
Recommended
Recommended
More Related Content
What's hot
What's hot (19)
Similar to Heterotrimerric g protein can helpto feed the world
Similar to Heterotrimerric g protein can helpto feed the world (20)
More from Upasana Mohapatra
More from Upasana Mohapatra (8)
Recently uploaded
Recently uploaded (20)
Heterotrimerric g protein can helpto feed the world
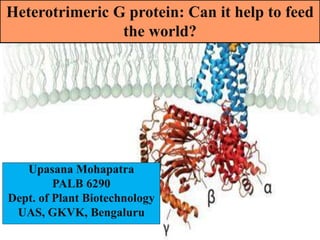
- 1. Heterotrimeric G protein: Can it help to feed the world? Upasana Mohapatra PALB 6290 Dept. of Plant Biotechnology UAS, GKVK, Bengaluru
- 2. CONTENTS Summary and future prospectives Case study G protein signaling in plant stress responses G protein signaling in plant physiological functions G protein subunits Structure and activation of G protein History April 13,2018 2Dept. of Plant Biotechnology,UAS(B)
- 3. HISTORY OF PLANT G-PROTEIN SCIENCE Urano et al., 2013 April 13,2018 3Dept. of Plant Biotechnology,UAS(B)
- 4. Structure of G protein • Heterotrimeric G protein signaling regulates a wide range of growth and developmental processes in animals and plants. • The membrane bound heterotrimeric G protein complex is composed of three subunits: alpha (Gα), beta (Gβ), and Gamma (Gγ) , G-protein-coupled receptors (GPCRs), and regulators of G-protein signaling proteins (RGSs). April 13,2018 4Dept. of Plant Biotechnology,UAS(B) ( Liu et al.,2017).
- 5. Activation of G protein • Activation is mediated by binding of an extracellular ligand to a 7-pass trans- membrane (7TM) G protein- coupled receptor (GPCR). • GPCRs are guanine-nucleotide exchange factors; they cause Gα to exchange GDP for GTP, leading to the dissociation of the complex into Gα -GTP and Gβγ dimers. April 13,2018 5Dept. of Plant Biotechnology,UAS(B) ( Liu et al.,2017).
- 6. The ‘G’ cycle of animals versus Arabidopsis Urano et al., 2013 April 13,2018 6Dept. of Plant Biotechnology,UAS(B)
- 7. Crystal structure and activation mechanisms of G protein Urano et al., 2013 April 13,2018 7Dept. of Plant Biotechnology,UAS(B)
- 8. Domain structures of Arabidopsis G protein-related proteins Urano et al., 2013 April 13,2018 8Dept. of Plant Biotechnology,UAS(B)
- 9. Domain structures of Arabidopsis G protein-related proteins Urano et al., 2013April 13,2018 9Dept. of Plant Biotechnology,UAS(B)
- 10. Structural characteristics of G protein signaling related proteins Stateczny et al., 2016April 13,2018 10Dept. of Plant Biotechnology,UAS(B)
- 11. Structural characteristics of G protein signaling related proteins Stateczny et al., 2016 April 13,2018 11Dept. of Plant Biotechnology,UAS(B)
- 12. Model for an intracellular kinase-dependent G protein cycle in Arabidopsis Stateczny et al., 2016 April 13,2018 12Dept. of Plant Biotechnology,UAS(B)
- 13. Hypothetic model for a membrane associated kinase-dependent G protein mechanism in Arabidopsis: Stateczny et al., 2016 April 13,2018 13Dept. of Plant Biotechnology,UAS(B)
- 14. G protein subunits In Plant cell • In Arabidopsis- one canonical Gα (AtGPA1) one Gβ (AGβ1) three Gγ(AGγ 1, AGγ 2,AGγ3) • In rice-one canonical Gα , one Gβ and 5 Gγ –subunits • γ -subunits are four times the average mammalian size. • Many plant γ -subunits do not contain an isoprenylation motif at their C-terminus. In Human beings • 23 Gα, five Gβ, and 12 Gγ subunits • γ -subunits are very small proteins (less than 100 amino acids) • An obligate requisite in all animal γ -subunits and essential for membrane anchoring. April 13,2018 14Dept. of Plant Biotechnology,UAS(B) Stateczny et al., 2016
- 15. Physiological Functions of G Protein Cell proliferation Root development Leaf and fruit development Chloroplast development Hormonal regulation of seed germination Auxin and sugar signalling ABA, Ca2+ and ROS signalling in guard cells. Stomatal opening and closure Phytochrome and cryptochrome mediated responses April 13,2018 15Dept. of Plant Biotechnology,UAS(B) Stateczny et al., 2016
- 16. 1. Arabidopsis T-DNA insertion lines for Gα (gpa1–4), Gβ (agb1–2) or RGS1 (rgs1–2) and wild-type Col-0 were grown for 37 days in a short day chamber (8 L : 16 D cycle, 100 µmol m-2 s-1) at 230C. 2. Nipponbare (wild-type), the Gα knockout (d1,) or Gβ knockdown lines of rice were grown in a short day chamber (8 L : 16 D cycle, 34 0C during day per 28 0C during night time, 320 µmol m-2 s-1) for 47 days. April 13,2018 16Dept. of Plant Biotechnology,UAS(B)
- 17. Physiological Function of G Protein April 13,2018 17Dept. of Plant Biotechnology,UAS(B)
- 18. April 13,2018 18Dept. of Plant Biotechnology,UAS(B)
- 19. The α subunit encoded by a single gene (GPA1) in Arabidopsis, is a modulator of plant cell proliferation. 1) gpa1 mutants have reduced cell division. 2) Inducible over-expression of GPA1 in Arabidopsis confers inducible ectopic cell division which causes premature advance of the nuclear cycle and the premature appearance of a division wall. April 13,2018 19Dept. of Plant Biotechnology,UAS(B)
- 20. •Cell production rates were determined for the RAM and lateral root formation in gpa1 and agb1 single and double mutants, and in transgenic lines over expressing GPA1 or AGB1. •In the RAM that the heterotrimeric complex acts as an attenuator of cell proliferation, whereas Gα-subunit’s role is a positive modulator. •For the formation of lateral roots, the Gβγ-dimer acts independently of the Gα -subunit to attenuate cell division. April 13,2018 20Dept. of Plant Biotechnology,UAS(B)
- 21. April 13,2018 21Dept. of Plant Biotechnology,UAS(B)
- 22. G Protein in Stress Response Oxidative stress and hypoxia responses Pathogen responses Drought stress Salinity stress April 13,2018 22Dept. of Plant Biotechnology,UAS(B)
- 23. 1. TheAGB1 mutants (agb1-2-1 and agb1-3-2) were more sensitive to drought than the wild-type. 2. The overexpression of mulberry (Morus alba L.) G-protein b subunit in transgenic tobacco (Nicotiana tabacum L.) significantly enhanced the plants' drought tolerance. 3. The transgenic tobacco plants had higher proline contents and peroxidase activities, and lower malonaldehyde and hydrogen peroxide contents and superoxide free radical accumulations under drought conditions and antioxidative genes, NtSOD and NtCAT, are increased in drought-stressed transgenic tobacco plants April 13,2018 23Dept. of Plant Biotechnology,UAS(B)
- 24. Overexpression of mulberry (Morus alba L.) G-protein β subunit in transgenic tobacco (Nicotiana tabacum L.) TheAGB1 mutants (agb1-2- 1 and agb1-3-2) were more sensitive April 13,2018 24Dept. of Plant Biotechnology,UAS(B)
- 25. •The knockout mutant of the Arabidopsis heterotrimeric G-protein Gβ subunit, agb1, is hypersensitive to salt, exhibiting a leaf bleaching phenotype. •AGB1 is mainly involved in the ionic toxicity component of salinity stress and plays roles in multiple processes of Na+ homeostasis. April 13,2018 25Dept. of Plant Biotechnology,UAS(B)
- 26. The knockout mutant of the Arabidopsis heterotrimeric G- protein Gβ subunit, agb1, is hypersensitive to salt, exhibiting a leaf bleaching phenotype. Three-week-old hydroponically grown plants treated with 100 mm NaCl for 10 d. Survival rate is less in agb-1 mutant plants. April 13,2018 26Dept. of Plant Biotechnology,UAS(B)
- 27. PAMP-triggered immunity Cell death and defence responses Signalling against bacterial pathogens Elicitor-induced stomata closure Resistance against necrotrophic fungi Non-host resistance R protein mediated disease resistance Heterotrimeric G proteins in plant defence mechanisms Nitta et al., 2015 April 13,2018 27Dept. of Plant Biotechnology,UAS(B) Gβ Gβ Gα Gβ,Gγ Gα,Gβ Gα,Gβ,Gγ Gβ,Gγ
- 28. Defence-related phenotypes observed in heterotrimeric G protein mutants Nitta et al,2015 April 13,2018 28Dept. of Plant Biotechnology,UAS(B)
- 29. The Gα subunit is not involved in the Gβ-mediated defense pathway. i) chorosis, and necrosis were less developed in agb1-2 ii) inflorescence growth was also inhibited in gpa1-4 plants, although to a lesser extent than in agb1-2; and iii) endogenous SA did not affect resistance to CMV and TuMV and did not interfere with AGB1- mediated responses. April 13,2018 29Dept. of Plant Biotechnology,UAS(B)
- 30. Khatri et al,2012 April 13,2018 30Dept. of Plant Biotechnology,UAS(B)
- 31. Case study 1. The Gβ protein is essential for plant survival and growth. 2. Gα provides a foundation for grain size expansion. 3. Three Gγ proteins, DEP1, GGC2 and GS3, antagonistically regulate grain size. DEP1 and GGC2, individually or in combination, increase grain length when in complex with Gβ. April 13,2018 31Dept. of Plant Biotechnology,UAS(B)
- 32. Grain size is a trait for grain quality focused by rice breeders, as long and slender grains are preferred by rice consumers in many countries Recent advances in rice functional genomics facilitated the cloning of a series of loci controlling grain size, including genes 1) Grain length -GS3, GL3. An-1, GLW7, GS2 genes 2) Grain width -GW2, GW5, GS5, GW8 , GW7 3) Grain weight-GIF1, GE, TGW6, GW6a, BG1,XIAO Introduction April 13,2018 32Dept. of Plant Biotechnology,UAS(B)
- 33. Types of Gγ Type A • Small in size • Has C-terminal CaaX isoprenylation motif • eg-RGG1- regulation of abiotic stress Type B • Small in size • Lack CaaX motif • eg-RGG2- regulation of abiotic stress Type C • 2 well defined region- • 1)an N-terminal domain with high similarity to g subunit • 2) A C terminal cystein rich subunit • eg- DEP1,GS3- QTL for grain size. April 13,2018 33Dept. of Plant Biotechnology,UAS(B)
- 34. Methods 1 • Field planting and trait measurement. • Vector construction and plant transformation -Constructs of GS3-1OE, GS3-4OE, DEP1OE, dep1OE and GGC2OE , under an ubiquitin gene promoter. 2 • Generation of knock-out mutants • Single mutant, double mutant and triple mutant of GS3,DEP1, GGC2 3 • RNA extraction and qRT-PCR. • Western blot. 4 • Statistical analysis-The two-tailed t test was used for comparing agronomic traits of each transgenic line with the control April 13,2018 34Dept. of Plant Biotechnology,UAS(B)
- 35. •Elevated DEP1 accumulation increased the grain length by 6.85–9.58% with a normal plant stature. •Plants over expressing dep1 showed very similar phenotype to DEP1Ri plants, both of which reduced grain length by ~4.5%, together with dwarf stature and erect panicles. •The expression level of endogenous DEP1 in dep1OE was not reduced. • Thus dep1 showed a dominant-negative effect over DEP1 in regulating grain size rather than co-suppression of the two genes.April 13,2018 35Dept. of Plant Biotechnology,UAS(B)
- 36. •GS3-1OE showed an average 9.07% reduction in grain length together with reduced plant stature. •GS3-4OE produced even smaller plants and grain size with an average 19.10% reduction of grain length . •GS3-1Ri resulted in an average 5.78% increase of grain length. April 13,2018 36Dept. of Plant Biotechnology,UAS(B)
- 37. April 13,2018 Dept. of Plant Biotechnology,UAS(B) 37 GS3-1Ri X dep1OE F1 Reduced grain length, similar to dep1OE transgenic plants. F2 Also showed the dep1OE phenotype with respect to grain size, plant height and panicle length Interaction between DEP1 and GS3 in grain size regulation
- 38. GS3-1ko mutant-increased grain size, DEP1ko mutant - reduced grain size. GS3-1koDEP1ko double mutant- intermediate between that of the single mutants of GS3-1ko and DEP1ko GS3-1koDEP1ko double mutant X ZH11 F2 Increased grain length was observed when over expressing DEP1 in GS3-1Ri background, whereas the DEP1OE/GS3-1OE hybrid showed the GS3-1OE phenotype of short grain April 13,2018 38Dept. of Plant Biotechnology,UAS(B)
- 39. An atypical Gγ subunit GGC2 functions additively with DEP1 Over expression of GGC2 (GGC2OE) in ZH11 increased grain length significantly, knock-out mutant of GGC2ko in ZH11 reduced the grain length. Knocking out DEP1ko GGC2ko resulted in much smaller grains than either of the single knock-out mutants, DEP1 and GGC2 worked additively in positive regulation of grain length. The DEP1koGGC2koGS3-1ko triple mutant produced similar phenotype to that of DEP1koGGC2ko, thus GS3-1ko mutant could not increase grain length when both DEP1 and GGC2 were knocked-out DEP1 and GGC2 functioned positively in regulating grain size in an additive manner, while the role of GS3 in grain size regulation was to repress the effects of DEP1 and GGC2 on increasing grain size. April 13,2018 39Dept. of Plant Biotechnology,UAS(B)
- 40. Functional dependence of the Gγ subunits on RGB1 and RGA1. The transgenic plants with suppressed expression of RGB1 (Knocked down RGB1 by RNAi (RGB1Ri) )showed reduced grain size and plant height. GS3-1Ri X RGB1Ri DEP1OE X RGB1Ri Reduced grain length Reduced grain length Overexpressed GGC2 in RGB1Ri. Overexpressed DEP1 in RGB1Ri. Reduced grain length The effects of grain length increase by DEP1OE, GGC2OE and GS3-1Ri are dependent on RGB1 and RGA1 April 13,2018 40Dept. of Plant Biotechnology,UAS(B)
- 41. Schematic representation of the functions of the G proteins in grain size regulation April 13,2018 41Dept. of Plant Biotechnology,UAS(B)
- 42. Conclusion • DEP1 and GGC2, when coupled with RGB1, promote grain size by tail-mediated signaling. • GS3,reduces grain size by blocking the interaction of DEP1 and GGC2 with RGB1. • The increased grain size is at the cost of fitness. Thus from fitness viewpoint medium grain is the optimal for rice reproduction. • Thus, while DEP1 and GGC2 function to promote grain size,which is essential for yield increase in breeding, GS3 plays a role to keep balance for maintaining fitness in response to natural selection. April 13,2018 42Dept. of Plant Biotechnology,UAS(B)
- 43. 1) The interactions of three Gγ proteins can be used for a predictable design of grain size in rice, by manipulating these genes, individually or in combination, to improve rice grain yield and quality. 2) As the Gγ proteins are highly conserved in a very wide range of plants, manipulating these proteins may provide a general strategy for modifying organ size and yield in crop breeding. 3) G signalling is at the heart of many plant physiologies of agronomic importance, such as disease resistance, abiotic stress tolerance and harvest index. Translational work on G protein signalling will certainly improve agriculture by providing new targets and strategies for increasing yield. April 13,2018 43Dept. of Plant Biotechnology,UAS(B)
- 44. Future Prospectives • What are the receptors upstream of G-proteins, what lies downstream of G-proteins, and how the proteins connect to the established modules of hormone, defense or stress-related signaling. • Although the G proteins have been shown to have key roles in regulating plant growth, stress tolerance and grain yield potential, the molecular mechanisms underlying the regulation of the heterotrimeric G protein cycles of activation and deactivation and downstream effectors still remain unclear. April 13,2018 44Dept. of Plant Biotechnology,UAS(B)
- 45. April 13,2018 Dept. of Plant Biotechnology,UAS(B) 45 THANK YOU Science of G protein is awaiting your research efforts, to be explored…….
- 46. REFERENCES BOTELLA, J.R., 2012, Can heterotrimeric G proteins help to feed the world?. Trends in plant science, 17(10) : 563-568. CHEN, J.G., GAO, Y. AND JONES, A.M., 2006, Differential roles of Arabidopsis heterotrimeric G-protein subunits in modulating cell division in roots. Plant Physiology, 141(3), pp.887-897. LIU, C., XU, Y., LONG, D., CAO, B., HOU, J., XIANG, Z. AND ZHAO, A., 2017, Plant G-protein β subunits positively regulate drought tolerance by elevating detoxification of ROS. Biochemical and biophysical research communications, 491(4): 897-902. STATECZNY, D., OPPENHEIMER, J. AND BOMMERT, P., 2016, G protein signaling in plants: minus times minus equals plus .Current opinion in plant biology, 34 :127-135. SUN, S., WANG, L., MAO, H., SHAO, L., LI, X., XIAO, J., OUYANG, Y. AND ZHANG, Q., 2018, A G-protein pathway determines grain size in rice. Nature communications, 9(1): 851. URANO, D., CHEN, J.G., BOTELLA, J.R. AND JONES, A.M., 2013, Heterotrimeric G protein signalling in the plant kingdom. Open biology, 3(3): 120186. XU, Q., ZHAO, M., WU, K., FU, X. AND LIU, Q., 2016, Emerging insights into heterotrimeric G protein signaling in plants. Journal of Genetics and Genomics, 43(8): 495-502. April 13,2018 46Dept. of Plant Biotechnology,UAS(B)
Editor's Notes
- History of plant G protein science. In the 1970s, G proteins were identified as a signal transducer connecting the hormone receptor and the adenylyl cyclase in mammals. In the early 1990s, plant G protein genes were cloned and shown to have conserved domains and motifs with the animal genes. In the late 1990s, much effort went towards physiological roles of G proteins using genetics. In the 2000s, the Gβγ-subunits, the regulators (GPCR-like genes and a 7TM-RGS gene) and effectors of G protein were cloned and characterized genetically and biochemically. In 2007, the ‘self-activating’ property of the plant G protein was revealed. In addition, the transcriptome, proteome and interactome analyses revealed comprehensive knowledge of the plant G protein pathways. In the last few years, the crystal structure and computational simulation solved the mechanism of self-activation. Publications on the physiological functions and signalling components of G protein pathways are exponentially increasing, providing evidence for their important and divergent functions in plants.
- These two functional subunits activate different signaling cascades until the intrinsic GTPase activity of the Gα subun
- RGS-Regulator of G proteins signaling The ‘G’ cycle of animals versus Arabidopsis. (a) G protein regulation in mammalian cells. In the absence of liGα nd, G protein forms an inactive heterotrimer with Gβγ dimer (bottom left). LiGα nd-bound GPCR promotes GDP dissociation and GTP binding on G protein (top). GTP-bound Gα dissociates from Gβγ dimer, and both activated Gα and freely released Gβγ modulate activity of the effectors (bottom right). Gα hydrolyses GTP to GDP, and re-binds to Gβγ to return to its inactive state. (b) G protein regulation modelled in Arabidopsis. Arabidopsis G protein (AtGPA1) can spontaneously dissociate GDP and activate itself (bottom left). AtGPA1 does not hydrolyse its GDP rapidly; however, AtRGS1, a 7TM-RGS protein, promotes the GTP hydrolysis of AtGPA1 (top). d-glucose or other stimuli functions on AtRGS1 directly or indirectly, and decouples AtGPA1 from AtRGS1 (bottom right). Once released from AtRGS1, AtGPA1 does not hydrolyse its GTP efficiently, maintaining its active state and modulating the effector activities.
- Crystal structure and activation mechanisms of G protein. (a) Structural basis of animal G protein activation. Left: Gα protein forms stable heterotrimer with Gβg dimer (grey and black) at the steady state. GDP (green) is tightly bound to a Ras domain (orange) of the a-subunit, and covered by the helical domain (sky blue). Right: in the presence of liGα nd-bound receptor, the helical domain moves and changes orientation. The structural change causes GDP dissociation from the a-subunit, the subsequent GTP binding and activation. (b) Structure of Arabidopsis AtGPA1 is entirely similar to mammalian Gα proteins. However, the helical domain of AtGPA1 fluctuates spontaneously. The spontaneous fluctuation initiates GDP dissociation, and nucleotide exchange. Crystal structures shown are animal heterotrimeric G protein (PDB: 1GOT) [56], G protein and b2 adrenergic receptor (PDB: 3SN6) [6], and Arabidopsis AtGPA1 (PDB: 2XTZ) [41]. The cartoon for the animal model was adapted from Rasmussen et al. [6].
- 1) AtRGS1 keeps the heterotrimeric complex in its inactive state by preventing the exchange of GDP to GTP. (2) D-Glucose induction promotes the phosphorylation of AtRGS1 by WNK8 (intracellular kinase) (3) Phosphorylated AtRGS1 undergoes endocytosis and the uncoupled Gα protein is capable to exchange GDP for GTP. GTP bound Gα and the detached Gβγ dimer can activate downstream effectors.
- (1) AtRGS1 keeps the heterotrimeric complex in its inactive state by preventing the exchange of GDP to GTP. (2) Upon binding of a liGα nd, the RLK dimer phosphorylates AtRGS1. (3) Phosphorylated AtRGS1 undergoes endocytosis and the uncoupled Gα protein is capable to exchange GDP for GTP. GTP bound Gα and the detached Gβγ dimer can activate downstream effectors.
- G protein g-subunits exhibit an extraordinary level of structural diversity
- These results suggest that Arabidopsis heterotrimeric G-protein subunits have differential and opposing roles in the modulation of cell division in roots
- These results suggest that Arabidopsis heterotrimeric G-protein subunits have differential and opposing roles in the modulation of cell division in roots
- Cell death is regulated by AGB1 ..as agb1 is not there there will be stoppage of cell growth and no necrosis and chlorosis.so the defence is regulated by Gb. SA is not involved inhis regulation pathway