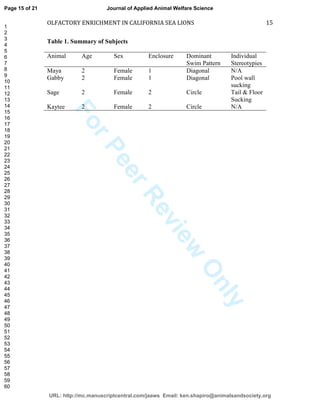
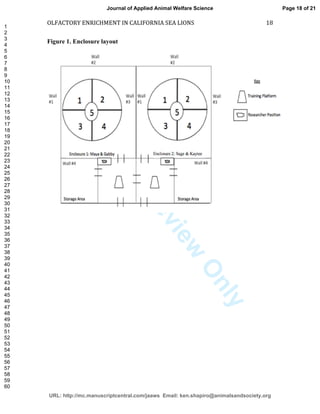
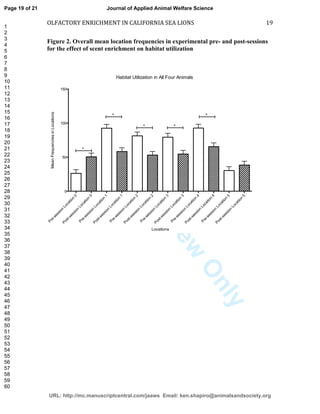
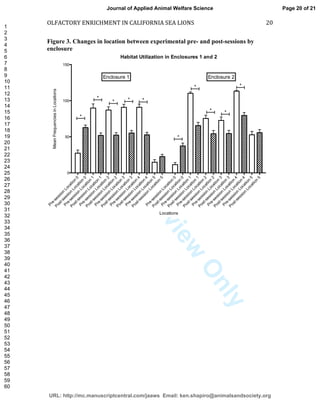
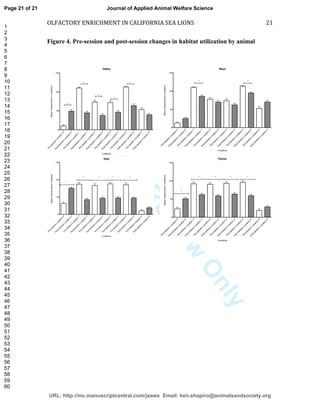
This study investigated the effects of olfactory enrichment on the behavior of captive California sea lions. Researchers introduced natural and non-natural scents and recorded the sea lions' behavior before and after. They found that scent enrichment significantly increased habitat usage, reducing repetitive swimming behaviors. Both natural and non-natural scents had this effect, with sea lions spending more time out of the water and less time swimming in repetitive patterns. This suggests that olfactory enrichment can improve welfare for captive sea lions by encouraging more natural behaviors.