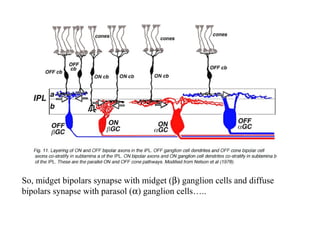
1. So, midget bipolars synapse with midget (b) ganglion cells and diffuse
bipolars synapse with parasol (a) ganglion cells…..
2. Ganglion Cell Projections to the
LGN
• It was already shown that the receptive fields
of parasol ganglion cells are considerable
larger than that of midget ganglion cells at the
same retinal eccentricity.
3. • It is argued that the smaller receptive fields of
the midget cells provide greater spatial
resolution than the parasol cells, which
integrate energy across a wider range of
photoreceptors (via horizontal, bipolar, and
amacrine cells).
– On the other hand, the sensitivity of the parasol
cells would be superior.
• They also differ in their temporal responses.
– Midget (parvo) cells responding with sustained
spike trains as long as a light source is projected
onto the excitatory portions of its receptive fields.
– Parasol (magno) cells respond at the onset, but
firing rate quickly goes down the spontaneous rate
(transient firing).
4. • The projection of visual space onto
the retina is such that information
about objects in the left visual field
is projected to the right
hemisphere and information from
the right field is projected to the left
hemisphere.
• The nasal portions of the retina
cross, while the temporal portions
project ipsilaterally.
• In evolution, the decussating
(crossing) path is the oldest.
– As the eyes moved medially, the
ipsilateral pathway developed.
– This results in greater visual field
overlap (C-D), and the ipsilateral
pathway assures that the inputs
from the overlapping fields go to
both cortices.
Input (letters A-F) from the right visual field are mapped in an orderly fashion
to the left LGN, while the left visual field projects to the right LGN.
The top 4 layers form the parvocellular layers and receive input from midget
ganglion cells.
The bottom 2 layers from the magnocellular layers and receive input from
parasol ganglion cells.
5. • If one measures the conduction
times for electrical signals traveling
from the retina to the parvocellular
layers of the LGN, they are longer
than the latencies to the
magnocellular layer (on the
average).
o Schiller and Malpeli (1978)
applied an electrical stimulus at
the optic chiasm and measured
the time it took the signal to
travel to the various layers.
• The receptive fields of LGN cells
are not appreciably different from
those of ganglion cells, but LGN
cells are influenced by descending
input from the cortex (visual and
other areas), the brainstem, from
other cells in the thalamus, from
other LGN cells
• The descending input from cortex
to the LGN is actually more
substantial the that projections from
the LGN to the cortex!
6. • The fact that the LGN contains a
retinotopic map can be seen in
oblique, electrode tracks.
• This is significant because it
demonstrates that neurons entering
the LGN are arranged so that fibers
carrying signals from the same area
of the retina end up the same area of
the LGN, and neighboring retinal
regions project to neighboring LGN
regions.
• Retinotopic maps occur in each of
the layers, and the maps line up
with each other, as seen in
perpendicular electrode penetrations
(all neurons would have receptive
fields at the same locations).
B A
CC’ B’A’
C
7. • The corpus callosum, which consists of fiber
tracts between the two hemispheres,
integrates the left and right visual fields so
well that we do not notice that they are
encoded independently.
• The geniculostriate pathway (LGN to cortex)
is clearly the most important and most
recently evolved.
• About 90% of the optic nerve fibers go to
the lateral geniculate body.
• The other 10% go to the superior colliculus,
consisting of collaterals from the
geniculostriate pathway and possibly a few
direct fibers projecting from the optic nerves.
• In non-mammalian species (e.g., birds and
fish) superior colliculus is called the optic
tectum, and it serves the function of the
geniculostriate pathway (color, form).
8. • For mammals, the superior colliculus
appears to play a role in the
orientation of the animal in space.
• Snyder found that lesions of the
superior colliculi of hamsters
produced behavior that is consistent
with deficits in orientation but not
discrimination.
• Animals forced to discriminate
horizontal from vertical bars do
horribly if they must run down a left
or right alley, but perform well in
go/no go tasks.
o It is as though they can discriminate
vertical from horizontal but cannot
tell left from right (they do not get
reinforcement because they bump
into objects along the way).
o Note the importance of the task
performed by the animal, because
many early researchers concluded
that lesions of the superior colliculi
produced blindness while others
claimed no effect of the lesions.
9. Striate Cortex
• The very rear of the occipital lobe is
where the LGN projects.
• The area has several different
names: primary visual cortex, V1,
area 17, or striate cortex (because
of the striped pattern it takes on
after staining).
• It consists of 6 major layers, some
having sublayers.
Fibers from the LGN project mainly
into layer 4, with magnocellular
neurons (2 ventral LGN layers)
coming into layer 4Cα and
parvocellular neurons (4 dorsal
layers) coming into layer 4Cβ.
α
b
11. Recording From Units in V1
• The first recordings in Area 17 were made by
Jung in Germany in the mid 1950's from cats.
– At the time, little was known about the responses
of the earlier cells in the pathway, and the study
was a dismal failure.
– Jung presented flashes of light and concluded that
90 95% of the cells in the visual ‑ cortex simply did
not respond to light.
– This was most likely due to the size of his flashes,
which produced a balance of inhibition and
excitation from the center‑surround fields.
12. Recording From Units in V1
• All that changed in the late 1950’s with
the pioneering work of David Hubel and
Torsten Wiesel.
15. Recording From Units in V1
• David Hubel and Torsten Wiesel knew what types of information
were passed along from lower levels of the system, since
Torsten Wiesel had worked in Stephen Kuffler’s lab at Johns
Hopkins in 1955.
– Kuffler had carried out measurements of receptive fields of cat
ganglion cells, and this knowledge of center-surround antagonism
meant that Hubel and Wiesel stood a much better chance of asking
intelligent questions of the cortex.
– Because they knew of surround inhibition, they used patterned
stimuli that could maximize the probability of evoking responses.
– Their major contribution was that they found cells whose receptive
fields were elongated, orientationally specific, and more spatially
selective than LGN cells.
– Even with this knowledge, they still had difficulty getting cells to
respond to light.
• As they gained a better understanding of what sorts of
information were being processed, a greater percentage of cells
could be driven.
– In 1959 they claimed that 50% could be driven, but by 1962 the
percentage was around 90 (once they found the length specificity).
• They were awarded the Nobel Prize in Physiology or Medicine
in 1981.
16. • "Simple" cells were the first from which recordings were
made, with receptive fields consisting of discrete
inhibitory and excitatory regions.
• Some of these have bipartite fields and others have
tripartite fields.
• They had clearly defined excitatory and inhibitory
regions.
• About 80% of the simple cells are binocular, having
similar receptive fields for the two eyes.
17. • The elongation makes these cells orientation specific, with
the preferred orientation varies from cell to cell.
• One idea was to take the outputs of LGN cells an align
them in such a way to produce various elongated
receptive fields.
18. Complex Cells
1 2 3 4 5
• "Complex" cells do not have discrete excitatory and inhibitory
subregions.
– If their receptive fields are mapped with small spots of light, one
finds a mixture of small areas of excitation and inhibition, with only
very small responses.
– The optimal stimulus is a light or dark bar somewhere in the field
that must not cover too large of a region.
– Complex cells respond to the bar in any one of the subregions, but
the response diminishes as the bar covers more that one region at
a time; they all prefer moving bars.
– About 25% are directionally selective, preferring a moving stimulus
in one direction across the field (15 vs. 51).
– Like simple cells, complex cells are orientationally selective.
– As it turns out, approximately 75% of cortical neurons are classified
as complex.
• As such, it is hardly surprising that researchers had difficulty getting
them to respond to light, since most used stationary stimuli.
19. Hypercomplex" cells are like simple or complex cells, except that they are
end stopped on one or both sides to produce ‑ length specificity.
They are now thought to reflect subclasses of simple and complex cells.
Simple cells
20. Hierarchical Model
• Hubel and Wiesel believed that the
outputs of center-surround ganglion cells
projected to the LGN (remaining center-surround),
with multiple cells from the LGN
then projecting onto a single cortical
neuron.
• Multiple simple cell outputs could then
project onto a 2nd level cortical neuron,
producing complex cell receptive fields.
23. A Video Showing the Difference
between Simple and Complex Cells
www.youtube.com/watch?v=8VdFf3egwfg
24. The Hubel and Wiesel View of
Spatial Vision
• Because they demonstrated receptive fields that
were either bipartite (edge detectors) or tripartite
(line or bar detectors), their findings were
consistent with an atomistic approach.
– The argued that the fundamental building blocks of
objects were lines and edges at particular positions,
orientations, widths, lengths, contrasts, etc.
– Higher level shapes could be constructed by
assembling the receptive fields of simple, complex,
and hypercomplex cells found in V1.
25. Source of Inhibition?
• Note that all of the inhibition in the hierarchical model is
generated within the retina.
• Creutzfeld and his colleagues (1974) recorded
intracellularly from cortical cells (a monumental task)
while stimulating units in the LGN.
– In all cases the LGN input was excitatory, and the inhibition
observed had a longer latency (probably stemming from
cortical interactions).
• Sillito (1975, 1980) performed experiments in which
GABA antagonists (bicuculine) was applied
iontophoretically to the cortex in the vicinity of a
recording site.
– It eliminated both orientation and direction of movement
tuning, implying that they arise from interactions within cortex.
• Another criticism of the hierarchical model is the fact
that it is quite difficult to imagine how the response
properties of complex cells can be generated by
recombining simple cell outputs.
27. Striate Architecture
• Given that cells are “tuned” to different orientations, position,
sizes, colors, etc., the question arises as to how these features
are distributed across the cortex.
– This is a question of the architecture of the striate cortex—what is
the spatial layout and pattern of interconnections among cells tuned
to different values of these different stimulus dimensions?
– Answering this required Herculean recording sessions in which
researchers would find a cell and record from it until they
determined the optimal location, orientation, size, and eye
dominance.
– The microelectrode would then by moved a but further until another
cells was isolated.
• It’s “best features” would then be determined.
• This process was repeated until a great many cells were examined,
then the animal would be “sacrificed” and its brain examined
microscopically to determine the location of the electrode tracts.
– This endeavor was vastly progressed by the development of
autoradiographic techniques that rely on the uptake of radioactive
sugar into highly active cells (2-deoxyglucose studies), which
generally corroborated the single-unit recording data.
28. Retinotopic Map
• The layered sheets of cells that comprise
primary visual cortex within each hemisphere
are laid out in a retinotopic map of exactly half
the visual field.
• The map preserves retinal topography, with
nearby points on the retina projecting to nearby
cortical points.
• The metric properties of the map on the cortex
are distorted, however.
– The main distortion is due to cortical magnification of
central (foveal) areas relative to peripheral ones.
29. • Magnification is from the 2-deoxyglucose
study of Tootell et al. (1982).
30. • Tootel, Silverman, Switkes, and De Valois
(1982) • Since glucose is the metabolite of cortical
neurons, more is used by active cells.
• 2-deoxyglucose (2DG) is taken up by cells
as if it were glucose, but it remains in cells
(isn’t actually metabolized).
• Since it is radioactive, one determines
where it accumulates when a particular
stimulus is presented.
• A “rings and rays” pattern (A) centered on
the fovea was presented while the monkey
was injected with 2DG.
o The rings and rays display was
composed of small (randomly sized)
rectangles that flickered over time,
with the rings spaced logarithmically
(from the center).
31. • Tootel, Silverman, Switkes, and De Valois
(1982) •The cortical surface is flattened, then sliced
thinly parallel to the surface and placed on
X-ray sensitive film.
•The logarithmically spaced rings stimulate
strips in V1 that are about equally spaced
on the cortex, indicting that a small region
near the fovea activates a
disproportionately large area of cortex.
o Peripheral regions stimulate smaller
Foveal cortical regions.
On left Activation caused by hemicircles.
Activation caused by radii.
32. Ocular Dominance Slabs
• We have two eyes, and both project to both
hemispheres.
• This raises the question as to whether we have
separate retinotopic maps in the cortex or one
integrated one.
– The answer lies somewhere in between—there is one
global map for each cortex, within which cells that are
dominated by one eye or the other are interleaved.
– Ocular dominance varies from one eye being
dominant to both eyes being equally effective at
driving a cell.
33. In population studies of ocular dominance, Hubel and Wiesel studied
hundreds of cells and categorized each one as belonging to one of seven
arbitrary groups. A group 1 cell was defined as a cell influenced only by
the contralateral eye—the eye opposite to the hemisphere in which it sits.
A group 2 cell responds to both eyes but strongly prefers the
contralateral eye. And so on.
Clearly there are differences between cat and macaque….. Rhesus macaques show few
cells that are driven equally well by the two eyes while they are quite prevalent in cats.
34. Light is right Stimulus was a vertical line; eye, dark is left eye
spacing is about every 0.5 mm.
The figure at the left is an optical image
of superimposed orientation columns.
Again it is found that the full range of
orientations is represented every 0.5
mm.
35. The Hypercolumn
• These findings lead us to the concept of
the hypercolumn.
– Overall, V1 is composed of many smaller
cortical modules called hypercolumns.
– They are long and then running perpendicular
to the cortical surface through all 6 layers.
– Every 1 mm2 represents a full range of
orientations for right and left eye dominance.
36. Shown here are two adjacent
hyperocolumns, representing
adjacent point on the retina.
Every square mm represented
both occular dominances, with
orientations between 0 and 180o
represented twice
http://www.sinauer.com/wolfe2e/chap3/hypercolumnsF.htm
39. Adaptation
• The rationale of psychophysical adaptation
studies is that long term exposure to a given
stimulus fatigues channels responsive to it,
so that later perception is based on an altered
distribution of activity across channels tuned
to some dimension.
• This shift results in a change in the percept
experienced in the unadapted state.
• This allows psychophysical studies to
elucidate the presence of tuned channels.
• The following slides use orientation tuning as
an example….
40. • Let your gaze move back and forth over
the fixation dash, adapting the upper
half of your visual field to a tilt of -20o
and the lower half to +20o.
42. • Most subjects report that the vertical
lines in the upper half appear to be
tilted to the right, while the lower vertical
lines appear to be tilted to the left.
• What’s going on here?
43. In the unadapted state, Orientation X causes equal activity of channel
A and B. Say you adapt to Orientation W, reducing the
responsiveness of the A orientation channel. Orientation X would now
be perceived to have a greater orientation, since it is causing greater
activation of Channel B than Channel A.
A B
Orientation
Response
X
W
44. On the other hand, say you adapt to Orientation Y, reducing the
responsiveness of the B orientation channel. Orientation X would now
be perceived to have a smaller degree of tilt, since it is causing
greater activation of Channel A than Channel B.
A B
Orientation
Response
X
Y
45. Spatial Frequency Analysis
• No one doubts the contributions made by
Hubel and Wiesel, and the enormous leap
forward the visual science made on account
of their ability to “drive” visual cortical
neurons.
– At issue is the question of whether or not cells
truly prefer bars of different widths.
• I introduced the idea of a spatial modulation
transfer function as a measure of the ability of
humans to resolve spatial frequency.
– Threshold contrast was measured as a function of
the spatial frequency of sinusoidal gratings,
yielding functions like this:
46. • The spatial MTF shows best sensitivity to a
mid range of spatial frequencies (5-7 cycles
per degree), with sensitivity to higher and
lower spatial frequencies being somewhat
lower.
47. • This can be easily
explained on the
basis of center-surround
receptive
fields found at the
bipolar cell,
ganglion cell, and
LGN levels.
• Low spatial
frequencies excite
both center and
surround uniformly,
as do high spatial
frequencies.
• Intermediate spatial
frequencies excite
the center but not
the surround (or
vice versa).
48. • It was Campbell and Robson (1969) who had the audacity to
propose that the overall spatial MTF was based on the
“envelope” of tuned spatial frequency channels, shown in the
right panel.
– Essentially the visual system would consist of multiple spatial
frequency-tuned channels, and we would know the form of the
stimulus by knowing what spatial frequencies were present.
• At the heart of spatial frequency theory is the notion that all
complex distributions of luminance fluctuations across space
can be recreated by adding spatial sinusoids of known spatial
frequency, amplitude (contrast), orientation, and phase.
• It seems strange to consider spatial frequencies as the
“primitives” or atoms of visual perception because we do not
consciously experience their presence with analyzing complex
scenes.
49. • Odd integer harmonics are added
together at an amplitude that is harmonic
number….
50. • The idea is that we would perceive a
square wave because spatial frequency
tuned channels at f, 3f, 5f, 7f, etc would
be active, each less active that the one
preceding it since there is less power in
higher harmonics.
51. Back to Campbell and
Robson…
• If one adapts to a 7 c/deg grating, sensitivity is only lost near 7 c/deg.
• Sensitivity is only lost near the adapting spatial frequency, as though
the channel were fatigued by the adapting stimulus.
• The middle panel shows the difference between the unadapted and
adapted MTF, and can be thought of as inferring the shape of a spatial
frequency channel.
• But does it mean that spatial frequency per se is the variable encoded
by the visual system rather than bar width?
– Unfortunately, the visual system could be encoding the sinusoidal grading
as a blurry bar of a particular width, so one could interpret these findings as
demonstrating the loss of sensitivity to bars of particular widths.
52. • So what if one adapted to a square
wave?
– If the visual system were tuned to bar
widths, then this adapting stimulus should
cause reductions in sensitivity at the spatial
frequency corresponding to the bar width,
but not at other spatial frequencies.
– If, on the other hand, the extracted
dimension were spatial frequency per se,
then sensitivity should be lost at the odd
harmonics.
53. 3 9
Spatial Freq. (c/deg)
• There is loss at the fundamental (3 c/deg) and the 3rd harmonic
(9 c/deg)!
– Unless the fundamental frequency is very low, there is no real
opportunity to observe the loss in sensitivity at the 3F because (a)
sensitivity falls off so abruptly with spatial frequency and (b) there is
likely inhibition between adjacent spatial frequency channels.
• The inhibition between channels means that 1F and 3F and 3F and 5F
are likely to reduce the effectiveness of each other.
• Since the power in the stimulus goes down by the harmonic number, 1F
will squash the activation level of 3F and 3F with squash the activation
level of 5F IN THE VISUAL SYSTEM!
54. • Graham and Nachmias (1971)
found that the threshold for
detecting a compound of f+3f
could be predicted from the
magnitudes of the individual
components regardless of whether
they are added in “peaks add” or
“peaks subtract” phase.
• If the system computed the
contrast of the pattern, sensitivity
to “peaks add” stimuli would have
been much better than to the
“peaks subtract” stimuli because
of the manner in which contrast is
computed.
• Graham and Nachmias (1971)
found that the threshold for
detecting a compound of f+3f
could be predicted from the
magnitudes of the individual
components regardless of
whether they are added in
“peaks add” or “peaks subtract”
phase.
• If the system computed the
contrast of the pattern, sensitivity
to “peaks add” stimuli would
have been much better than to
the “peaks subtract” stimuli
because of the manner in which
contrast is computed.
C o n t r a s t
L L
L L
=
-
+
m a x m i n
m a x m i n
55. Adapt to the following gratings, ala
Blakemore and Sutton (1969)
56.
57.
58. In the un-adapted state, Spatial Frequency X causes equal activity of
channel A and B. causes equal activation of the short and long
channels. Say you adapt to Spatial Frequency W, reducing the
responsiveness of the B channel. Spatial Frequency X would now be
perceived to have a lower spatial frequency, since it is causing
greater activation of Channel A than Channel B (adapting to a higher
spatial frequency shifts the appearance to lower spatial frequencies).
A B C
Spatial Frequency
Response
X
W
59. Adapting to lower spatial frequencies makes higher spatial
frequencies look even higher, since the C channel is now much more
active than channel B.
A B C
Spatial Frequency
Response
X
W
60. Cortical Recordings
• Recordings from cortical cells are often interpreted now in terms of the range
of spatial frequencies to which the cells respond rather than in terms of the
bar widths to which they are sensitive.
• If gratings are used, cortical cells seem to be rather narrowly tuned, with
bandwidths of about 1.5 octaves (log base 2 of bandwidth) at points at which
sensitivity has fallen by a factor of 2 (relative to the peak).
– This means that the ratio of the higher to lower spatial frequencies at the half-sensitivity
points is 21.5 or 2.8 on the average.
• The distribution of bandwidths is quite large, with the monkey's foveal cortex
containing as many cell with bandwidths of 2.5 octaves as there are cells
with bandwidths of 0.7 octaves.
– In general, about a third of the cortical cells have bandwidths between 0.5 and 1.2
octaves, while a small sample are tuned like LGN cells.
– By comparison, the bandwidths of cells in the LGN (X-cells) are 3-4 octaves in the
cat and may exceed 5 octaves in the monkey, so the narrower cortical bandwidths
must be due to intracortical interactions.
• In general cortical cells have bandwidths that increase logarithmically with
peak spatial frequency, so the "octave" measure of tuning stays roughly
constant with peak spatial frequency (it declines slightly with increasing peak
spatial frequency).
• Differences in peak frequency are slight for simple and complex cells--
complex cells tend to be tuned to slightly higher spatial frequencies.
• Larger receptive fields (and low peak SFs) are generally found to emanate
from parafoveal regions, and there are fewer high-spatial frequency tuned
cells in extrafoveal cortical regions.
61. It is critical for the theory that any point in space be analyzed by elements
tuned to different spatial frequencies, so the previous statement reflects
general trends when one measures best spatial frequency as a function of
retinal eccentricity.
62. Local Spatial Frequency
Analysis
Since receptive fields of cortical neurons
is restricted, we believe that the system
carries out a local spatial frequency
analysis (no cell “sees” the entire visual
field).
The elements are modeled as Gabor
functions (Gaussian multiplied sine
waves).
63. • The figure below shows the relative contributions of
high and low spatial frequency information.
– (a) shows a complete face, (b) presents the same face
with only high spatial frequency components and (c)
shows the same face with only low spatial frequency
components.
– Low frequencies convey information about general shape
and form, while high frequency information provides the
detail.