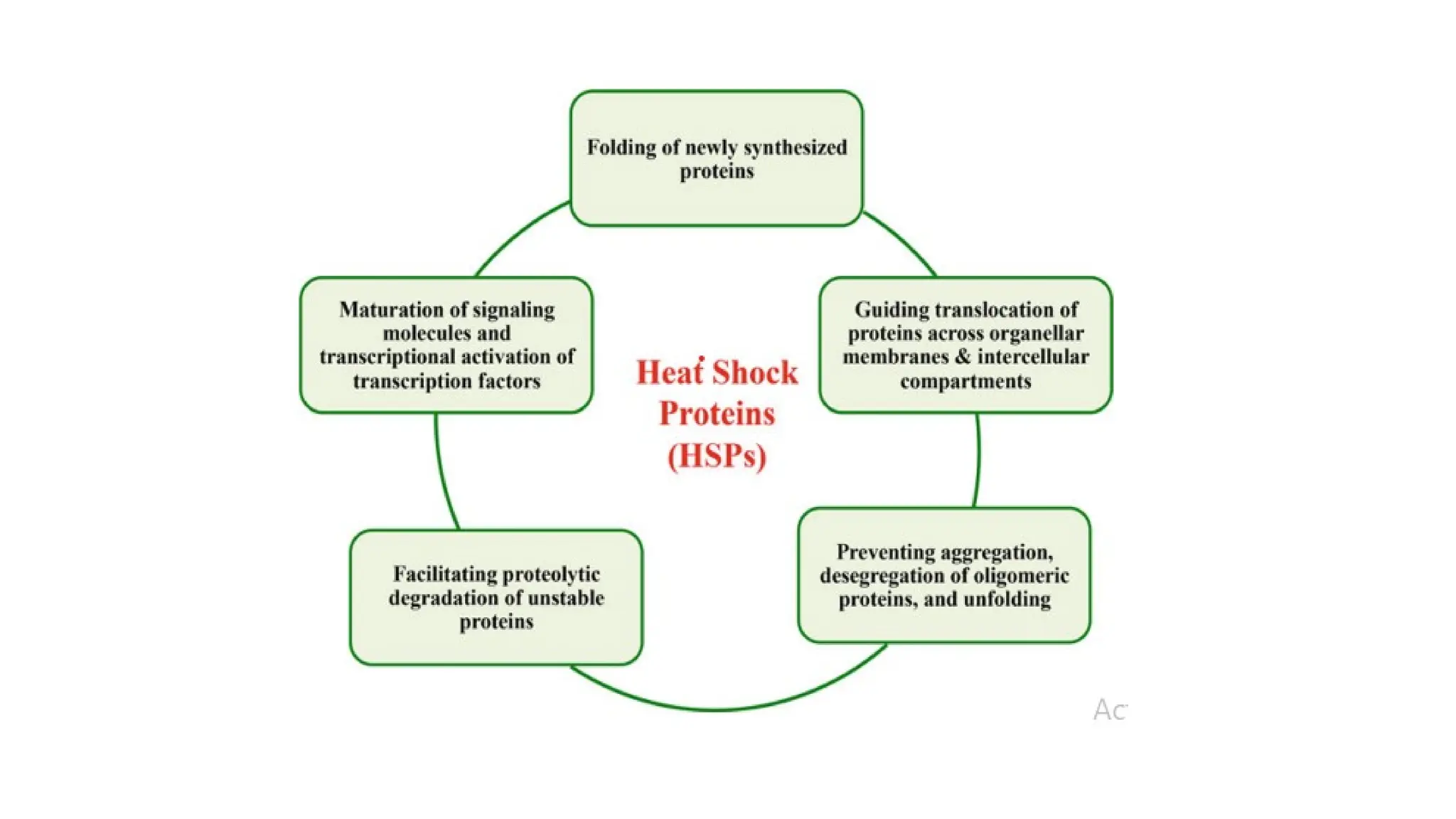
Functions
(1) folding orassisting folding of newly synthesized proteins (Hsp70,
Hsp60),
(2) guiding translocation of proteins across organellar membranes and
between intercellular compartments (Hsp70)
(3) preventing aggregation, desegregation of oligomeric proteins, and
unfolding (Hsp70, Hsp100, Hsp90, small Hsps)
(4) facilitating proteolytic degradation of unstable proteins
(Hsp70, Hsp100),
(5) maturation of signaling molecules, signal transduction and
transcriptional activation of transcription factors (Hsp70, Hsp90
52.
• sHsps arefound to be most prevalent in plants and their expression can be
increased up to 200 folds under stress
• sHsps range in size from 10 to 42 kDa
• localized in compartments like cytosol, endoplasmic reticulum (ER),
mitochondria and chloroplast
• sHsps do not actively participate in refolding of non-native proteins
• They possess a high capacity of binding to nonnative proteins, through
hydrophobic interaction
• The abundance of sHsps in plants and their functional characteristics of binding
and stabilizing denatured proteins suggest that sHsps play an important role in
plant-acquired stress tolerance
53.
• TaHSP26 isinduced by heat stress in vegetative and floral tissues in both wheat
and Arabidopsis.
• The induction of TaHSP26 at 37 °C is quite high and rapid (induced within 10 min)
in 10-day-old shoots and root tissues.
• The induction of sHSP26 is faster in wheat (present study) than in rice,
where reported 20 min of lag period in its induction upon high-temperature stress.
• Similar kinetics has also been reported for other plant small HSPs
• This may be indicative of the fact that wheat is a temperate crop while rice is a
predominately sub-tropical crop.
• Additionally, TaHSP26 transcript also accumulated in early stages of seed
development upon heat stress, followed by constitutive accumulation at advanced
stages of seed maturation when it undergoes the desiccation phase.
• Transcript accumulation also takes place in Arabidopsis seedling tissues,
inflorescence and developing siliques upon heat stress during early and later stages;
however, unlike wheat, we did not find accumulation of AtHSP26 transcript
in Arabidopsis siliques under non-stressed conditions
54.
Physiological Maturity, MassMaturity and
Harvest Maturity
• Two stages of maturity have been defined viz. physiological (or mass)
maturity and harvest maturity.
• Physiological maturity is the end of the seed-filling period (Harrington
1972), whereas harvest maturity is the point of time that coincides
with the end of maturation drying
55.
Seed Development andMaturation in
Relevance to Seed Quality
• Since protection mechanisms are mainly built during the late seed
maturation phase, the stage of harvest becomes the most critical
factor for seed quality and storability.
• Harvesting seeds too early when there is inadequate development of
essential structures and protection mechanisms may result in poor
quality.
• Similarly, harvesting too late may increase the risk of shattering and
may decrease the quality of seed due to ageing.
• If harvesting is delayed, incidence of adverse environmental conditions
such as rain and humidity may result in precocious germination
59.
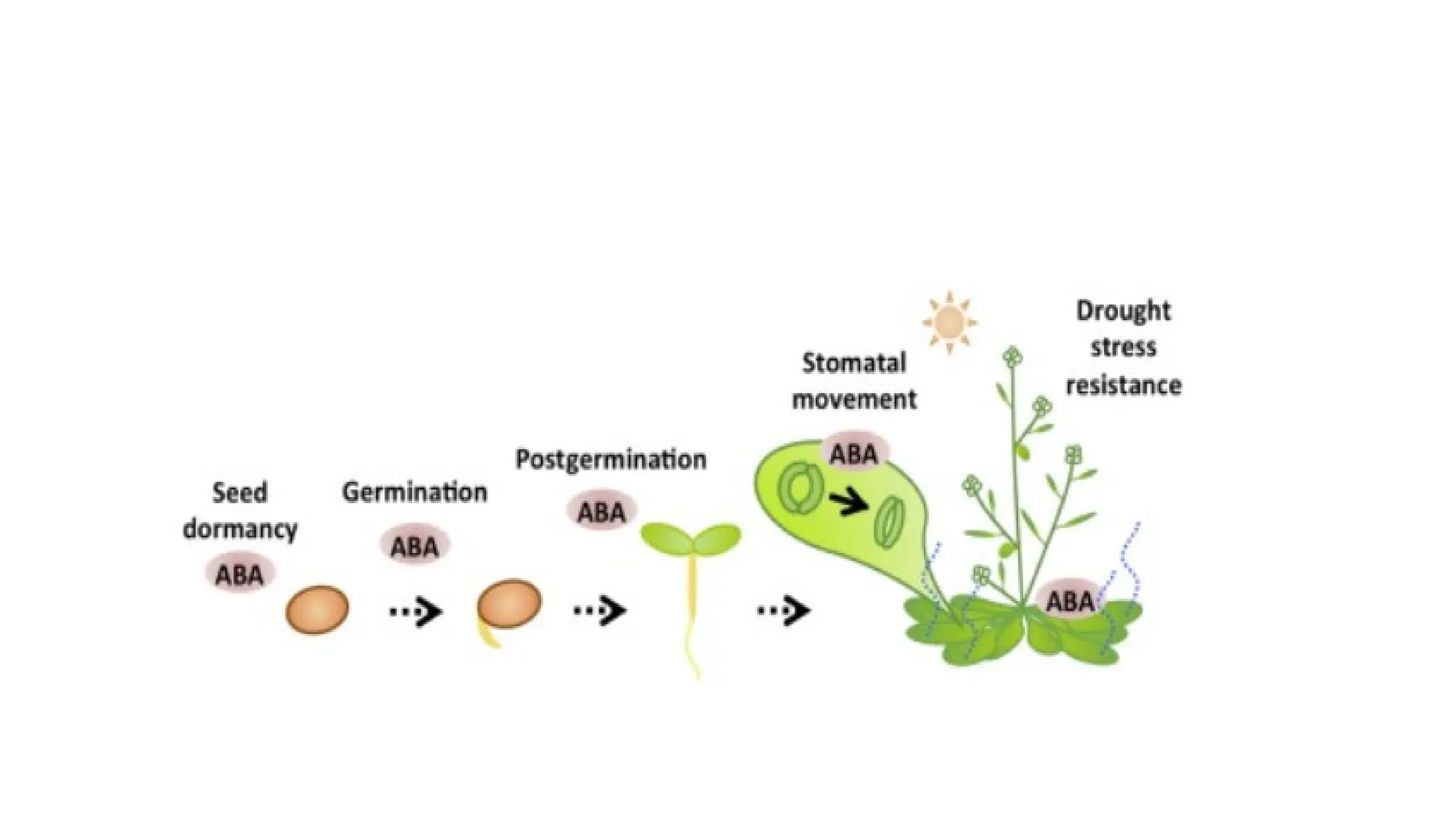
Function of AbscisicAcid
• Plants have openings on the bottom side of their leaves, known as
stomata. Stomata take in carbon dioxide and regulate water content.
Abscisic acid has been found to function in the closing of these
stomata during times when the plant does not require as much
carbon dioxide or during times of drought when the plant cannot
afford to lose much water through transpiration.
• One of the crucial functions of abscisic acid is to inhibit seed
germination.
• ABA functions in many plant developmental processes, including seed
and bud dormancy, the control of organ size and stomatal closure.
It is especially important for plants in the response to environmental
stresses, including drought, soil salinity, cold tolerance, freezing
tolerance, heat stress and heavy metal ion tolerance.
60.
• Abscisic acid(ABA) is one of the most important
phytohormones that influence seed development and
germination.
• At the molecular level, ABA biosynthesis, degradation, and
signaling genes were identified to play important roles in
seed development and germination
• Additionally, the crosstalk between ABA and other
hormones such as gibberellins (GA), ethylene (ET),
Brassinolide (BR), and auxin also play critical roles
61.
• Functional ABAbiosynthesis genes show specific roles for ABA accumulation
at different stages of seed development and seedling establishment.
• De novo ABA biosynthesis during embryogenesis is required for late seed
development, maturation, and induction of primary dormancy.
• ABA plays multiple roles with the key LAFL hub to regulate various
downstream signaling genes in seed and seedling development.
• Key ABA signaling genes ABI3, ABI4, and ABI5 play important multiple
functions with various cofactors during seed development such as degreening,
desiccation tolerance, maturation, dormancy, and seed vigor.
• The crosstalk between ABA and other phytohormones are complicated and
important for seed development and seedling establishment.
65.
• There aretwo important phases of seed development which include
zygotic embryogenesis, seed maturation
66.
• Physiological maturityis marked as the time when seeds attain
maximum dry weight and thereby, maximum yield when it concerns
crop production.
Editor's Notes
#2 Morpho –shape, size colour and arrangement physical-weght n height physiological biological systems operation respiration, metabolism biochemical; biosynthesis, enzyme activity hormonal regulation
#4 Endosperm nucleus undergoes DNA replication (endoreplication) here there is no cytokinesis , so nuclear DNA Increases within single cytoplasm.As endoreduplication continues, multiple nucli form within same cytoplasmic compartment multinucleate endopserm,. Eventully cellularization occurs.
#5 seed development begins with fertilization of both egg cell and central cell to form the diploid embryo and triploid endosperm. Cytokinesis cytoplasm divide into two daughter cells and divide into two nucli during mitosis..
#9 Dry weight : weight of plant once water us removed structural components, cellulose, proteins and lipids so remains stable
#10 Dessication tolerance: ability to withstand extree dehydration; quiscient: stem cells wait for differentiation until signalling occurs)
#13 Structura integrity :cell wall composition, turgor pressire, lignification, collenchyma and sclerenchyma for mechanical suppport






















































































![ipr[2].pptxfor agriculture students purpose](https://cdn.slidesharecdn.com/ss_thumbnails/ipr2-250817081026-ecef0da2-thumbnail.jpg?width=640&height=640&fit=bounds)



















