Otolith Shape Variations Among Antarctic Fish Species
•Download as PPSX, PDF•
0 likes•1,295 views

The changes of the otolith shape with the growth of the swimming speed and changes in the environment
![Antarctic Circumpolar
WHITE-BLOODED: high Antarctic; ice pack zone; temperate
80°S 74, ~70°S; 63, ~60°S; 63, ~57°30’S; 52, ~45°S 30°S
S. Orkney SouthGeorgia
-100
-200
-300
-400
-500
-600
-700
-800
Current
[m]
S. japonicus
Channichthyidae
Ch. aceratus
Ps. georgianus
C. gunnari
Terrestrial observations of separate geographical and vertical living on different age groups and
species of fish suggest that differences in otolith shape among them became from difference in their
environment conditions. (Extracted and enlarged otoliths are over or near the fish heads: Median or Transverse plane)
21 km/h
1 km/h
0.9 km/h
0.1 km/h](data:image/gif;base64,R0lGODlhAQABAIAAAAAAAP///yH5BAEAAAAALAAAAAABAAEAAAIBRAA7)
Recommended
More Related Content
More from ryszardtraczyk
More from ryszardtraczyk (14)
Recently uploaded
Recently uploaded (20)
Otolith Shape Variations Among Antarctic Fish Species
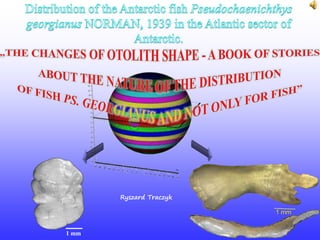
- 1. 1 mm 1 mm by Ryszard Traczyk 21.3 km/h ~0.9 km/h ~0.8 km/h
- 2. Antarctic Circumpolar WHITE-BLOODED: high Antarctic; ice pack zone; temperate 80°S 74, ~70°S; 63, ~60°S; 63, ~57°30’S; 52, ~45°S 30°S S. Orkney SouthGeorgia -100 -200 -300 -400 -500 -600 -700 -800 Current [m] S. japonicus Channichthyidae Ch. aceratus Ps. georgianus C. gunnari Terrestrial observations of separate geographical and vertical living on different age groups and species of fish suggest that differences in otolith shape among them became from difference in their environment conditions. (Extracted and enlarged otoliths are over or near the fish heads: Median or Transverse plane) 21 km/h 1 km/h 0.9 km/h 0.1 km/h
- 3. Antarctic Circumpolar WHITE-BLOODED: high Antarctic; ice pack zone; temperate 80°S 74, ~70°S; 63, ~60°S; 63, ~57°30’S; 52, ~45°S 30°S S. Orkney SouthGeorgia -100 -200 -300 -400 -500 -600 -700 -800 Current [m] Decrease ofOtolith Length, increase of OtolithHeight S. japonicus Channichthyidae Ch. aceratus Ps. georgianus C. gunnari increaseofOtolithLength A S WAT E R T E M P E R AT U R E I S D R O P P I N G M andflattening (T plane) T M T M T T T T M M M M M 21 km/h 0.9 km/h 0.1 km/h 1 km/h
- 4. 4 N3=6; MOśr=0,032±0,00185g; s=0,0023; TLśr=37 cm; MCśr=453,3g; Gśr=1,7; Żśr=0,8 N4=102; MOśr=0,043±0,00051g; s=0,0026; TLśr=47,1 cm; MCśr=1113,7g; Gśr=3,7; Żśr=0,8 N5=19; MOśr=0,052±0,00119g; s=0,0027; TLśr=49,1 cm; MCśr=1229,2g; Gśr=3,4; Żśr=0,5 0 1 2 3 4 5 6 7 0 1 2 3 4 5 6 7 8 9 10 11 Agegroup N OW [g] South Orkney Is 30.XII.1978 N6=5; MOśr=0,067±0,00126 g; s=0,0014; TLśr=50,8 cm; MCśr=1471g; Gśr=3,4; Żśr=0,4 N3=26;MOśr=0,034±0,00058g; s=0,0015;TLśr=43,2cm; MCśr=752,5g; Gśr=2,3; Żśr=1,7 N4=9; MOśr=0,041±0,0011g; s=0,0017; TLśr=47,6cm; MCśr=1158,9g; Gśr=4; Żśr=2 N5=3; MOśr=0,052±0,00233g; s=0,0021; TLśr=52,3cm; MCśr=1673,3g; Gśr=3,3; Żśr=2,7 0 1 2 3 4 5 6 7 8 0 1 2 3 4 Agegroup N OW [g] King George, 25.III.1979 N=39 N2=1; MO=0,021; TL=31cm; MC=210g; G=1; Ż=2 Age groups of Ps. georgianus seperated in time scale confirm geographical divide of otolith mass frequency as separate age groups on Antarctic islands: 2 aged at Pamer Archipelago, in February, 3 aged at King George in March, and 4 aged and older fish at S. Orkney in December. N3=62; MOśr=0,033±0,00045g; s=0,0018; TLśr=42,1cm; MCśr=740,6g; Gśr=2,4; Żśr=1,9 N4=20; MOśr=0,041±0,00126g; s=0,0029; TLśr=47,8cm; MCśr=1224,8g; Gśr=3,2; Żśr=2,5 N2=10; MOśr=0,023±0,00081g; s=0,0013; TLśr=30,4cm; MCśr=260g; Gśr=1,5; Żśr=1,2 N6=1; MO=0,064; TL=51cm; MC=1750g; G=3; Ż=3 0 1 2 3 4 5 6 7 0 1 2 3 4 5 6 7 8 9 10 11 0.010 0.012 0.014 0.016 0.018 0.020 0.022 0.024 0.026 0.028 0.030 0.032 0.034 0.036 0.038 0.040 0.042 0.044 0.046 0.048 0.050 0.052 0.054 0.056 0.058 0.060 0.062 0.064 0.066 0.068 0.070 0.072 0.074 Agegroup N OW [g] N5=4; MOśr=0,052±0,00151g; s=0,0015; TLśr=50,8 cm; MCśr=1548,8g; Gśr=3,8; Żśr=2 Palmer A., Deception, Elephant Is: 19-22.II.1979, N=97 Traczyk, 2012
- 5. 5 N3=6; MOśr=0,032±0,00185g; s=0,0023; TLśr=37 cm; MCśr=453,3g; Gśr=1,7; Żśr=0,8 N4=102; MOśr=0,043±0,00051g; s=0,0026; TLśr=47,1 cm; MCśr=1113,7g; Gśr=3,7; Żśr=0,8 N5=19; MOśr=0,052±0,00119g; s=0,0027; TLśr=49,1 cm; MCśr=1229,2g; Gśr=3,4; Żśr=0,5 0 1 2 3 4 5 6 7 0 1 2 3 4 5 6 7 8 9 10 11 Agegroup N OW [g] South Orkney Is 30.XII.1978 N6=5; MOśr=0,067±0,00126 g; s=0,0014; TLśr=50,8 cm; MCśr=1471g; Gśr=3,4; Żśr=0,4 N3=26;MOśr=0,034±0,00058g; s=0,0015;TLśr=43,2cm; MCśr=752,5g; Gśr=2,3; Żśr=1,7 N4=9; MOśr=0,041±0,0011g; s=0,0017; TLśr=47,6cm; MCśr=1158,9g; Gśr=4; Żśr=2 N5=3; MOśr=0,052±0,00233g; s=0,0021; TLśr=52,3cm; MCśr=1673,3g; Gśr=3,3; Żśr=2,7 0 1 2 3 4 5 6 7 8 0 1 2 3 4 Agegroup N OW [g] King George, 25.III.1979 N=39 N2=1; MO=0,021; TL=31cm; MC=210g; G=1; Ż=2 Age groups of Ps. georgianus seperated in time scale confirm geographical divide of otolith mass frequency as separate age groups on Antarctic islands: 2 aged at Pamer Archipelago, in February, 3 aged at King George in March, and 4 aged and older fish at S. Orkney in December. N3=62; MOśr=0,033±0,00045g; s=0,0018; TLśr=42,1cm; MCśr=740,6g; Gśr=2,4; Żśr=1,9 N4=20; MOśr=0,041±0,00126g; s=0,0029; TLśr=47,8cm; MCśr=1224,8g; Gśr=3,2; Żśr=2,5 N2=10; MOśr=0,023±0,00081g; s=0,0013; TLśr=30,4cm; MCśr=260g; Gśr=1,5; Żśr=1,2 N6=1; MO=0,064; TL=51cm; MC=1750g; G=3; Ż=3 0 1 2 3 4 5 6 7 0 1 2 3 4 5 6 7 8 9 10 11 0.010 0.012 0.014 0.016 0.018 0.020 0.022 0.024 0.026 0.028 0.030 0.032 0.034 0.036 0.038 0.040 0.042 0.044 0.046 0.048 0.050 0.052 0.054 0.056 0.058 0.060 0.062 0.064 0.066 0.068 0.070 0.072 0.074 Agegroup N OW [g] N5=4; MOśr=0,052±0,00151g; s=0,0015; TLśr=50,8 cm; MCśr=1548,8g; Gśr=3,8; Żśr=2 Palmer A., Deception, Elephant Is: 19-22.II.1979, N=97
- 6. 0 5 10 15 20 25 30 N TL [cm] 6 5 4 3 III IV V VI S. Orkney, 30.XII.1978; N = 142 Theoretical;Empirical AgeGroup N3=7; TL3=37,4±1,6 cm, N4=108; TL4=47±0,4 cm, s4=2,1 N5=21; TL5=49,2±0,7 s5=1,6 N6=6; TL6=51±1,3 0 5 10 15 20 25 N TL [cm] 6 5 4 3 2 II III IV V Palmer A., Deception, Elephant, 18-22.II.1979, N=171; Theoretical;Empirical AgeGroup N2=16; TL2=31,1±0,9 cm, s2=1,9 N3=97; TL3=41,8±0,4 s3=2 N4=41; TL4=48,2±0,5 s4=1,5 N5=10; TL5=50,9±0,8 s5=1,3 N6=7; TL6=51,7 ±0,8 s6=1,1 0 2 4 6 8 10 12 14 16 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 N TL [cm] 2 5 4 3 II III IV V Palmer A., Deception, King George, Elephant; 25.III.1979; N=145 AgeGroup Theoretical;empirical N2=24; TL2=30,8±0,7 cm, s2=1,7 N3=83; TL3=42±0,5 cm, s3=2,4 N4=34; TL4=48,5±0,5 cm, s4=1,4 N5=4; TL5=52,3 ±0,5 cm, s5=0,5 This same is for age groups in length frequency geographically divided on separated Antarctic Islands.
- 7. 0 5 10 15 20 25 30 N TL [cm] 6 5 4 3 III IV V VI S. Orkney, 30.XII.1978; N = 142 Theoretical;Empirical AgeGroup N3=7; TL3=37,4±1,6 cm, N4=108; TL4=47±0,4 cm, s4=2,1 N5=21; TL5=49,2±0,7 s5=1,6 N6=6; TL6=51±1,3 0 5 10 15 20 25 N TL [cm] 6 5 4 3 2 II III IV V Palmer A., Deception, Elephant, 18-22.II.1979, N=171; Theoretical;Empirical AgeGroup N2=16; TL2=31,1±0,9 cm, s2=1,9 N3=97; TL3=41,8±0,4 s3=2 N4=41; TL4=48,2±0,5 s4=1,5 N5=10; TL5=50,9±0,8 s5=1,3 N6=7; TL6=51,7 ±0,8 s6=1,1 0 2 4 6 8 10 12 14 16 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 N TL [cm] 2 5 4 3 II III IV V Palmer A., Deception, King George, Elephant; 25.III.1979; N=145 AgeGroup Theoretical;empirical N2=24; TL2=30,8±0,7 cm, s2=1,7 N3=83; TL3=42±0,5 cm, s3=2,4 N4=34; TL4=48,5±0,5 cm, s4=1,4 N5=4; TL5=52,3 ±0,5 cm, s5=0,5 This same is for age groups in length frequency geographically divided on separated Antarctic Islands.: 2 aged fish at Pamer A., in March, 3 aged fish dominated at Elephan in February, and 4 aged and olders fish at South Orkney in December.
- 8. 8 0 2 4 6 25 30 35 40 45 50 55 % TL [cm] Palmer A. N=59 27.1%; TL=30.8cm; s=1.8 45.8%; TL=40cm; s=1.7 25.4%; TL=47cm; s=1.5 1.7%; TL=51.5cm; s=0.5 0 5 10 15 20 % TL [cm] 12.3%; TL=30.6cm; s=1.6 60.7%; TL=41.7cm; s=2 20.9%; TL=47.9cm; s=1.4 4.3%; TL=50.4cm; s=0.8 1.8%; TL=51.1cm; s=0.2 Deception I., N=163 0 2 4 6 8 10 % TL [cm] 2.5%; TL=33.5cm; s=2.1 King George I., N=79 48.1%; TL=43.7cm; s=1.9 35.4%; TL=49.3cm; s=1.4 13.9%; TL=53.1cm; s=0.9 0 3 6 9 12 15 18 21 % TL [cm] S. Orkney, Western part, N=133 0.8%; TL=40.5cm; s=0.5 73.7%; TL=48cm; s=1.8 21.8%; TL=49.9cm; s=2.3 3.8%; TL=52.2cm; s=2 0 2 4 6 8 10 12 14 16 18 20 % TL [cm] S. Orkney, Eastern part, N=138 4.3%; TL=37cm; s=2.1 76.1%; TL=47.2cm; s=2.1 15.9%; TL=48.9cm; s=1.6 3.6%; TL=50.8cm; s=1.8 Geographical separation of age groups on shelf of different islands indicates that marine habitats of these areas have different properties.
- 9. 9 0 2 4 6 25 30 35 40 45 50 55 % TL [cm] Palmer Arch, N=59 27.1%; TL=30.8cm; s=1.8 45.8%; TL=40cm; s=1.7 25.4%; TL=47cm; s=1.5 1.7%; TL=51.5cm; s=0.5 0 5 10 15 20 % TL [cm] 12.3%; TL=30.6cm; s=1.6 60.7%; TL=41.7cm; s=2 20.9%; TL=47.9cm; s=1.4 4.3%; TL=50.4cm; s=0.8 1.8%; TL=51.1cm; s=0.2 Deception I., N=163 0 2 4 6 8 10 % TL [cm] 2.5%; TL=33.5cm; s=2.1 King Edward I., N=79 48.1%; TL=43.7cm; s=1.9 35.4%; TL=49.3cm; s=1.4 13.9%; TL=53.1cm; s=0.9 0 3 6 9 12 15 18 21 % TL [cm] S. Orkney, Western part, N=133 0.8%; TL=40.5cm; s=0.5 73.7%; TL=48cm; s=1.8 21.8%; TL=49.9cm; s=2.3 3.8%; TL=52.2cm; s=2 0 2 4 6 8 10 12 14 16 18 20 % TL [cm] S. Orkney, Eastern part, N=138 4.3%; TL=37cm; s=2.1 76.1%; TL=47.2cm; s=2.1 15.9%; TL=48.9cm; s=1.6 3.6%; TL=50.8cm; s=1.8 such that Ps. georgianus from the younger age groups prefer the western part of the Atlantic Antarctic, and the older age group East - North part. Age group is identified by otolith shape, which, indicate the different habitats, development stages and strategies of life: type of swimming.
- 10. 10 0 2 4 6 25 30 35 40 45 50 55 % TL [cm] Palmer Arch, N=59 27.1%; TL=30.8cm; s=1.8 45.8%; TL=40cm; s=1.7 25.4%; TL=47cm; s=1.5 1.7%; TL=51.5cm; s=0.5 0 5 10 15 20 % TL [cm] 12.3%; TL=30.6cm; s=1.6 60.7%; TL=41.7cm; s=2 20.9%; TL=47.9cm; s=1.4 4.3%; TL=50.4cm; s=0.8 1.8%; TL=51.1cm; s=0.2 Deception I., N=163 0 2 4 6 8 10 % TL [cm] 2.5%; TL=33.5cm; s=2.1 King Edward I., N=79 48.1%; TL=43.7cm; s=1.9 35.4%; TL=49.3cm; s=1.4 13.9%; TL=53.1cm; s=0.9 0 3 6 9 12 15 18 21 % TL [cm] S. Orkney, Western part, N=133 0.8%; TL=40.5cm; s=0.5 73.7%; TL=48cm; s=1.8 21.8%; TL=49.9cm; s=2.3 3.8%; TL=52.2cm; s=2 0 2 4 6 8 10 12 14 16 18 20 % TL [cm] S. Orkney, Eastern part, N=138 4.3%; TL=37cm; s=2.1 76.1%; TL=47.2cm; s=2.1 15.9%; TL=48.9cm; s=1.6 3.6%; TL=50.8cm; s=1.8 such that Ps. georgianus from the younger age groups prefer the western part of the Atlantic Antarctic, and the older age group East - North part. Age group is identified by otolith shape, which, indicate the different habitats, development stages and strategies of life: t ype of swimming . Otolith, M-plane, after Hecht, 1987 0.4 km/h 1.6 km/h and speed
- 11. BransfieldStrait - 1 - 1 0 1 1 2 3 4 >3 >4-2 0-1 1 2 3 4 5 22.6 98.1 South Orkney 14.4 5.1 56 58 60 62 64 B Feb.1979 – the R/V “Pr Nov. 78 – Feb. 79 – the Ps. georgianus A, B, C - transects Water temperature determine distribution of Antarctic fish? So older age groups fish 48 cm found at South Orkney Islands may have been resulted from lower temperature of waters below 0°C up to - 1°C. Younger age groups of 3 aged fish 43 cm found most numerous at Elephan have little warmer waters up to 0°C. The smallest fish 30 cm length of 2 age group appear at Palmer Archipelago have warmer water of above 0°C up to 1°C. Potential temperature [°C] at 200 m Sahrhage, 1988; Maslennikov, 1988; Sievers, 1988)
- 12. BransfieldStrait - 1 - 1 0 1 1 2 3 4 >3 >4-2 0-1 1 2 3 4 5 22.6 98.1 South Orkney 14.4 5.1 56 58 60 62 64 B Feb.1979 – the R/V “Pr Nov. 78 – Feb. 79 – the Ps. georgianus A, B, C - transects Water temperature determine distribution of Antarctic fish. So older age groups, fish 49 centimeters found at South Orkney Islands may have been resulted from lower temperature of waters below 0 up to minus 1 Celsius degree. Younger age groups of 3 aged fish 43 cm found most numerous at Elephan have little warmer waters up to 0 degrees centigrade. The smallest fish 30 cm length of II age group appear at Palmer Archipelago have warm water of above 0 up to 1 Celsius degree. Potential temperature [°C] at 200 m
- 13. BransfieldStrait - 1 - 1 0 1 1 2 3 4 >3 >4-2 0-1 1 2 3 4 5 22.6 98.1 South Orkney 14.4 5.1 56 58 60 62 64 B Feb.1979 – the R/V “Pr Nov. 78 – Feb. 79 – the Ps. georgianus A, B, C - transects Potential temperature [°C] at 200 m
- 14. BransfieldStrait - 1 - 1 0 1 1 2 3 4 >3 >4-2 0-1 1 2 3 4 5 22.6 98.1 South Orkney 14.4 5.1 56 58 60 62 64 B Feb.1979 – the R/V “Pr Nov. 78 – Feb. 79 – the Ps. georgianus A, B, C - transects Potential temperature [°C] at 200 m
- 15. Swimming of young fish in habitat thermally other than for elderly fish may be due to a decrease (increase) with the age: the production or activity of AFGP antifreeze protein. Having AFGP Channichthyidae prosper in icy cavities and crevices Detrich at all, 2012, Bilyk 2011
- 16. A1-6 A2 A1 A3 A4 A5 A6 AFGP is adsorbing and form Hydrogen bond with ice network Th-OH····OH2 w sposób zamka błyskawicznego A2 A1 A3 A4 A5 A6 AFGP decrease temperature of blood freezing at or below -2.2 ° C, which is lower than the freezing point of sea water = -1.9 ° C. It is by inhibit the growth of ice embryo into star ice crystals AFGP adsorption Peltier at all, 2010, Wohrmann, 1996
- 17. 0 year by 0.21°C C. aceratus from larvae to fish of 2 year old, living in a warm pelagic water and preying krill, has weaker activity of AFGP proteins (equals to 0,21-0,41 °C) than adult fish. Adult fish at the age of 4.5 years, while descend to the colder depths of -1.47 °C have large activity of AFGP, that reduce freezing point of blood by 0.57°C, that is accompanied with 4 times increase of their swimming possibility and diet change to eating fish. Young fish with delayed activation of AFGP protein can avoid cold water and swim to North, to water without ice. Traczyk, 2013; Arkive nature.pl; Hureau, 1985 2 years by 0.41°C of freezing point 4.2 years by 0.57°C Bilyk 2011 0.45 km/h 0.1 km/h 1.8 km/h
- 18. E F G P a c t i v i t y ( ) d r o p p o f f r e e z i n g p o i n t b y food: fish ×4 large swimming speed Arkive nature.pl; Hureau, 1985C. aceratus 0.45 km/h 0.1 km/h 1.8 km/h
- 19. food type: fish ×4 large swimming speed Arkive nature.pl; Hureau, 1985 Protein activity of AFGP is suspended in icefish. It increases during adaptation of fish development stages to the transition from pelagic to bottom, or from warm to cold water. C. aceratus 0.45 km/h 0.1 km/h 1.8 km/h
- 20. It seems to be the role, because also concentration of protein activity AFGP for the high Antarctic fish depends on the stage of development of the fish - synchronizing with space system of water temperature and food type (Wöhrmann 1996). -300 -100 -200 -400 _500 E a s t W i n d D r i f t Pleurogramma antarcticumcm, SL 22 cm spawning adults I c e s h e l f 0.15 0.133 0.219 0.109 0.17 0.139 0.10 1 0.194 0.188 larvae, 1 year SL 6 cm juvenes, SL 10 cm T < -0,5 °C Ice Shelves Water T < -2,0 °C larvae, 0 year juvenes, 2 year AFGP (% wet weight) high molar m. AFGP (% wet weight) low molar m. AFGP (% wet weight) Wohrmann, 1996 0.3 km/h
- 21. C. gunnari: 0,98°C -1,85°C Ch. wilsoni: 1,29°C -2,23°C Antarctic Circumpolar Ps. georgianus: 1,03°C -1,91°C Cold water descent Activity AFGP [°C] Blood feeezing: [°C] Concentration, the activity of the AFGP protein in the white blooded fish increasing in colder water in the direction to the pole, and with the depth., Bilyk, 2011. Ch. hamatus: 1,45°C -2,44°C WHITE-BLOODED: high Antarctic; ice pack zone; temperate 80°S 74, ~70°S; 63, ~60°S; 63, ~57°30’S; 52, ~45°S 30°S S. Orkney SouthGeorgia -100 -200 -300 -400 -500 -600 -700 -800 Current C. aceratus: 0,54°C -1,47°C [m]
- 22. BransfieldStrait - 1 - 1 0 1 1 2 3 4 >3 >4-2 0-1 1 2 3 4 5 22.6 98.1 South Orkney 14.4 5.1 56 58 60 62 64 B Feb.1979 – the R/V “Pr Nov. 78 – Feb. 79 – the Ps. georgianus A, B, C - transects Separation of different habitats between different stages of development (different age) provides a better use of environmental resources (food). It seems to be a role also for Ps. georgianus - with appropriate small correction that Ps. georgianus has about 2 times greater activity of frost protection of AFGP than C. aceratus. Pelagic young fish in shallow warm water swim slower – they have drop of freezing point by ~0.21 Celsius degrees. Pelagic ones that, in transition to bottom, swim with average speed – they have dropping freezing point by ~0.41 Celsius degrees. Bottom fish, in cold water, swim 4 time faster – they have dropping freezing point by ~0.57 Celsius degrees. Potential temperature [°C] at 200 m 1978/79
- 23. C. gunnari: 0,98°C -1,85°C Ch. wilsoni: 1,29°C -2,23°C Antarctic Circumpolar Ps. georgianus: 1,03°C -1,91°C Cold water descent Activity AFGP [°C] Blood feeezing: [°C] Ps. georgianus in compare to C. aceratus has a higher activity of AFGP = 1.03 °C, can swim in colder waters up to -1.91°C and finally is larger predator. Ch. hamatus: 1,45°C -2,44°C WHITE-BLOODED: high Antarctic; ice pack zone; temperate 80°S 74, ~70°S; 63, ~60°S; 63, ~57°30’S; 52, ~45°S 30°S S. Orkney SouthGeorgia -100 -200 -300 -400 -500 -600 -700 -800 Current C. aceratus: 0,54°C -1,47°C [m] Bilyk 2011
- 24. 3; 19; 52 % 9; 30; 55; 86 % 20; 46; 63 % 15; 41; 56 % 0 10 20 30 40 6 11 16 21 26 31 36 41 46 51 56 % 49 mm; 983 mg;1394 szt/h; 137 kg/h z56; z65; z66 65 66 55 56 67 69 40 73 71 41 82 8593 78 74 45°47°49°51°53°55°57° 59° 62° 60° 61° 45°47°49°51°53°55°57° 62° 60° 61° 62 61 5853 52 50 46 45 43 39 37 35 33 31 27 25 24 22 48 57 6; 34; 51; 86 % 5; 35 % Krill catches in the sea ice zone of the Scotia Sea in1988/89 0 5 10 15 20 6 11 16 21 26 31 36 41 46 51 56 % SL, mm 52 mm; 1171 mg; 3339 szt/h; 391 kg/h z78 0 5 10 15 20 25 6 11 16 21 26 31 36 41 46 51 56 % SL, mm 29 mm, I.; 158 mg; 712 szt/h, 11 kg/h z71; z73; z74 0 5 10 15 20 6 11 16 21 26 31 36 41 46 51 56 % SL, mm 27 mm, XII; 139 mg; 537 szt/h, 7 kg/h 37 mm; 392 mg; 774 szt/h; 30 kg/h z40; z41 0 10 20 30 6 11 16 21 26 31 36 41 46 51 56 % SL, mm 25 mm, XII; 107 mg; 25,3 ml/1000m3 b39; b45 There is synchronization of the size of the fish, of its ability to swim to the size of the food. In 1989 at the Western shores of the S. Orkney and Elephant Islands, where most of Ps. georgianus were older and larger, there were also adult and large krill. Additionally on West and Southern shores of the King George Is., there were small juvenes krill and larvae of Ps. georgianus that can swim in colder water. bongo net 3-364 kg/h; II-III 2009; SGI=753 2-0°C <0°C XII.1996, SGI Data: Traczyk, 1993, Traczyk, 2012; Van Cise, 2009; White 1998.
- 25. 3; 19; 52 % 9; 30; 55; 86 % 20; 46; 63 % 15; 41; 56 % 0 10 20 30 40 6 11 16 21 26 31 36 41 46 51 56 % 49 mm; 983 mg;1394 szt/h; 137 kg/h z56; z65; z66 65 66 55 56 67 69 40 73 71 41 82 8593 78 74 45°47°49°51°53°55°57° 59° 62° 60° 61° 45°47°49°51°53°55°57° 62° 60° 61° 62 61 5853 52 50 46 45 43 39 37 35 33 31 27 25 24 22 48 57 6; 34; 51; 86 % 5; 35 % krill catches in the sea ice zone of the Scotia Sea in1988/89 0 5 10 15 20 6 11 16 21 26 31 36 41 46 51 56 % SL, mm 52 mm; 1171 mg; 3339 szt/h; 391 kg/h z78 0 5 10 15 20 25 6 11 16 21 26 31 36 41 46 51 56 % SL, mm 29 mm, I.; 158 mg; 712 szt/h, 11 kg/h z71; z73; z74 0 5 10 15 20 6 11 16 21 26 31 36 41 46 51 56 % SL, mm 27 mm, XII; 139 mg; 537 szt/h, 7 kg/h 37 mm; 392 mg; 774 szt/h; 30 kg/h z40; z41 0 10 20 30 6 11 16 21 26 31 36 41 46 51 56 % SL, mm 25 mm, XII; 107 mg; 25,3 ml/1000m3 b39; b45 There is synchronization of the size of the fish, of its ability to swim to the size of the food. In 1989 at the Western shores of the S. Orkney and Elephant Islands, where most of Ps. georgianus were older and larger, there were also adult and large krill. Additionally on West and Southern shores of the King George Is., there were small juvenes krill and larvae of Ps. georgianus that can swim in colder water. bongo net 3-364 kg/h; II-III 2009; SGI=753 2-0°C <0°C XII.1996, SGI
- 26. 26 kg/h 0 25 50 75 100 23 kg/h 3 kg/h14 kg/h 78 82 74 73 40 41 69 67 66 65 56 55 71 larvaehaul No 40 41 55 56 65 66 67 69 71 73 74 78 82 larvaehaul No 40 41 55 56 65 66 67 69 71 73 74 78 82 coastal demersal bottom shelf, deep-water C. aceratus 5 1 Cr. antarcticus 1 1 4 1 C. rastrospinosus 5 2 1 Neopagetopsis sp. 1 C. wilsoni 1 3 2 3 bathypelagic T. eulepidotus 1 N. ionach 2 1 1 L. larseni 1 1 N. coatsi 2 1 P. macropterus 2 pelagic, oceanic Notothenia sp. 1 Pl. antarcticum 1 4 coastal pelagic E. carlsbergi 20 C. gunnari 1 E. antarctica 48 35 Pagetopsis sp. 5 2 3 1 G. opisthopterus 1 bottom shelf, seamounts - ice edge: 26.XII.88 - 8.I. 89; 9.I - 13.I.89 D. eleginoides 2 Seamounts and ice combine S. Orkney with Elephant I. SGI - 1989 Anon. 1990
- 27. 27 kg/h 0 20 40 60 80 100 23 kg/h 3 kg/h14 kg/h 78 82 74 73 40 41 69 67 66 65 56 55 71 Ice edge connecting K. George, Elephant with S. Orkney, were located over mountains that combine undersea above islands. Undersea seabed belt of mountains and ridges play role of channel for conducting sea currents which carries krill and fish larvaes
- 28. Geographic age groups in otolith shape, we can find being in well agreement with their food size of krill migrating, drifting and growing with currents to the East. II age group were catch at Palmer Archipelago among juvenile krill. Adults fish, age IV and above were cought at S. Orkney Island among adults krill. 58 ° 60 ° 62 ° 64 ° Feb.1979, R/V “Prof. Siedlecki” (N=67) Nov 78 – Feb 79 – M/T “Sirius” (N=30) Krill (A- adult, S- small, J- juveniles) migration in the spawning period spring - summer extended by ice edge to the S. Orkney Is. Ps. georgianus catch Orkady Pd. S J S S S S S J A A A A J A J S S A A A A S J 90 g/m3 200 g/m3 150 g/m3 350 g/m3 73 g/m3 60 ° 56 ° 52 ° 48 ° 44 °64 ° 60 ° 62 ° 64 ° 60 ° 56 ° 52 ° 48 ° 44 °64 ° geostrophic current Krill migrations Ice edge 1 - 8 Jan 1989 Ice edge 8 - 15 Jan 1989 Weddell-Scotia Confluence Fish and environment data: Sahrhage, 1988, Traczyk, 2012; Sahrhage, 1988; Witek, 1988; Ślósarczyk, 1985; White, 1998
- 29. Geographic age groups in otolith shape, we can find being in well agreement with their food size of krill migrating, drifting and growing with currents to the East. The smallest fish of 1979 fishing: II age group were catch at Palmer Archipelago among juvenile krill. Ps. georgianus of III age group were at King George Is among average krill. Adults fish, age IV and above were cought at Orkney Is., among adults krill, occurs there along the continental slope. At the coast smaller krill was dominated. Fish juvenile were at the coastal waters of K. George, where krill juvenes was. Fish larvaes this species were at Palmer Archipelago and King George I. 58 ° 60 ° 62 ° 64 ° Feb.1979, R/V “Prof. Siedlecki” (N=67) Nov 78 – Feb 79 – M/T “Sirius” (N=30) Krill (A- adult, S- small, J- juveniles) migration in the spawning period spring - summer extended by ice edge to the S. Orkney Is. Ps. georgianus catch Orkady Pd. S J S S S S S J A A A A J A J S S A A A A S J 90 g/m3 200 g/m3 150 g/m3 350 g/m3 73 g/m3 60 ° 56 ° 52 ° 48 ° 44 °64 ° 60 ° 62 ° 64 ° 60 ° 56 ° 52 ° 48 ° 44 °64 ° geostrophic current Krill migrations Ice edge 1 - 8 Jan 1989 Ice edge 8 - 15 Jan 1989 Weddell-Scotia Confluence Sahrhage, 1988, Traczyk, 2012 Krill as main food: (Sarah Clarke, 2008; Chojnacki, 1987)
- 30. 58 ° 60 ° 62 ° 64 ° Feb.1979, R/V “Prof. Siedlecki” (N=67) Nov 78 – Feb 79 – M/T “Sirius” (N=30) Krill (A- adult, S- small, J- juveniles) migration in the spawning period spring - summer extended by ice edge to the S. Orkney Is. Ps. georgianus catch Orkady Pd. S J S S S S S J A A A A J A J S S A A A A S J 90 g/m3 200 g/m3 150 g/m3 350 g/m3 73 g/m3 60 ° 56 ° 52 ° 48 ° 44 °64 ° 60 ° 62 ° 64 ° 60 ° 56 ° 52 ° 48 ° 44 °64 ° geostrophic current Krill migrations Ice edge 1 - 8 Jan 1989 Ice edge 8 - 15 Jan 1989 Weddell-Scotia Confluence Sahrhage, 1988, Traczyk, 2012
- 31. While small and juvenes krill asocjate inshore, adult krill in spring and summer migrate onshore to North for spawning and move with currents to East and this suggest that their active swimming carnivores follow after them and acumulated in the East - as the older and large fish were found there. 58 ° 60 ° 62 ° 64 ° Feb.1979, R/V “Prof. Siedlecki” (N=67) Nov 78 – Feb 79 – M/T “Sirius” (N=30) Krill (A- adult, S- small, J- juveniles) migration in the spawning period spring - summer extended by ice edge to the S. Orkney Is. Ps. georgianus catch S. Orkney S J S S S S S J A A A A J A J S S A A A A S J 90 g/m3 200 g/m3 150 g/m3 350 g/m3 73 g/m3 60 ° 56 ° 52 ° 48 ° 44 °64 ° 60 ° 62 ° 64 ° 60 ° 56 ° 52 ° 48 ° 44 °64 ° geostrophic current Krill migrations Ice edge 1 - 8 Jan 1989 Ice edge 8 - 15 Jan 1989 Weddell-Scotia Confluence Sahrhage, 1988, Traczyk, 2012 Fish and environment data: Sahrhage, 1988, Traczyk, 2012; Witek, 1988; Ślósarczyk, 1985; White, 1998; Siegel, 1988
- 32. On the North along the continental slope were adult krill migrate for spawning there were Southern Front of the Antarctic Circumpolar Current leading its life to the East near S. Orkney. This current is the largest current in the World. On the Southern shores of South Shetland there are Bransfield Current, which carries smaller life from Shetlads to South Orkney. 58 ° 60 ° 62 ° 64 ° Feb.1979, R/V “Prof. Siedlecki” (N=67) Nov 78 – Feb 79 – M/T “Sirius” (N=30) (A- adult, S- small, J- juveniles) migration in the spawning period spring - summer extended by ice edge to the S. Orkney Is. Ps. georgianus catch: South Orkney I.S 150 g/m3 350 g/m3 73 g/m3 60 ° 56 ° 52 ° 48 ° 44 °64 ° 60 ° 62 ° 64 ° 60 ° 56 ° 52 ° 48 ° 44 °64 ° Surface currents <200 Ice edge 1 - 8 Jan 1989 Ice edge 8 - 15 Jan 1989 Weddell-Scotia Confluence Krill migrations 90 g/m3 J SS S S J A A A J A J S S A A A A A S J S 200 g/m3 Deep currents >200 m Fish and environment data: Sahrhage, 1988, Traczyk, 2012; Sahrhage, 1988; Witek, 1988, BAS, 2014, Murphy, 2013; Ślósarczyk, 1985; White, 1998
- 33. On the North along the continental slope were adult krill migrate for spawning (catched by adult Ps. georgianus), there were Southern Front of the Antarctic Circumpolar Current leading its life to the East near S. Orkney. This current is the largest current in the World. On the Southern shores of South Shetland were small and juvenes both krill and Ps. georgianus were cought, there are Bransfield Current, branch of the SF ACC, which carries smaller life near ice edges from Shetlads over seabed ridges to S. Orkney 58 ° 60 ° 62 ° 64 ° Feb.1979, R/V “Prof. Siedlecki” (N=67) Nov 78 – Feb 79 – M/T “Sirius” (N=30) (A- adult, S- small, J- juveniles) migration in the spawning period spring - summer extended by ice edge to the S. Orkney Is. Ps. georgianus catch: South Orkney I.S 150 g/m3 350 g/m3 73 g/m3 60 ° 56 ° 52 ° 48 ° 44 °64 ° 60 ° 62 ° 64 ° 60 ° 56 ° 52 ° 48 ° 44 °64 ° Surface currents <200 Ice edge 1 - 8 Jan 1989 Ice edge 8 - 15 Jan 1989 Weddell-Scotia Confluence Krill migrations 90 g/m3 J SS S S J A A A J A J S S A A A A A S J S 200 g/m3 Deep currents >200 m
- 34. Thus on the North–East of Shetland Islands large adults Ps. georgianus, of age IV (and above) from King George to South Orkney have to withstand stronger surface currents and whirls. In opposite South–West side of Shetlands larvaes and small fish have protective environment on their shelves extended by ice edge zone to the South Orkney. 58 ° 60 ° 62 ° 64 ° Feb.1979, R/V “Prof. Siedlecki” (N=67) Nov 78 – Feb 79 – M/T “Sirius” (N=30) Krill (A- adult, S- small, J- juveniles) migration in the spawning period spring - summer extended by ice edge to the S. Orkney Is. Ps. georgianus catch South Orkney I.S J S S S S S J A A A A J A J S S A A A A S J 90 g/m3 200 g/m3 150 g/m3 350 g/m3 73 g/m3 60 ° 56 ° 52 ° 48 ° 44 °64 ° 60 ° 62 ° 64 ° 60 ° 56 ° 52 ° 48 ° 44 °64 ° geostrophic current Krill migrations Ice edge 1 - 8 Jan 1989 Ice edge 8 - 15 Jan 1989 Weddell-Scotia Confluence
- 35. Thus on the North – East of Shetland islands large adults Ps. georgianus, of age IV and above from King George to South Orkney have to withstand stronger surface currents and whirls – large fish this species swim in shallow water. In opposite South – West side of Shetlands larvaes and small fish have protective environment on their shelves extended by ice edge zone to the S. Orkney. In addition there are confluence of Scotia Sea and Weddell Sea, greate source of strong whirls and countercurrents. 58 ° 60 ° 62 ° 64 ° Feb.1979, R/V “Prof. Siedlecki” (N=67) Nov 78 – Feb 79 – M/T “Sirius” (N=30) Krill (A- adult, S- small, J- juveniles) migration in the spawning period spring - summer extended by ice edge to the S. Orkney Is. Ps. georgianus catch South Orkney I.S J S S S S S J A A A A J A J S S A A A A S J 90 g/m3 200 g/m3 150 g/m3 350 g/m3 73 g/m3 60 ° 56 ° 52 ° 48 ° 44 °64 ° 60 ° 62 ° 64 ° 60 ° 56 ° 52 ° 48 ° 44 °64 ° geostrophic current Krill migrations Ice edge 1 - 8 Jan 1989 Ice edge 8 - 15 Jan 1989 Weddell-Scotia Confluence
- 36. .15 .20 .25 .30 .35 .40 .30 .20 47.8 22.6 3.2 14.4 5.1 98.1 South Orkney I. South Georgia I. S c o t i a S e a W e d d e l l S e a 52 48 44 40 36 54 56 58 60 62 64 Elephant I. Feb.1979 – the R/V “Prof. Siedlecki” (N=67) Nov. 78 – Feb. 79 – the M/T “Sirius” (N=30) Ps. georgianus capture, [kg/h] 54 56 58 60 62 64 64 60 56 52 48 44 40 36 .31 .20 .19 .24 .35 .26 .28 .30 Secondary Frontal Zone Area of high krill abundance Current flow rel. to 5MPa surface [Dyn m] Weddell-Scotia Confluence (s>34‰ at 20m) WSC 73 200 350 90 150 550 23 14 krill density [g/m2] and cluster extending 48 The Antarctic Circumpolar Current, the Weddell Scotia Confluence creates in Shetland a system of currents, whirls and countercurrents, where Ps. georgianus lives and to which it has to have a strong adaptation to survive.
- 37. .15 .20 .25 .30 .35 .40 .30 .20 47.8 22.6 3.2 14.4 5.1 98.1 South Orkney I. South Georgia I. S c o t i a S e a W e d d e l l S e a 52 48 44 40 36 54 56 58 60 62 64 Elephant I. Feb.1979 – the R/V “Prof. Siedlecki” (N=67) Nov. 78 – Feb. 79 – the M/T “Sirius” (N=30) Ps. georgianus capture, [kg/h] 54 56 58 60 62 64 64 60 56 52 48 44 40 36 .31 .20 .19 .24 .35 .26 .28 .30 Secondary Frontal Zone Area of high krill abundance Current flow rel. to 5MPa surface [Dyn m] Weddell-Scotia Confluence (s>34‰ at 20m) WSC 73 200 350 90 150 550 23 14 krill density [g/m2] and cluster extending 48 This system together with ice edge zone inhabited by krill and fish larvaes have great potential for migration of fish even for shelf species. Such conditions are often extend to South Georgia
- 38. The currents have great influence on krill distribution. Typically, krill is carried from South Shetland Is., and from the Weddell Sea through Bransfield Strait into the Antarctic Circumpolar Current up to outside of South Georgia. However, the extreme south movement of air move currents and thicken the abundance of larger krill in the areas of South Orkney Island.Sahrhage, 1988, Vincent, 1988
- 39. kilometry 50°W 50°W 1+ mg/m2 Chl a 0-0,2 mg/m2 Chl a 1+ mg/m2 Chl a 40°W Antarctic Peninsula Nowadays ice cover in winter has very small range, while in the past in the 80s can reached South Georgia.Marschall, 1988; 2012; Węsławski, 2011; Damerau M., 2014; BAS, 2014; Kaufmann R.S., 1995; Murphy, 2013)
- 40. kilometry 50°W 50°W 1+ mg/m2 Chl a 0-0,2 mg/m2 Chl a 1+ mg/m2 Chl a 40°W Antarctic Peninsula In summer similar: ice cover in the 80s can reached South Orkney while now is only near Antarctic Peninsula
- 41. .15 .20 .25 .30 .35 .40 .30 .20 47.8 22.6 3.2 14.4 5.1 98.1 South Orkney I. South Georgia I. S c o t i a S e a W e d d e l l S e a 52 48 44 40 36 54 56 58 60 62 64 Elephant I. Feb.1979 – the R/V “Prof. Siedlecki” (N=67) Nov. 78 – Feb. 79 – the M/T “Sirius” (N=30) Ps. georgianus capture, [kg/h] 54 56 58 60 62 64 64 60 56 52 48 44 40 36 .31 .20 .19 .24 .35 .26 .28 .30 Secondary Frontal Zone Area of high krill abundance Current flow rel. to 5MPa surface [Dyn m] Weddell-Scotia Confluence (s>34‰ at 20m) WSC 73 200 350 90 150 550 Ps. georgianus show different geographic distributions in number of fish. At S. Georgia icefish were more numerous. This could be related to swimming posibility: at S. Georgia we can see strong turbulences and eddys, and also more krill for food . 23 14 krill density [g/m2] and cluster extending 48 Marschall, 1988; 2012; Sahrhage, 1988; Siegel, 1988
- 42. 10 20 30 40 50 1 9 7 8 / 7 9 [kg/hCloser to the continent catch drops 2 × (on r/v „Prof. Siedlecki”). S. Georgia S. Orkney. K. George Palmer A. 10 kg/h 20 30 40 50 47.8 22.6 3.2 14.4 5.1 98.1 South Orkney I. South Georgia I. S c o t i a S e a W e d d e l l S e a 60 56 52 48 44 40 36 54 56 58 60 62 64 Elephant I. Feb.1979 – the R/V “Prof. Siedlecki” (N=67) Nov. 78 – Feb. 79 – the M/T “Sirius” (N=30) Ps. georgianus capture, [kg/h] 54 56 58 60 62 64 64 60 56 52 48 44 40 36 0-500 m 48 23 14 K. George Traczyk, 2012
- 43. 47.8 22.6 3.2 14.4 5.1 98.1 South Orkney I. South Georgia I. S c o t i a S e a W e d d e l l S e a 60 56 52 48 44 40 36 54 56 58 60 62 64 Elephant I. Feb.1979 – the R/V “Prof. Siedlecki” (N=67) Nov. 78 – Feb. 79 – the M/T “Sirius” (N=30) Ps. georgianus capture, [kg/h] 54 56 58 60 62 64 64 60 56 52 48 44 40 36 48 23 14 0-500 m K. George 2° 0° >2° >2° >2° >2° >2° <0° <0° 2° 0° average water temperature in the summer at a depth of ~20 m Isotherm [°C]
- 44. 47.8 22.6 3.2 14.4 5.1 98.1 South Orkney I. South Georgia I. S c o t i a S e a W e d d e l l S e a 60 56 52 48 44 40 36 54 56 58 60 62 64 Elephant I. Feb.1979 – the R/V “Prof. Siedlecki” (N=67) Nov. 78 – Feb. 79 – the M/T “Sirius” (N=30) 54 56 58 60 62 64 64 60 56 52 48 44 40 36 0-500 m 48 23 14 Ps. georgianus capture, [kg/h]King George I. The density and extent of krill clusters [g/m3] Traczyk, 2013
- 45. kilometry 50°W 50°W 1+ mg/m2 Chl a 0-0,2 mg/m2 Chl a 1+ mg/m2 Chl a 40°W Antarctic Peninsula
- 46. 47.8 22.6 14.4 5.1 98.1 South Orkney I. South Georgia I. S c o t i a S e a W e d d e l l S e a 60 56 52 48 44 40 36 54 56 58 60 62 64 Elephant I. Feb.1979 – the R/V “Prof. Siedlecki” (N=67) Nov. 78 – Feb. 79 – the M/T “Sirius” (N=30) 54 56 58 60 62 64 64 60 56 52 48 44 40 36 0-500 m 48 23 3.2 14 Ps. georgianus capture, [kg/h]King George I. P.A. The density [g/m3] and extent of krill clusters
- 47. 22 K mt 4 K mt 500 mt P - Palmer A. & S. Shetland: D – Deception, KG – King George, E – Elephan Ps. georgianus catch for 1975 to 2012 confirm trend; catch became seven times smaller in the direction to the continent. A few fish at South Sangwich Is (S). G – S. Georgia (SR – Shag Rock) - 21733 mt O – S. Orkney - 3687 mt P & S. Shetland- 473 mt 100 200 300 400 500 600 800 K 2K 3K 4K 5K 6K 7K 8K 10K 20K Traczyk, 2012
- 48. 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 55° 54° 30' 30' 40° 39° 38° 37° 36° 35° 34° 53° 55° 54° 30' 30' 30' 53° >1°C; <2°C >0°C; <1°C Temperature et depth 200 m 0 5 10 15 14 19 24 29 34 39 44 49 54 33,8 cm TL Haul 17, 24, 25 0 1 2 3 14 19 24 29 34 39 44 49 54 51,2 cm, TL Haul: 91, 108 0 1 2 3 14 19 24 29 34 39 44 49 54 45,9 cm, TL zac: 70, 75, 76 0 1 2 3 4 5 6 11 16 21 26 31 36 41 46 51 56 2 21 cm TL haul.: 102, 106, 110, 111, 112 0 1 2 3 4 5 13 18 23 28 33 38 43 48 53 58 haul. 77, 80, 79, 88, 82 28.1 cm, N=18, grupa wieku: 1, 2 0 10 20 30 13 18 23 28 33 38 43 48 53 58 haul: 15P, 16P, 27, 1P 44.9 cm; N=164 Age group: 3, 4, 5, 6 0 1 14 19 24 29 34 39 44 49 54 haul. 53, 52 28,2 cm; N=5; grupa wieku: 1, 2, 3 0 1 2 14 19 24 29 34 39 44 49 54 59 haul 28 59 cm; N=1; N=318 Ps. georgianus 18.XII.87-08.I.88 34° 34° 1P 2 4P 5 5P 7P 8 10 11 13 15p 16p 1719 20 21 23 24 25 26 27 28 4046 52 53 70 75 76 77 79 80 82 88 90 91 94 95 101 102 103 105 106 108110 111112 113 122 124 116 69 68 115 85 13P - 1; postlarvae/h from 0 ege group - 6; 24; 72 fish/h from age group III, IV, V, VI - 3; 11 fish/h, from I age group - 3; 9 126 fish/h,from II age group Traczyk, 2013
- 49. 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 55° 54° 30' 30' 40° 39° 38° 37° 36° 35° 34° 53° 55° 54° 30' 30' 30' 53° >1°C; <2°C >0°C; <1°C Temperature et depth 200 m 0 5 10 15 14 19 24 29 34 39 44 49 54 33,8 cm TL Haul 17, 24, 25 0 1 2 3 14 19 24 29 34 39 44 49 54 51,2 cm, TL Haul: 91, 108 0 1 2 3 14 19 24 29 34 39 44 49 54 45,9 cm, TL zac: 70, 75, 76 0 1 2 3 4 5 6 11 16 21 26 31 36 41 46 51 56 2 21 cm TL haul.: 102, 106, 110, 111, 112 0 1 2 3 4 5 13 18 23 28 33 38 43 48 53 58 haul. 77, 80, 79, 88, 82 28.1 cm, N=18, grupa wieku: 1, 2 0 10 20 30 13 18 23 28 33 38 43 48 53 58 haul: 15P, 16P, 27, 1P 44.9 cm; N=164 Age group: 3, 4, 5, 6 0 1 14 19 24 29 34 39 44 49 54 haul. 53, 52 28,2 cm; N=5; grupa wieku: 1, 2, 3 0 1 2 14 19 24 29 34 39 44 49 54 59 haul 28 59 cm; N=1; N=318 Ps. georgianus 18.XII.87-08.I.88 34° 34° 1P 2 4P 5 5P 7P 8 10 11 13 15p 16p 1719 20 21 23 24 25 26 27 28 4046 52 53 70 75 76 77 79 80 82 88 90 91 94 95 101 102 103 105 106 108110 111112 113 122 124 116 69 68 115 85 13P - 1; postlarvae/h from 0 ege group - 6; 24; 72 fish/h from age group III, IV, V, VI - 3; 11 fish/h, from I age group - 3; 9 126 fish/h,from II age group
- 50. 50 42° 41° 40° 39° 38° 37° 36° 35° 34°43° 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 54° 55° Shag Rocks 55° 54° 500m 500m 30' 30' 30' 150m 30' 30' 30' Currents at surface at 5MPa [Dyn m] WSC-Weddella-Scotia Confluence (s > 34‰ per 20 m) krill high density regions Secondary Frontal Zone, SFZ 0,3 0,3 0,35 Krill550g/m3 Witek, 1988; Sahrhage, 1988
- 51. 51 42° 41° 40° 39° 38° 37° 36° 35° 34°43° 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 54° 55° Shag Rocks 55° 54° 500m 500m 30' 30' 30' 150m 30' 30' 30' WSC-Weddella-Scotia Confluence (s > 34‰ per 20 m) krill high density regions Secondary Frontal Zone, SFZ 0,3 0,3 0,35 Krill550g/m3 Currents at surface at 5MPa [Dyn m] Ps. georgianus has krill as dominant food (Sarah Clarke, 2008; Chojnacki, 1987).
- 52. 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 55° 54° 30' 30' 30' 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 53° 55° 54° 30' 30' 30' 53° >1°C; <2°C >0°C; <1°C Temperature at depth 200 m N=676 Ps. georgianus 1-13.II.89 1 2 3 5 6 7 8 9 10 11 12 14 15 17 18 19 2021 22 23 24 252627 28 29 30 31 32 3436 37 38 39 4041 43 44 45 46 47 48 51 52 54 55 59P 60P 57P 58P wieku III, IV, V, VI. - 5 postlarv/h; 15 postlarvae/h of age group 0. - 10 fish/h; 15 101 fish/h; 330 fish/h of age - 2 fish/h; 15 54 fish/h, of age group I. - 4 fish/h; 15 20 fish/h, of age group II. 0 2 4 6 5 10 15 20 25 30 35 40 45 50 55 N=40 x=40,3 haul. 46, 47, 48, 51, 52 0 2 4 6 8 10 12 14 16 5 10 15 20 25 30 35 40 45 50 55 N=54 x=46,2 haul. 45, 54, 55 0 2 4 6 8 10 5 10 15 20 25 30 35 40 45 50 55 N=65 x=34,3 haul. 34 0 2 4 5 10 15 20 25 30 35 40 45 50 55 N=20 x=44,1 haul. 37, 40, 43, 44 0 10 20 30 40 50 60 70 80 90 5 10 15 20 25 30 35 40 45 50 55 N=404 x=48,3haul. 19 - 26, 1 0 2 4 6 8 10 12 14 16 18 20 5 10 15 20 25 30 35 40 45 50 55 N=68 x=48,7 haul. 2, 3, 6 - 15
- 53. 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 55° 54° 30' 30' 30' 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 53° 55° 54° 30' 30' 30' 53° >1°C; <2°C >0°C; <1°C Temperature at depth 200 m N=676 Ps. georgianus 1-13.II.89 1 2 3 5 6 7 8 9 10 11 12 14 15 17 18 19 2021 22 23 24 252627 28 29 30 31 32 3436 37 38 39 4041 43 44 45 46 47 48 51 52 54 55 59P 60P 57P 58P wieku III, IV, V, VI. - 5 postlarv/h; 15 postlarvae/h of age group 0. - 10 fish/h; 15 101 fish/h; 330 fish/h of age, - 2 fish/h; 15 54 fish/h, of age group I. - 4 fish/h; 15 20 fish/h, of age group II. 0 2 4 6 5 10 15 20 25 30 35 40 45 50 55 N=40 x=40,3 haul. 46, 47, 48, 51, 52 0 2 4 6 8 10 12 14 16 5 10 15 20 25 30 35 40 45 50 55 N=54 x=46,2 haul. 45, 54, 55 0 2 4 6 8 10 5 10 15 20 25 30 35 40 45 50 55 N=65 x=34,3 haul. 34 0 2 4 5 10 15 20 25 30 35 40 45 50 55 N=20 x=44,1 haul. 37, 40, 43, 44 0 10 20 30 40 50 60 70 80 90 5 10 15 20 25 30 35 40 45 50 55 N=404 x=48,3haul. 19 - 26, 1 0 2 4 6 8 10 12 14 16 18 20 5 10 15 20 25 30 35 40 45 50 55 N=68 x=48,7 haul. 2, 3, 6 - 15
- 54. 0 10 20 30 40 50 60 1-10.II. 1989 45 cm TL, 808 g. 46 cm TL, 870 g. ~49 cm TL, 1275 g. ×105 fish 500 250 50 150 0 100 200 300 400 500 600 700 1-10.II. 1989 45 cm TL, 808 g. 46 cm TL, 870 g. 49 cm TL, 1275 g. Fish/km2 250 150 50 500 0 5 10 15 20 25 % TL, cm 50-150 m: n=71, TL=48,9 cm S.Georgia Is., 1-10.II.1989 0 5 10 15 % TL, cm 150-250 m: n=348, TL=46,3 cm S.Georgia Is., 1-10.II.1989 0 5 10 15 15 20 25 30 35 40 45 50 55 60 % TL, cm 250-500 m: n=252, TL=44,6 cm S.Georgia Is., 1-10.II.1989
- 55. 0 10 20 30 40 50 60 1-10.II. 1989 45 cm TL, 808 g. 46 cm TL, 870 g. ~49 cm TL, 1275 g. ×105 fish 500 250 50 150 0 100 200 300 400 500 600 700 1-10.II. 1989 45 cm TL, 808 g. 46 cm TL, 870 g. 49 cm TL, 1275 g. Fish/km2 250 150 50 500 0 5 10 15 20 25 % TL, cm 50-150 m: n=71, TL=48,9 cm S.Georgia Is., 1-10.II.1989 0 5 10 15 % TL, cm 150-250 m: n=348, TL=46,3 cm S.Georgia Is., 1-10.II.1989 0 5 10 15 15 20 25 30 35 40 45 50 55 60 % TL, cm 250-500 m: n=252, TL=44,6 cm S.Georgia Is., 1-10.II.1989
- 56. 1 1 0.5 0.25 11.25 1.5 1.75 2 2.2522.252.5 20 40 60 80 100 120 140 160 180 200 0 0.4 0.6 1 2 30.5 0.7 0.8 1.25 1.5 1.75 2.25 2.5 0.75 0.75 1500 1000 500 Krillbiomass[t/nm2] m Witek, 1988; Sahrhage, 1988
- 57. 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 55° 54° 30' 30' 30' 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 53° 55° 54° 30' 30' 30' 53° >0,6°C; <1°C Temperature at depth 200 m Ps. georgianus 06-26.I.90 0 5 10 15 20 25 30 35 40 6 11 16 21 26 31 36 41 46 51 56 23,6 cm N=125 zac: 19, 20, 23 0 5 10 15 20 6 11 16 21 26 31 36 41 46 51 56 x=26,4N=65 zac: 48, 49, 54, 57 0 20 40 60 80 6 11 16 21 26 31 36 41 46 51 56 x=23,6 N=219 zac: 76, 77, 78, 79 0 1 2 3 4 5 6 7 8 6 11 16 21 26 31 36 41 46 51 56 x=49,3 N=23haul. 42 0 1 2 3 4 6 11 16 21 26 31 36 41 46 51 56 x=35,7 N=22haul. 43 0 2 4 6 8 6 11 16 21 26 31 36 41 46 51 56 x=43,3 N=57zac: 33, 34, 35 0 2 4 6 8 10 12 14 6 11 16 21 26 31 36 41 46 51 56 x=48,5 N=48 zac: 60, 63 0 1 2 3 4 6 11 16 21 26 31 36 41 46 51 56 x=34,5 N=41 zac: 70, 71, 80 0 1 2 3 4 5 6 11 16 21 26 31 36 41 46 51 56 x=24,8 N=21 zac: 29, 31 0 2 4 6 8 10 12 6 11 16 21 26 31 36 41 46 51 56 x=45,5 N=73 zac: 45, 50 0 20 40 60 80 100 120 140 5 10 15 20 25 30 35 40 45 50 55 x=31,6N=856 haul.: 5-82 11 5 82 0 1 2 6 11 16 21 26 31 36 41 46 51 56 x=46,8 N=6zac: 5, 11, 82 70 28 53 29 26 30 25 79 77 76 71 63 60 54 50 48 45 44 43 42 36 31 18 19 23 39 38 37 13 16 17 20 22 32 34 51 15 80 66 65 59 57 56 52 49 4175 74 24 78 35 21 27 33 72 47 81 73 - 2; 28; 92 postlarvae/h of age group 0. - 6; 28; 90 fish/h of age group III - Δ, IV - ×, V. - 10; 73; 176 fish/h, of age group I. - 4; 19 fish/h, of age group II.
- 58. 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 55° 54° 30' 30' 30' 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 53° 55° 54° 30' 30' 30' 53° >0,6°C; <1°C Temperature at depth 200 m Ps. georgianus 06.I.90-26.I.90 0 5 10 15 20 25 30 35 40 6 11 16 21 26 31 36 41 46 51 56 23,6 cm N=125 zac: 19, 20, 23 0 5 10 15 20 6 11 16 21 26 31 36 41 46 51 56 x=26,4N=65 zac: 48, 49, 54, 57 0 20 40 60 80 6 11 16 21 26 31 36 41 46 51 56 x=23,6 N=219 zac: 76, 77, 78, 79 0 1 2 3 4 5 6 7 8 6 11 16 21 26 31 36 41 46 51 56 x=49,3 N=23haul. 42 0 1 2 3 4 6 11 16 21 26 31 36 41 46 51 56 x=35,7 N=22haul. 43 0 2 4 6 8 6 11 16 21 26 31 36 41 46 51 56 x=43,3 N=57zac: 33, 34, 35 0 2 4 6 8 10 12 14 6 11 16 21 26 31 36 41 46 51 56 x=48,5 N=48 zac: 60, 63 0 1 2 3 4 6 11 16 21 26 31 36 41 46 51 56 x=34,5 N=41 zac: 70, 71, 80 0 1 2 3 4 5 6 11 16 21 26 31 36 41 46 51 56 x=24,8 N=21 zac: 29, 31 0 2 4 6 8 10 12 6 11 16 21 26 31 36 41 46 51 56 x=45,5 N=73 zac: 45, 50 0 20 40 60 80 100 120 140 5 10 15 20 25 30 35 40 45 50 55 x=31,6N=856 haul.: 5-82 11 5 82 0 1 2 6 11 16 21 26 31 36 41 46 51 56 x=46,8 N=6zac: 5, 11, 82 70 28 53 29 26 30 25 79 77 76 71 63 60 54 50 48 45 44 43 42 36 31 18 19 23 39 38 37 13 16 17 20 22 32 34 51 15 80 66 65 59 57 56 52 49 4175 74 24 78 35 21 27 33 72 47 81 73 - 2; 28; 92 postlarvae/h of age group 0. - 6; 28; 90 fish/h of age group III - Δ, IV - ×, V. - 10; 73; 176 fish/h, of age group I. - 4; 19 fish/h, of age group II.
- 59. 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 55° 54° 30' 30' 30' 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 53° 55° 54° 30' 30' 30' 53° >1°C; <2°C >0.6°C; <1°CTemperature at depth 200 m 48 51 54 60 62 66 6972 75 79 83 85 88 91 99 101 105 108 113 116 121123 126 129 133 136 138141 145 147 151 154 156 159 162 165 169172 174 178 181 184 189193 195 198 201 204 208 211 214 217 220 223 226 229 232 235 241244 247 250 253 256 258 262265 268 270 272 275 280 282 3 7 10 13 16 21 24 27 31 34 38 41 45 0 2 4 6 8 6 11 16 21 26 31 36 41 46 51 56 n TL, cm n=34 x=33,5 zac: 189, 204 7; 32; 124 fish/h of age group IV, V. 8; 31; 72; 115; 221 fish/h z III. - 6; 20; 1 39; 61 fish/h, of age group I. 6; 22; 39 ; 61 fish/h, of age group II. 0 1 2 3 4 5 6 7 6 11 16 21 26 31 36 41 46 51 56 n TL, cm N=39 x=36,4zac.=48 S. Georgia I. 1992, Ps. georgianus, n=1878 0 10 20 30 40 50 60 70 6 11 16 21 26 31 36 41 46 51 56 n TL, cm N=486 x=37,5 zac: 241, 244, 250, 256 0 10 20 30 40 50 60 70 6 11 16 21 26 31 36 41 46 51 56 n TL, cm N=502 x=45,2 zac: 121, 129, 136, 138, 141 0 10 20 30 40 50 6 11 16 21 26 31 36 41 46 51 56 n TL, cm N=390 x=39,4 zac: 51, 66 - 91 - 95 - 101
- 60. 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 55° 54° 30' 30' 30' 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 53° 55° 54° 30' 30' 30' 53° >1°C; <2°C >0°C; <1°CTemperature at depth 200 m 48 51 54 60 62 66 6972 75 79 83 85 88 91 99 101 105 108 113 116 121123 126 129 133 136 138141 145 147 151 154 156 159 162 165 169172 174 178 181 184 189193 195 198 201 204 208 211 214 217 220 223 226 229 232 235 241244 247 250 253 256 258 262265 268 270 272 275 280 282 3 7 10 13 16 21 24 27 31 34 38 41 45 0 2 4 6 8 6 11 16 21 26 31 36 41 46 51 56 n TL, cm n=34 x=33,5 zac: 189, 204 7; 32; 124 fish/h of age group IV, V. 8; 31; 72; 115; 221 fish/h z III. - 6; 20; 1 39; 61 fish/h, of age group I. 6; 22; 39 ; 61 fish/h, of age group II. 0 1 2 3 4 5 6 7 6 11 16 21 26 31 36 41 46 51 56 n TL, cm N=39 x=36,4zac.=48 S. Georgia I. 1992, Ps. georgianus, n=1878 0 10 20 30 40 50 60 70 6 11 16 21 26 31 36 41 46 51 56 n TL, cm N=486 x=37,5 zac: 241, 244, 250, 256 0 10 20 30 40 50 60 70 6 11 16 21 26 31 36 41 46 51 56 n TL, cm N=502 x=45,2 zac: 121, 129, 136, 138, 141 0 10 20 30 40 50 6 11 16 21 26 31 36 41 46 51 56 n TL, cm N=390 x=39,4 zac: 51, 66 - 91 - 95 - 101
- 61. 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 55° 54° 30' 30' 30' 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 53° 55° 54° 30' 30' 30' 53° >1°C; <2°C >0°C; <1°C Temperature at depth 200 m N=676 Ps. georgianus 1.II.89-I.92 1 2 3 5 6 7 8 9 10 11 12 14 15 17 18 19 2021 22 23 24 252627 28 29 30 31 32 3436 37 38 39 4041 43 44 45 46 47 48 51 52 54 55 59P 60P 57P 58P wieku III, IV, V, VI. - 5 postlarvaes/h; 92 postlarvaes/h of age group 0. - 10 fish/h; 101 fish/h; 330 fish/h of age above III, - 2 fish/h; 15 176 fish/h, of age group I. - 4 fish/h; 15 20 fish/h, of age group II. 0 2 4 6 5 10 15 20 25 30 35 40 45 50 55 N=40 x=40,3 haul. 46, 47, 48, 51, 52 0 2 4 6 8 10 12 14 16 5 10 15 20 25 30 35 40 45 50 55 N=54 x=46,2 haul. 45, 54, 55 0 2 4 6 8 10 5 10 15 20 25 30 35 40 45 50 55 N=65 x=34,3 haul. 34 0 2 4 5 10 15 20 25 30 35 40 45 50 55 N=20 x=44,1 haul. 37, 40, 43, 44 0 10 20 30 40 50 60 70 80 90 5 10 15 20 25 30 35 40 45 50 55 N=404 x=48,3haul. 19 - 26, 1 0 2 4 6 8 10 12 14 16 18 20 5 10 15 20 25 30 35 40 45 50 55 N=68 x=48,7 haul. 2, 3, 6 - 15 7; 32; 124 fish/h of age group IV, V. 0 5 10 15 20 6 11 16 21 26 31 36 41 46 51 56 x=26,4N=65 haul 48, 49, 54, 57 0 1 2 3 4 5 6 7 8 6 11 16 21 26 31 36 41 46 51 56 x=49,3 N=23haul. 42 0 2 4 6 8 10 12 6 11 16 21 26 31 36 41 46 51 56 x=45,5 N=73 haul 45,50 0 20 40 60 80 6 11 16 21 26 31 36 41 46 51 56 x=23,6 N=219 haul 76, 77, 78, 79
- 62. 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 55° 54° 30' 30' 30' 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 53° 55° 54° 30' 30' 30' 53° >1°C; <2°C >0°C; <1°C Temperature at depth 200 m N=676 Ps. georgianus 1.II.89-I.92 1 2 3 5 6 7 8 9 10 11 12 14 15 17 18 19 2021 22 23 24 252627 28 29 30 31 32 3436 37 38 39 4041 43 44 45 46 47 48 51 52 54 55 59P 60P 57P 58P wieku III, IV, V, VI. - 5 postlarvaes/h; 92 postlarvaes/h of age group 0. - 10 fish/h; 101 fish/h; 330 fish/h of age above III, - 2 fish/h; 15 176 fish/h, of age group I. - 4 fish/h; 15 20 fish/h, of age group II. 0 2 4 6 5 10 15 20 25 30 35 40 45 50 55 N=40 x=40,3 haul. 46, 47, 48, 51, 52 0 2 4 6 8 10 12 14 16 5 10 15 20 25 30 35 40 45 50 55 N=54 x=46,2 haul. 45, 54, 55 0 2 4 6 8 10 5 10 15 20 25 30 35 40 45 50 55 N=65 x=34,3 haul. 34 0 2 4 5 10 15 20 25 30 35 40 45 50 55 N=20 x=44,1 haul. 37, 40, 43, 44 0 10 20 30 40 50 60 70 80 90 5 10 15 20 25 30 35 40 45 50 55 N=404 x=48,3haul. 19 - 26, 1 0 2 4 6 8 10 12 14 16 18 20 5 10 15 20 25 30 35 40 45 50 55 N=68 x=48,7 haul. 2, 3, 6 - 15 7; 32; 124 fish/h of age group IV, V. 0 5 10 15 20 6 11 16 21 26 31 36 41 46 51 56 x=26,4N=65 haul 48, 49, 54, 57 0 1 2 3 4 5 6 7 8 6 11 16 21 26 31 36 41 46 51 56 x=49,3 N=23haul. 42 0 2 4 6 8 10 12 6 11 16 21 26 31 36 41 46 51 56 x=45,5 N=73 haul 45,50 0 20 40 60 80 6 11 16 21 26 31 36 41 46 51 56 x=23,6 N=219 haul 76, 77, 78, 79
- 63. 63 Surface current <200 m Deep-sea current >200 m 40°W 38°W Sea currents pattern (Murphy, 2013)
- 64. 64 Surface current <200 m Deep-sea current >200 m 40°W 38°W
- 65. Surface current <200 m Deep-sea current >200 m 40°W 38°W On the north eastern side of the island large fish supposed to be well swimming, with using and opposing whirls (threatening to carry off from the shelf) could persist in them, and feed on accumulated in these whirls large specimens of krill. Separated geographically age groups were differentiated by the ability to swim in the currents and eddies needed to get food in them. This ability is managed by otolith shape. Why? As it is follow below
- 66. Surface current <200 m Deep-sea current >200 m 40°W 38°W On the north eastern side of the island large fish supposed to be well swimming, with using and opposing whirls (threatening to carry off from the shelf) could persist in them, and feed on accumulated in these whirls large specimens of krill. Separated geographically age groups were differentiated by the ability to swim in the currents and eddies needed to get food in them. This ability is managed by otolith shape. Why? As it is follow below:
- 67. 67 The efficiency of swimming in the currents increases the chances of survival on shelf. Success in obtaining food provides higher speed of swimming. To achieve that the flowing shapes of slightest resistance are created for all objects need to have high-speed of swimming to catch food, to escape, or to maintain themself in the habitat of currents. 2 2 1 2 )(, SvCR xha S ρ thanks to have it, fish mastered ocean space. This has also a reference to the otoliths provide in swimming balance and precision - a sense of balance needed for each free living organism even for simplest metazoans. The flattened shape of the otoliths poses little resistance in endolymph and increases the perception of positions in fast swimming. Indicating deviations of otoliths with flowing shape are 8 times less distorted than with spherical by the endolymph turbulences. In higher speeds flattening must be increased. Hydrodynamic resistance Ra,h is the smallest for flowing shape = 1 Anon, 2006
- 68. 68 Fish to catch other fish must be bigger and stronger and have body shape with little resistance. Escaping fish are elongated to have smaller body resistance Photo Gallery by Alex Robinson
- 69. 69 The efficiency of swimming in the currents increases the chances of survival on shelf. Success in obtaining food provides higher speed of swimming. To achieve that the flowing shapes of slightest resistance are created for all objects need to have high-speed of swimming to catch food, to escape, or to maintain themself in the habitat of currents. 2 2 1 2 )(, SvCR xha S ρ thanks to have it, fish mastered ocean space. This has also a reference to the otoliths provide in swimming balance and precision. The flattened shape of the otoliths poses little resistance in endolymph and increases the perception of positions in fast swimming. Hydrodynamic resistance Ra,h is the smallest for flowing shape = 1
- 70. 70 When catching body fish is bent in an arc, pelvic fins as stabilizers, pectoral fins on the body to reduce resistance Photo Gallery by Alex Robinson
- 71. 71 Swimming development in fish. asc, psc, lsc – anterior, posterior, lateral semicircular canals, c – cristae, l – lagena, ml, ms, mu – macula lagenae, sacculi, utriculi, s – saccule, u – utriculi, ed – endolymphatic duct, c – cochlea, bm – basilar membrane, pb – papilla basilaris. sensorial microvilli Vestibular nerve fibers Otoliths have changes of shape from spherical to streamline elliptical to more elongated shape as the speed of fish swimming increase, which takes place during the ontogeny and phylogeny. The surface of the otoliths is plastically formed by labyrinth and by measured the changes of endolymphatic pressure induced by activity. Changes of the pressure in the endolymph arrange her ingredients from which at start of that changes in stationary larvae they assembling into spherical otoliths. 21 cm TL ml l c psc asc ms s u mu lsc ms drift swimming slow swimming stationary fast swimming ms ~0.3 km/h~0.1 km/h~0.01 km/h
- 72. Spherical otolith of larvae during development of their tissues, by the first movement of the body (by energetic bending of the body on sides) change its shape on ellipsoid flattened on sides correspondingly to the increased pressure of the endolymph on these sides. These fish swim with movements of the pectoral fins, relatively slow, so flattening their otoliths are small. 72 21 cm TL
- 73. 73 After hatching otoliths shape becomes more flattened from 2 to even 3 times. It is because hatched larvae after get off from the egg obtain large free space for physical activity and for swimming: from inside the eggs to the space of coastal waters, for example of Cumberland Gulf, where the larvae of Ps. georgianus were found. 0 .4 0 .2 0 .2 0 .6 0 .4 0 .4 0.2 0.2 0.4 0.6 0 .06 0.04 0.02 0 0.02 0.04 37.3 1 1.2 1 Traczyk, 2013
- 74. 74 LN, seperated larval microstrukture easy disamble by EDTA from other parts of otolith Changing environment and physiology in hatching is as large as it is clearly marked in the microstructure. For Ps. georgianus the mark of that shift is as wide layer having more calcium that washed out separates the hatching nucleus from the rest of the otolith. Traczyk, 2013
- 75. 75 LN, seperated larval microstrukture easy disamble by EDTA from other parts of otolith
- 77. 0.6 0 .4 0.2 0 0.2 0.4 0.6 Further flattening of otoliths runs from postlarvae to fish of I age group (from 2.8 to 3). Postlarvae swims faster than larvae as begins to migrate to the waters further from shore and deeper, their otoliths become more flattered. Postlarvae drifting in coastal current accumulate on west side of island. 1.20.80.400.40.81.21.6 8.2 1 0.3 1 SP LN 1mm 8.2 1 AP
- 78. The increase in the otolith flattening by Second Primordium on medial cross- section give the widest surface which in move of fish in dorsal direction increase the perception of balance during vertical migration 0.6 0 .4 0.2 0 0.2 0.4 0.6 1.20.80.400.40.81.21.6 8.2 1 0.3 1 SP LN 1mm 8.2 1
- 79. Second Primordium increase the flattening of otolith and decrease its front profile SEM 2nmplatinum+palladium SecondPrimordium 0.1 mm 8.2 1 0.3 1 Traczyk, 2013
- 80. 0 1 2 3 4 5 6 7 8 9 10 mm Inside egg inshore outshore deep water, below 200 m development Age group : 0 I II III IV V VI Traczyk, 2012; Traczyk, 2013
- 81. 0 1 2 3 4 5 6 7 8 9 10 mm Inside egg inshore outshore deep water, below 200 m development Age group : 0 I II III IV V VI
- 82. 0 1 2 3 4 5 6 7 8 9 10 mm Age group : 0 I II III IV V VI~0.3 km/h ~0.9 km/h
- 83. TL = 4,2463e0,3834·OH R² = 0,9819 TL = 3,3334e0,5477·OL R² = 0,9621 TL = 10,632·R9 1,0356 R² = 0,9805 TL = 9,382·ORL - 4,2732 R² = 0,9543 0 10 20 30 40 50 60 0 1 2 3 4 5 6 7 OH – height of adult otolith OLJ – otolith length of juvenes OL – otolith length ORL – otolith rostral length R9 – dorsal radius TL, cm mm Results are in age groups OH = 1,2623·OLJ - 0,2123 R² = 0,9277 OH= 0,8248·OL1,3048 R² = 0,9116 OH = 0,8064·ORL + 1,9359 R² = 0,9382 0 1 2 3 4 5 6 7 8 0 1 2 3 4 5 6 7 OL, mm OH,mm OH – otolith height of adults OLJ – otolith lenght of juvenes ORL – otolith rostral length OH > OL: OH=b·OL+a above y=x proportionality of the otolith dimensions are constant Anterior culliculum
- 84. OH = 1,2623·OLJ - 0,2123 R² = 0,9277 OH= 0,8248·OL1,3048 R² = 0,9116 OH = 0,8064·ORL + 1,9359 R² = 0,9382 0 1 2 3 4 5 6 7 8 0 1 2 3 4 5 6 7 OL, mm OH,mm OH – otolith height of adults OLJ – otolith lenght of juvenes ORL – otolith rostral length TL = 4,2463e0,3834·OH R² = 0,9819 TL = 3,3334e0,5477·OL R² = 0,9621 TL = 10,632·R9 1,0356 R² = 0,9805 TL = 9,382·ORL - 4,2732 R² = 0,9543 0 10 20 30 40 50 60 0 1 2 3 4 5 6 7 OH – height of adult otolith OLJ – otolith length of juvenes OL – otolith length ORL – otolith rostral length R9 – dorsal radius TL, cm mm Results are in age groups OH > OL: OH=b·OL+a above y=x proportionality of the otolith dimensions are constant Anterior culliculum
- 85. TL = 4,2463e0,3834·OH R² = 0,9819 TL = 3,3334e0,5477·OL R² = 0,9621 TL = 10,632·R9 1,0356 R² = 0,9805 TL = 9,382·ORL - 4,2732 R² = 0,9543 0 10 20 30 40 50 60 0 1 2 3 4 5 6 7 OH – height of adult otolith OLJ – otolith length of juvenes OL – otolith length ORL – otolith rostral length R9 – dorsal radius TL, cm mm Results are in age groups OH = 1,2623·OLJ - 0,2123 R² = 0,9277 OH= 0,8248·OL1,3048 R² = 0,9116 OH = 0,8064·ORL + 1,9359 R² = 0,9382 0 1 2 3 4 5 6 7 8 0 1 2 3 4 5 6 7 OL, mm OH,mm OH – otolith height of adults OLJ – otolith lenght of juvenes ORL – otolith rostral length OH > OL: OH=b·OL+a above y=x proportionality of the otolith dimensions are constant Anterior culliculum
- 86. How do we know that the shape of otolith indicates the speed of swimming? In comparison: faster species have them more flatter S. japonicus Ps. georgianus faster species Transverse plane median plane median plane Transverse plane 1 mm1 mm Data for speed of swimming (Żbikowski, 2008, Fuiman, 2002) 21.3 km/h ~1,6 km/h
- 87. the shape of otolith is chageing among species of fish of different depth of living Channichthyidae Ch. aceratus Macrouridae M. holotrachysDeep water species shelves species Transverse plane Transverse plane 1 mm 1 mm Body fish and otolith shape data (Hecht, 1987; Fischer, 1985; Grabowska, 2010; Traczyk, 1992)
- 88. 0 0.0002 0.0004 0.0006 0.0008 0.001 0.0012 0.0014 0.0016 0 5 10 15 20 25 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1 1.1 1.2 1.3 1.4 1.5 1.6 1.7 1.8 1.9 2 2.1 2.2 2.3 R3 mm mm Age groups = R3/ y/ 365 = 2,26 / 0,000775 / 365 = 8 y = 0,000775 mm mmY 000775,0 mmY 000276,0 1 mmS. japonicus, FL=39 cm Mauretania 21.I.2011 Haul 44. No 39 opticaldensityannualincrements 0 0.0001 0.0002 0.0003 0.0004 0.0005 0.0006 0 10 20 30 40 50 60 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 R11 mmAge = R11/ y / 365 = 0,81 / 0,000276 / 365 = 8 y = 0,000276 mm Large changes in the width increments of otolith mackerel on radii R3, R11 0 0.5 1 1.5 2 2.5 1,8 cm FL; 0,011 mm 4,2 cm; 0,00251 mm 14,6 cm; 0,00215 mm 21,7 cm; 0,00122 mm 26,4 cm; 0,00079 mm 29,8 cm; 0,00059 mm 33,1 cm; 0,00056 mm 35,3 cm; 0,00038 mm 38 cm; 0,00046 mm 40,7 cm; 0,00045 mm 42,5 cm; 0,00031 mm Back edge Ageannuli Spawn from January to May Fork Length FL; daily increments width from inner side (lack from outer) Nucleus 0 2 4 6 8 10 dailyincrementswidth R3 R11 otolith shape data (Traczyk, 2011)
- 89. 0 0.0002 0.0004 0.0006 0.0008 0.001 0.0012 0.0014 0.0016 0 5 10 15 20 25 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1 1.1 1.2 1.3 1.4 1.5 1.6 1.7 1.8 1.9 2 2.1 2.2 2.3 R3 mm mm Age groups = R3/ y/ 365 = 2,26 / 0,000775 / 365 = 8 y = 0,000775 mm mmY 000775,0 mmY 000276,0 1 mmS. japonicus, FL=39 cm Mauretania 21.I.2011 Haul 44. No 39 opticaldensityannualincrements 0 0.0001 0.0002 0.0003 0.0004 0.0005 0.0006 0 10 20 30 40 50 60 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 R11 mmAge = R11/ y / 365 = 0,81 / 0,000276 / 365 = 8 y = 0,000276 mm Otolith mackerel on R3 has large width increments, on R11 small, so has larger length than high. 0 0.5 1 1.5 2 2.5 1,8 cm FL; 0,011 mm 4,2 cm; 0,00251 mm 14,6 cm; 0,00215 mm 21,7 cm; 0,00122 mm 26,4 cm; 0,00079 mm 29,8 cm; 0,00059 mm 33,1 cm; 0,00056 mm 35,3 cm; 0,00038 mm 38 cm; 0,00046 mm 40,7 cm; 0,00045 mm 42,5 cm; 0,00031 mm Back edge Ageannuli Spawn from January to May Fork Length FL; daily increments width from inner side (lack from outer) Nucleus 0 2 4 6 8 10 dailyincrementswidth R3 R11
- 90. Extreme length of otolith compensated by reduction of it other sides because of S. japonicus spawn I-V 0 0.5 1 1.5 2 2.5 1,8 cm FL; 0,011 mm 4,2 cm; 0,00251 mm 14,6 cm; 0,00215 mm 21,7 cm; 0,00122 mm 26,4 cm; 0,00079 mm 29,8 cm; 0,00059 mm 33,1 cm; 0,00056 mm 35,3 cm; 0,00038 mm 38 cm; 0,00046 mm 40,7 cm; 0,00045 mm 42,5 cm; 0,00031 mm Back edge Ageannuli nucleus 0 2 4 6 8 10 Direction of lack of otolith growth Fork Length FL; daily increments width from inner side Otolith back radius (mm) high speed of Extreme length of mackerel otoliths arise from high speed of swimming (show by torpedo shape of body) - from their incessant fast swimming in the pelagic ocean (to not to allow to fall, from their body heavier than water). Increase in the dorsal edge of mackerel otolith is reduced, because this fish has stability resulting from high inertia, or from high frictional force.
- 91. Extreme length of otolith compensated by reduction of it other sides because of Lack of the increments on the one side of the labyrinth is probably from a pressure of swimming speed removing otoliths substrates from that side to the other, where a large acceleration locally concentrate them thus determining the constant rapid increase of the otolith length and their local elongation at the expense of not growing of the otolith inner surface. In contrast Ps. georgianus is slower swimmer but migrates vertically with perfection ensured by larger high of the otoliths. S. japonicus spawn I-V 0 0.5 1 1.5 2 2.5 1,8 cm FL; 0,011 mm 4,2 cm; 0,00251 mm 14,6 cm; 0,00215 mm 21,7 cm; 0,00122 mm 26,4 cm; 0,00079 mm 29,8 cm; 0,00059 mm 33,1 cm; 0,00056 mm 35,3 cm; 0,00038 mm 38 cm; 0,00046 mm 40,7 cm; 0,00045 mm 42,5 cm; 0,00031 mm Back edge Ageannuli nucleus 0 2 4 6 8 10 Direction of lack of otolith growth Fork Length FL; daily increments width from inner side Otolith back radius (mm) high speed of
- 92. S. japonicus Ps. georgianus 1 mm Speed in swimming is the source of shape diversity Dorsal margin nucleus Annual increments Outer dorsal side Transverse plane one side increments 1 mm nucleus Changes in the width increments on R3, R11 mmY 000775,0 mmY 000276,0 rostrum nucleus
- 93. 1 mm Dorsal edge of mackerel otoliths have increments tightened up and the growth radius becomes 3.7 and 5 times smaller than the radii of growth to back and front edges. In Ps. georgianus otolith radii growth in opposite pattern, wide microincrements form dorsal radius of 1.8 and 1.5 times larger than the radii of the back and front edges. Ps. georgianus rostrum Transverse plane one side increments Changes in the width increments on R3, R11 nucleus mmY 000775,0 mmY 000276,0 nucleus Outer dorsal side
- 94. 1 mm The high otoliths of SGI icefish - as fishing floats inform about vertical stability needed for vertical migrations and for lifting with currents. In opposite to that long otoliths of mackerel are sensitive on changes during swimming in the horizontal direction. Information important in the fast swimming for far distances. Ps. georgianus rostrum Transverse plane one side increments Changes in the width increments on R3, R11 nucleus mmY 000775,0 mmY 000276,0 nucleus Outer dorsal side
- 95. 95 Swimming depth is source of diversity in microstructure and shape of the otolith Transverse plane Otolith shape data Grabowska, 2010
- 96. 96 Transverse section similar to Channichthyidae, but otolith length is large similar to length of otolith mackerel: ~2×> otolith height: R >>R , M. carinatus live in deep waters 1200m 0 50 100 150 200 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1 1.1 1.2 1.3 1.4 1.5 1.6 1.7 1.8 mm High depth of swimming is the source of shape diversity
- 97. 97 C. aceratus M. carinatus M. carinatus has longer OL than C. aceratus, but is not faster - it swim deeper dimensions of radii: small dorsal and large ventral for M. carinatus and vice wersa for Channichthyidae inversed proportions
- 98. 98 2 directional growth in otolith shape: determined by pulsed swimming R9=0,5 mm/0,007 = 72 days = 2,4 months R9=1,21 mm/0,007 = 160 days = 5,3 months Squids swims slow with pulsation have twins hemispheres in otolith shape with the widest otolith’ increments 70 90 110 130 150 170 190 210 230 250 270 0 0.05 0.1 0.15 0.2 0.25 0.3 0.35 0.4 0.45 0.5 0.55 0.6 0.65 mm displacement segment density profile with about 7 measurements, ie 0.0069 mm of density segment profile from a distance of 0.05 to 0.65 mm from the center of otolith. y = 162.08x + 74610 R² = 0.927 0.05 0.052 0.054 0.056 0.058 0.06 0.062 0.064 0.066 0.068 75000 75200 75400 75600 75800 76000 76200 76400 76600 76800 77000 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 Average daily increment width = difference between adjacent minima or maxima = 0.0069 mm, 0.0079 mm age = otolith radius / average width of daily increments = 0.68 mm / 0.0079 mm = 86 days = 3 months mm Data: Arkhipkin, 1996; Arkhipkin, 1999
- 99. Otolith shape differentiates pattern of high energy swimming of mackerel Scombrus japonicus ~21.3 km/h Squids 1-3 km/h pulsed swimming in squids Żbikowski, 2008; Videler J.J., 1984 Gosline, 1985
- 100. low energy swimming of icefish Channichthyidae using the pectoral fins Large pectoral fins are floating fish in the depths; vertical movements, are measured more by deviation of otolith height from the vertical, that is, by higher otolith. High, laterally flattened body having a great fins has about 20 times more resistance of the lateral than the front and the current pressure on the concave side of curved body of flowing fish produces a hydrodynamic force increasing speed of fish swimming forward. An asymmetrical shape with respect to the direction axis of swimming causes asymmetric flow, that creates differential pressure on opposite surfaces, and thus the driving force to forward. 1 mm (Fuiman L, 2002; Anon, 2006)
- 101. 101 High body and wide, long fins reduce drift in currents. Fin fish and flat body during swimming provide hydrodynamic lifting force. The shape of the otoliths is plastically formed by distribution of endolymph pressure thank to that it provides for fish from otolith shape interpretation - the information about the speed and body movement and also provides information about sound and water vibrations causing vibration of endolymph and giving seeing yourself in surroundings. Similarly, by evolution the body shape is formed in order to obtain maximum speed to which the shape of otolith is adjusted. Thanks to this otolith shape and body shape are interrelated. Body shape of Ps. georgianus evolved as the shape of otoliths with respect to the same target is high, facilitating vertical migration and is laterally flattened facilitating swimming in currents with minimal energy consumption.
- 102. 102 High body and wide, long fins reduce drift in currents. Fin fish and flat body during swimming provide hydrodynamic lifting force. Body shape as an indicator of the shape of otolith, because it results from the speed of swimming and life strategy and that all depends on body shape adapted to environmental conditions.
- 103. 103 Lifting strategy, the use of currents and the uplift force of fins.migration
- 104. 104 Lifting strategy, low-energy swimming migration (Le François, 2014; 2014a; PolarTrec, 2013; Detrich, 2012; Uve, 2008; Byrd, 2012)(Walesby, 1982; Davison W., 1985; HARRISON, 1987; Twelves, 1972) (Żabrowski, 2000)
- 105. In lifting strategy and low-energy swimming of Ps. georgianus pectoral fins are moving their first rays as spars entailing the sheet of streamer. Forward with a minimum resistance of sheets fins flowing after trace of thin first ray and back with a large opposition of all returning fin surface. Pectoral fins in the first phase of motion, horizontal spreading out to the front and to the sides increase the horizontal plane of fish so keep, support fish to float at required depth level. In the second phase the fins retracted horizontally to the rear are pushing its all surface on water and pushing fish forward. Also locomotor activity have a caudal fin but much smaller. Fin is bent on sideways with the body when fish is turning. Le François, 2014
- 106. Le François, 2014
- 107. Le François, 2014
- 108. Le François, 2014
- 109. Le François, 2014
- 110. Le François, 2014
- 111. Le François, 2014
- 128. Le François, 2014
- 129. The water pressure creates hydrodynamic force acts on side of the flowing fish. V – speed of the fish, Rh,c – front resist Rh,b – side resist = 22 V FP FC FAE Rh,c = 1,5 FAE – aero-hydrodynamic force - the force exerted on the body by the environment, which is the result of movement of the body relative to the environment (gas or liquid). FC – driving force, thrust (force of pressure induced by pressure of current exerted on the body surface area). Operates forward because of body shape an the resistance of the lateral is 20 times greater than the frontal; FP - drift force; viscosity force (friction at the surface of the body). Anon, 2006
- 130. An asymmetrical shape with respect to the axis of swimming direction causes asymmetric flow that creates differential pressure on opposite surfaces, and thus the driving force to forward. V – speed of the fish, Rh,c – front resist Rh,b – side resist = 22 V FP FC FAE Rh,c = 1,5 FAE – aero-hydrodynamic force - the force exerted on the body by the environment, which is the result of movement of the body relative to the environment (gas or liquid). FC – driving force, thrust (force of pressure induced by pressure of current exerted on the body surface area). Operates forward because of body shape an the resistance of the lateral is 20 times greater than the frontal; FP - drift force; viscosity force (friction at the surface of the body). Anon, 2006
- 131. Factors increasing the hydrodynamic force acting on back of the body. Force: Fa,h = kv2; power: Pa,h = kv3; 2×Vcurrent → 4×FAE FAE – aero hydrodynamic force increases on larger body of fish. Larger, stronger ones are occurring closer to the sea surface, where the currents are stronger with turbulences and eddys. Smaller fish so weaker live deeper where the currents are weaker and also in regions with weak currents, V FP FC 2×FAE Rh,c = 1,5 Icefish have adaptation to cold water. One of them Ps. georgianus live and choose habitat of sea currents – so to exist in it, it adopt the shape of the body, fins and otoliths in liftting strategy of low-energy swimming Anon, 2006
- 132. 132 The smooth surface of the body increases the power of aero-hydrodynamic FA, H Channichthyidae have a smooth skin, without scales, allowing the feeling of each particle of the water flowing and gliding over the surface of the skin and react accordingly by deflection of the body, or by rearrangement positions of fins to reduce the resistance, to increase laminar flow and to eliminate turbulences. Lack of scales could be adopted as an adaptation of a low-energy swimming in cold strong currents, for which in a warm water there is high energy swimming. For example. Salmon, trout, or mackerel. We can find that the lack of scales for icefish is treat as an adaptation to increase the respiration of skin. Jakobowski however, argues that such a view is wrong, because the scales are below the epidermis to which oxygen diffuses and therefore scales do not interfere with the diffusion of oxygen through the skin. Certainly scaleless increase skin smoothness.Jakubowski, 1971, 1982
- 133. The sensitivity and skin elasticity in the perception of the body bending V FP FC FAE Rh,c = 1,5 When the stream of water on the side of after current detach and move disordered (turbulent), this will reduce the hydrodynamic forces. The bending body must always be tailored to the nature of the currents. Too big bow causes break away water streams from the surface of the body, for small bow quite similar paths and velocity of water particles on both sides of the odd fins causing a lack of hydrodynamic forces.
- 134. The fins increase the smoothness and flow velocity of after current side of the body FAE V Rh,c = 1,5 After the first front dorsal fin and before the second dorsal fin creates the nozzle that accelerates water flow on after currant side of second fin and body. Body shape as an indicator of the shape of otolith, because it results from the speed of swimming and life strategy and that all depends on body shape adapted to environmental conditions. This could be show by compare of species.
- 135. 135 Larger lateral surface of the body increases the FAE strength: body highest, large head and jaws defines predator creates an arrow, pelvic fins large effective for vertical migration. Longer, unpaired fins increase body side resistance . body less high, but fins: anal and dorsal longer , smaller head larger pectoral fins so larger horizontal migrations Body less high, but very large anal fin, dorsal and pectoral, so the greatest horizontal migrations. The smallest head reduces front resisting when swimming.Data: Fisher, 1985
- 136. 136 Environmental requirements with respect to efficiency of swimming High body helps in swimming using shelf currents and countercurrents and vertical migration. Big mouth helps predation. Intermediate species is not as high as Ps. georgianus so has greater diffusion and slim as C. gunnari, so has less diffusion than it The most slender body sacrifice the species for predators, but it gives little front resistance, with the big fins giving the greatest diffusion.Data: Fisher, 1985
- 137. Shape of otoliths (determined by microstructure) can show the process and direction of growth of the body, which is a response to factors of surrounding marine environment. Increase of otoliths taking place outside the cell in endolymph suffers from it the same factors of the marine environment reaching endolymph through the bones of the body. Therefore, the body and otoliths become a models of the fish growth – as reader of suffered environmental influences - to which fish during growth is adapting the otolith shape by changes of microstructure of the otoliths, that are treat as indicators of fish behavior. y = 0.2251x - 0.0242 R² = 0.9798 y = 0.3985x0.9976 R² = 0.9971 0 2 4 6 8 10 12 14 16 18 20 0 10 20 30 40 50 60 15-30% SL SL, cm cm 0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40 0 10 20 30 40 50 60 C. aceratus C. gunnari C. aceratus C. gunnari C. aceratus C. gunnari Ps. georgianus SL, cm N Ps. georgianus Ps. georgianus has greater body height and shorter lengths than the C. gunnari and C. aceratus – also have shorter dorsal and anal fins in favor of head size and the decline of swimming opportunities. Otoliths of Ps. georgianus like its body are high. Data: Fisher, 1985; Traczyk, 2013, Parkes, 1990
- 138. Realization various opportunities of swimming arising from various constructions of body that are adapted to the best use of different habitats of environment allows the perception of this swimming by otolith recording it with appropriate shape. y = 0.8064x + 1.9359 R² = 0.9382 y = 0.9743x0.9426 R² = 0.991 y = 1.0484x1.1338 R² = 0.9877 0 1 2 3 4 5 6 7 8 0 1 2 3 4 5 6 7 OL, mm OH, mm OH Ps. georgianus ORL Ps. georgianus OH C. aceratus y = 7.3835e0.424x R² = 0.9718 y = 6.7436e0.4989x R² = 0.9909 y = 6.6517e0.9547x R² = 0.9889 0 10 20 30 40 50 60 70 0 1 2 3 4 5 6 7 ~OL mm TL, cm OH Ps. georgianus OL Ps. georgianus R9 Ps. georgianus OH C. aceratus OL C. aceratus R9 C. aceratus C. aceratus Otoliths of Ps. georgianus and C. aceratus as species are similar, have same shapes similar (~OL), but instead of that have important differences. In otoliths of C. aceratus increments are narrower and proportion: length with respect to height is reversed. Ps. georgianus are smaller, TL Ps. georgianus Traczyk, 2013; Traczyk, 1992; Fischer, 1985
- 139. 139 Ps. georgianus has a smaller range of occurrence but higher vertical migration than C. aceratus C. aceratus have longer otoliths and has a greater range of occurrence than Ps. georgianus. Ps. georgianus, otolith height OH> otolith length OL, TL body length C. aceratus, OH< OL, TL Data: Hecht, 1978
- 140. .)) 2 sin(( 9 1 constx T Ay i i i i R9=2,35mm 1590 days Ch. aceratus, 45cm SL S. Georgia ,29.III.1979 hol 136, No 75 OW=0,0247 g OH=3,44 mm SP AP Additional centers, AP are also available in otoliths of C. aceratus.. They give however a lower elongation than the radius R9 of Ps. georgianus. Dorsal edge for otolith of older fish of Ps. georgianus grows more strongly than in otoliths of C. aceratus.. Otoliths of greater length than height indicate a greater range and speed of swimming. This confirms the elongated shape of the body with less weight and with longer dorsal and anal fins by about 10 rays. As otoliths of C. aceratus are not high, so height of their body is reduced. Data: Traczyk, 1992; 2014
- 141. 141 Ps. georgianus has smaller range of occurrence than the C. gunnari C. gunnari: OH < OL Ps. georgianus, otolith height OH> otolith length OL, TL body length Data: Hecht, 1978
- 142. 142 180 1,5 2,5 y = 8.6308x1.2495 R² = 0.9848 y = 9.5723x1.2236 R² = 0.9843 y = 21.245x1.2292 R² = 0.9868 0 10 20 30 40 50 60 0 1 2 3 4 5 6 7 TL, cm mm OH Ps. georgianus OL Ps. georgianus R9 Ps. georgianus OH C. gunnari OL C. gunnari R9 C. gunnari 6,4 15,3 24 27,8 34 35 45 50 y = 0.92x1.0199 R² = 0.9979 y = 0.4834x1.0103 R² = 0.9859 0 0.5 1 1.5 2 2.5 3 3.5 4 0 0.5 1 1.5 2 2.5 3 3.5 4 OL > OH 1,07 : 1 mm OL, otolith length [mm] TL, cm Otoliths C. gunnari are longer than height, indicating a wider occurrence and greater speed of swimming. It confirms the elongated body with a lower height. Otoliths C. gunnari nearly square, two times smaller than otoliths Ps. georgianus. Data: Hecht, 1978;Traczyk, 2013; 2014
- 143. 0 10 20 30 40 50 60 70 80 90 100 110 120 130 140 150 160 170 180 190 200 0.1 0.11 0.12 0.13 0.14 0.15 0.16 0.17 0.18 0.19 0.2 mm A₉=18,26, φ₉=-9,64, T₉=-0,07012; ; stała =85; ∑(yₑ-yᵢ)²=330042 A₁=31,08, φ₁=1, T₁=0,0263; A₂=3,88, φ₂=-2,5, T₂=0,00417; A₃=9,18, φ₃=-3,5, T₃=0,01566; A₄=9,09, φ₄=3,57, T₄=-0,00812; A₅=0,07, φ₅=3,19, T₅=-0,00076; A₆=4,12, φ₆=-3,27, T₆=0,005; A₇=3,33, φ₇=-6,78, T₇=0,0021; A₈=9,31, φ₈=-7,28, T₈=0,00577; power ~A2 85)78,6 0021,0 2 sin(33,37 xy constx T Ay i ii i ) 2 sin( 9 1 2 Data: Traczyk, 2013; 2013
- 144. R9=2,35 mm 1590 days C. aceratus, 45 cm SL S. Georgia , 29.III.1979 hol. 136, s . 75 OW=0,0247 g OH= =3,44 mm AP SP The otoliths shape of larvae is similar to an oval on median plane and flattened on the transverse plane to reduce resistance. The biggest flattened otolith has C. gunnari so it swims the fastest and farthest. Older fish swim faster, so flattening of its otoliths increases. Data: Traczyk, 2013
- 145. R9=2,35 mm 1590 days C. aceratus, 45 cm SL S. Georgia , 29.III.1979 hol. 136, s . 75 OW=0,0247 g OH= =3,44 mm AP SP Larvae otoliths shape is similar to an oval on median plane and flattened on the transverse plane to reduce resistance. The biggest flattened otolith has C. gunnari so it swims the fastest and farthest. Older fish swim faster, so flattening of its otoliths increases. Data: Traczyk, 2012
- 146. 0 1 2 3 4 5 6 7 8 9 1 1.5 2 2.5 3 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 mackerel SL, cm flattening y = 0.1585x + 1.7153 y = 0.0339x + 1.0898 y = 0.2521x + 5.2326 0 1 2 3 4 5 6 7 8 9 1 1.5 2 2.5 3 0 10 20 30 40 50 Scaleformackerel OH/OT OH – otolith height OT – otolith thick OH/OT SL, cm Data: Traczyk, 2012
- 147. 147 0 1 2 3 4 5 6 7 8 9 1 1.5 2 2.5 3 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 mackerel SL, cm flattening y = 0.1585x + 1.7153 y = 0.0339x + 1.0898 y = 0.2521x + 5.2326 0 1 2 3 4 5 6 7 8 9 1 1.5 2 2.5 3 0 10 20 30 40 50 Scaleformackerel OH/OT OH – otolith height OT – otolith thick OH/OT SL, cm
- 148. Shag Rock S.Georgia I. S.Sandwich I. S. Orkney I. Elephan I. K.GeorgeI. Deception Palmer A. Balleny Kerguelen I. Bouvet I. Heard I. Ch. aceratus: 5-770 m; 53°S-65°S Ch. gunnari : 0-700 m; 48°S-66°S Unusual catch location of Ps. georgianus Balleny I. Russia 2004/05 S. Sandwich I. Germany 1975/76; 1980/81 Kerguelen I. Australia 2003/04 Known catch location of fish: Ps. georgianus: 0-475 m; 53°S-66°S Data: Fischer, 1985; CCAMLR, 2012; Traczyk, 2013
- 149. Shag Rock S.Georgia I. S.Sandwich I. S. Orkney I. Elephan I. K.GeorgeI. Deception Palmer A. Balleny Kerguelen I. Bouvet I. Heard I. Ch. aceratus: 5-770 m; 53°S-65°S Ch. gunnari : 0-700 m; 48°S-66°S Unusual catch location of Ps. georgianus Balleny I. Russia 2004/05 S. Sandwich I. Germany 1975/76; 1980/81 Kerguelen I. Australia 2003/04 Known catch location of fish: Ps. georgianus: 0-475 m; 53°S-66°S
- 150. Shag Rock S.Georgia I. S.Sandwich I. S. Orkney I. Elephan I. K.GeorgeI. Deception Palmer A. Balleny Kerguelen I. Bouvet I. Heard I. Ch. aceratus: 5-770 m; 53°S-65°S Ch. gunnari : 0-700 m; 48°S-66°S Unusual catch location of Ps. georgianus Balleny I. Russia 2004/05 S. Sandwich I. Germany 1975/76; 1980/81 Kerguelen I. Australia 2003/04 Known catch location of fish: Ps. georgianus: 0-475 m; 53°S-66°S ?
- 151. Shag Rock S.Georgia I. S.Sandwich I. S. Orkney I. Elephan I. K.GeorgeI. Deception Palmer A. Balleny Kerguelen I. Bouvet I. Heard I. Ch. aceratus: 5-770 m; 53°S-65°S Ch. gunnari : 0-700 m; 48°S-66°S Unusual catch location of Ps. georgianus Balleny I. Russia 2004/05 S. Sandwich I. Germany 1975/76; 1980/81 Kerguelen I. Australia 2003/04 Known catch location of fish: Ps. georgianus: 0-475 m; 53°S-66°S ?
- 152. Shag Rock S.Georgia I. S.Sandwich I. S. Orkney I. Elephan I. K.GeorgeI. Deception Palmer A. Balleny Kerguelen I. Bouvet I. Heard I. Ch. aceratus: 5-770 m; 53°S-65°S Ch. gunnari : 0-700 m; 48°S-66°S Unusual catch location of Ps. georgianus Balleny I. Russia 2004/05 S. Sandwich I. Germany 1975/76; 1980/81 Kerguelen I. Australia 2003/04 Known catch location of fish: Ps. georgianus: 0-475 m; 53°S-66°S ?
- 153. Shag Rock S.Georgia I. S.Sandwich I. S. Orkney I. Elephan I. K.GeorgeI. Deception Palmer A. Balleny Kerguelen I. Bouvet I. Heard I. Ch. aceratus: 5-770 m; 53°S-65°S Ch. gunnari : 0-700 m; 48°S-66°S Unusual catch location of Ps. georgianus Balleny I. Russia 2004/05 S. Sandwich I. Germany 1975/76; 1980/81 Kerguelen I. Australia 2003/04 Known catch location of fish: Ps. georgianus: 0-475 m; 53°S-66°S ?
- 154. Shag Rock S.Georgia I. S.Sandwich I. S. Orkney I. Elephan I. K.GeorgeI. Deception Palmer A. Balleny Kerguelen I. Bouvet I. Heard I. Ch. aceratus: 5-770 m; 53°S-65°S Ch. gunnari : 0-700 m; 48°S-66°S Unusual catch location of Ps. georgianus Balleny I. Russia 2004/05 S. Sandwich I. Germany 1975/76; 1980/81 Kerguelen I. Australia 2003/04 Known catch location of fish: Ps. georgianus: 0-475 m; 53°S-66°S ?
- 155. Shag Rock S.Georgia I. S.Sandwich I. S. Orkney I. Elephan I. K.GeorgeI. Deception Palmer A. Balleny Kerguelen I. Bouvet I. Heard I. Ch. aceratus: 5-770 m; 53°S-65°S Ch. gunnari : 0-700 m; 48°S-66°S Unusual catch location of Ps. georgianus Balleny I. Russia 2004/05 S. Sandwich I. Germany 1975/76; 1980/81 Kerguelen I. Australia 2003/04 Known catch location of fish: Ps. georgianus: 0-475 m; 53°S-66°S ?
- 156. Shag Rock S.Georgia I. S.Sandwich I. S. Orkney I. Elephan I. K.GeorgeI. Deception Palmer A. Balleny Kerguelen I. Bouvet I. Heard I. C. aceratus: 5-770 m; 53°S-65°S C. gunnari : 0-700 m; 48°S-66°S Unusual catch location of Ps. georgianus Balleny I. Russia 2004/05 S. Sandwich I. Germany 1975/76; 1980/81 Kerguelen I. Australia 2003/04 Known catch location of fish: Ps. georgianus: 0-475 m; 53°S-66°S ?
- 157. Shag Rock S.Georgia I. S.Sandwich I. S. Orkney I. Elephan I. K.GeorgeI. Deception Palmer A. Balleny Kerguelen I. Bouvet I. Heard I. C. aceratus: 5-770 m; 53°S-65°S C. gunnari : 0-700 m; 48°S-66°S Unusual catch location of Ps. georgianus Balleny I. Russia 2004/05 S. Sandwich I. Germany 1975/76; 1980/81 Kerguelen I. Australia 2003/04 Known catch location of fish: Ps. georgianus: 0-475 m; 53°S-66°S ?
- 158. East Wind Drift Antarctic Circumpolar Current Polar Front East Wind Drift Antarctic Circumpolar Current Polar Front Channichthyidae concentrated in eddies and swim in current greatest in a World facilitating migration - they have running in large and small back and forth branches. In winter, the ice cover with reach under icy living world distributes the larvae of fish from shelf to the open ocean, connecting habitat between islands. Wide near and under-ice distribution of krill secures there the food to near-shore larvae lives in that caring under ice world on the open ocean. Channichthyidae have different flattening and proportions of otoliths that indicate different swimming capabilities specialized to cold sea currents in different environments. The fish larvae drift for food (Hecq, 2007)
- 159. In winter, the ice cover with reach under icy living world distributes the larvae of fish from shelf to the open ocean, connecting habitat between islands. In the 80s can reached South Georgia. Data: Sahrhage, 1988; Murphy, 2013; Bargelloni L., 2000; Kaufmann R.S., 1995; Eicken, 1992; Vincent, 1988
- 160. Wide near and under-ice distribution of krill secures there the food to near-shore larvae lives in that caring under ice world on the open ocean.Data: Sahrhage, 1988; Murphy, 2013; Bargelloni L., 2000; Kaufmann R.S., 1995; Eicken, 1992; Vincent, 1988
- 161. 161 At the surface, where the currents are strong, there were large fish. In the surface waters there were found only 3 postlarvae of Ps. georgianus. Rest, about 100 postlarvae were deeper, where water currents are weaker. 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 24 91 96 43° 42° 41° 40° 39° 38° 37° 36° 35° 34° 55° 54° Shag Rocks 10 - 12 XII 1986 (N = 115) 15 XII 1987 - 4 I 1988 (N = 134) 1 - 13 II 1989 (N = 65) 2 - 29 I 1990 (N = 70) 5 - 30 I 1991 (N = 87) lack of Ps. georgianus in the catch (N = 79);. bottom captureof the juvenes and adults (N = 26). pelagical capture of thejuveniles (N = 6); bottom captureof the juveniles (N = 6); lines - bottom capture of theadults Captureof Ps. georgianus 55° 54° 500m 500m 12 13 14 15 16 17 18 19 20 21 22 23 25 26 27 7 8 9 10 11 57 6160 565554 59 58 97 65 64 63 103 62 99 104 105 30' 30' 30'30' 96-105: old squares 1-27: new squares 654 321 9=55+92 10=56+93 19+23=64 7+12~96 16=61 15=60 14=59 11=57 150m 30' 30' Data: Traczyk, 2012, 2013
- 162. Juvenile Ps. georgianus also occurred in shallow pelagic waters, but in the case of running isobath of 150 m and in the northeastern part of the island, more sculptured and sheltered from the wind and West current. In places where there were greater depths juvenile observed deeper. In the surface water, in summer warmer by about 2 ° C, instead that the fish may have a higher rate of digestion and growth, they were not colonized by larvaes and juveniles, but only by a few large fish, better than larvaes swimers . Often in the layer of water limited by 150 m isobath juvenile fish did not occur with adults. Hence it can be assumed that, as in the Antarctic zone, here juvenile fish inhabit cooler, deeper water, with weak currents. Older fish inhabit warmer shallower waters, with strong current. Postlarvae of Ps. georgianus do not occur at the surface, they have high activity of antifreeze proteins. Post- larva of C. aceratus do so, because it has low activity of antifreeze proteins AFGP and from that could swim in warmer water. C. aceratus Ps. georgianusAll larvae No/1000m3 Data: Traczyk, 2013; Traczyk, 2012; North, 1991; Bilyk, 2011
- 163. The shape of the otoliths Channichthyidae: Ps. georgianus, C. aceratus and C. gunnari are similar due to the similar strategy of swimming – they have a similar body shape. However that species have a little different otolith shape and body what indicate
- 164. C. gunnari: 0,98°C -1,85°C Ch. wilsoni: 1,29°C -2,23°C Antarctic Circumpolar Ps. georgianus: 1,03°C -1,91°C Cold water descent Activity AFGP [°C] Blood feeezing: [°C] change tolerance of water temperature and swimming speed (pelagic life to bottom; temperate to high Antarctic) in a result of different content of AFGP. Why there are swimming differences, where are the causes and how it is go? Ch. hamatus: 1,45°C -2,44°C WHITE-BLOODED: high Antarctic; ice pack zone; temperate 80°S 74, ~70°S; 63, ~60°S; 63, ~57°30’S; 52, ~45°S 30°S S. Orkney SouthGeorgia -100 -200 -300 -400 -500 -600 -700 -800 Current C. aceratus: 0,54°C -1,47°C [m] Bilyk 2011
- 165. C. gunnari: 0,98°CCh. wilsoni: 1,29°C -2,23°C Antarctic Circumpolar Ps. georgianus: 1,03°C -1,91°C Cold water descent Activity AFGP [°C] Blood feeezing: [°C] Ch. hamatus: 1,45°C -2,44°C WHITE-BLOODED: high Antarctic; ice pack zone; temperate 80°S 74, ~70°S; 63, ~60°S; 63, ~57°30’S; 52, ~45°S 30°S S. Orkney SouthGeorgia -100 -200 -300 -400 -500 -600 -700 -800 Current C. aceratus: 0,54°C -1,47°C [m] increase in activity and production of antifreeze proteins -1,85°C increaseinactivityandproductionofAFGP Why are there swimming differences, where are the causes and how it is go? Bilyk 2011 Together or parallel with their evolution. Bilyk, 2011; Wöhrmann, 1996; Clarke A., 1996; Cheng, 1999; Chen, 250)
- 166. Antarctic Circumpolar WHITE-BLOODED: high Antarctic; ice pack zone; temperate 80°S 74, ~70°S; 63, ~60°S; 63, ~57°30’S; 52, ~45°S 30°S S. Orkney SouthGeorgia -100 -200 -300 -400 -500 -600 -700 -800 Current [m] Ch. hamatus: Nototheniidae; 0,4; 2,5 P. antarcticum [mln./mm3]; [g/100 ml] Nototheniidae; 0,8, 8,3 S. japonicus: 4, 18 Channichthyidae: 0; 0 Ch. esox: 0; 0 L. squamifrons Ch. aceratus Ps. georgianus 0; 0 Bathydraconidae: 0,2; 0,8 Macrouridae; 0,99, 3,9 C. gunnari 0; 0 0; 00; 0 All species of Channichthyidae have lost hemoglobin that reduced oxygen transport Jakubowski, 1971; Near, 2010; Everson, 1977; Fisher, 1985
- 167. Antarctic Circumpolar WHITE-BLOODED: high Antarctic; ice pack zone; temperate 80°S 74, ~70°S; 63, ~60°S; 63, ~57°30’S; 52, ~45°S 30°S S. Orkney SouthGeorgia -100 -200 -300 -400 -500 -600 -700 -800 Current [m] reduction in thenumber of and content Ch. hamatus: Nototheniidae; 0,4; 2,5 P. antarcticum [mln./mm3]; [g/100 ml] Nototheniidae; 0,8, 8,3 S. japonicus Channichthyidae: 0; 0 Ch. esox: 0; 0 L. squamifrons Ch. aceratus Ps. georgianus 0; 0 Bathydraconidae: 0,2; 0,8 Macrouridae; 0,99, 3,9 C. gunnari 0; 0 0; 00; 0 reductionofredcellsandhem 4; 1.8 B U T I N S T E A D O F T H E M A I N T R E N D ( W I T H L O W E R I N G W AT E R T E M P E R AT U R E ) LOWTEMPERATURE hem Jakubowski, 1971; Everson, 1977
- 168. Antarctic Circumpolar WHITE-BLOODED: high Antarctic; ice pack zone; temperate 80°S 74, ~70°S; 63, ~60°S; 63, ~57°30’S; 52, ~45°S 30°S S. Orkney SouthGeorgia -100 -200 -300 -400 -500 -600 -700 -800 Current [m] Ch. hamatus: Nototheniidae; 0,4; 2,5 P. antarcticum [mln./mm3]; [g/100 ml] Nototheniidae; 0,8, 8,3 S. japonicus: 4, 18 Channichthyidae: 0; 0 Ch. esox: 0; 0 L. squamifrons Ch. aceratus Ps. georgianus 0; 0 Bathydraconidae: 0,2; 0,8 Macrouridae; 0,99, 3,9 C. gunnari 0; 0 0; 00; 0 they spread on all Antarctica from temperate to high Antarctic waters, from surface across pelagic to bottom deep waters. Jakubowski, 1971; Fisher, 1985
- 169. High Salinity Shelf Water Antarctic Surface 34,18 ‰-100 -200 -300 -400 -500 -600 Removingfishfrom theshelfbyglaciers IncreaseO2pressurein thebloodandtissues increasesaturationO2 inhemoglobin dropintemperature whenpO2=const. increasing the affinity of hemoglobin for O2 impedes, even impairs putting into tissues; Blood viscosity increase when the drop in temperature 100% O2 Dyfuzja 70-90% O2 Water d i f f u s i o n lowers blood viscosity, and this > flow, transport of O2 [m] ? Not ! → Bottom fish Fishmigration Fishmigration reduction of red blood cells changeswith changeswith changeswith Fishmigration migrationto bottom Jakubowski, 1971; Near, 2010; Everson, 1977; Fischer, 1985 Kunzmann, 1991; Rakusa - Suszczewski, 1989; White, 1977
- 170. High Salinity Shelf Water Antarctic Surface 34,18 ‰-100 -200 -300 -400 -500 -600 Removingfishfrom theshelfbyglaciers IncreaseO2pressurein thebloodandtissues increasesaturationO2 inhemoglobin dropintemperature whenpO2=const. increasing the affinity of hemoglobin for O2 impedes, even impairs putting into tissues; Blood viscosity increase when the drop in temperature 100% O2 Dyfuzja 70-90% O2 Water lowers blood viscosity, and this > flow, transport of O2 [m] ? Not ! → Bottom fish Fishmigration Fishmigration reduction of red blood cells changeswith changeswith changeswith Fishmigration Reduction in the number of red cells and heme reduces transport, storage of oxygen to the muscles of the body and thus their reduction and develop swimming strategy with low energy in all Antarctic species. migrationto bottom
- 171. At first white-blooded in relation to red-blooded have a larger size so they have reduced heat loss due to the lower surface. Bergman’s rule of energy benefits. Channichthyidae achieve larger body size. eggs, mm Larvae, mm Adult, cm incr, cm/y White-blooded – Channichthyidae 5 4-17 43 6-10 Red-blooded – Nototheniidae 36 Red-blooded – Bathydraconidae 26 Red-blooded – Harpagiferidae 15 To fulfill strategy of swimming speed to different environment each species has a different compensation of reduced oxygen transport what differentiates their ability to swim and thus differentiates the shape of the body and the otoliths. Icefish have different life strategies and occupy different habitats (La Mesa M., 2004) 4 1 2 2 4 8 Mass = 1 Mass=8 Area = 28 Area = 112 Area/mass = 28 Area/mass = 14 Jakubowski, 1977; Wells, 1985; Kock, 1991; Johnston, 1983 additional oxygen from skin breathing of large head for the heart (Detrich, 2012).
- 172. Bottom lifestyle of Channichthyidae. Chaenocephalus aceratus has the body weak with a reduction in the axial muscles, which is probably due to the large volume of blood, up to 9% of body weight usually poorly vascularised. Jakubowski, 1977; Wells, 1985
- 173. White-blooded also have a reduction in ossification as a result of displacement of benthic fish fauna from shelf by glaciers to greater depth or to the pelagic. It enforced a reduction in body weight, because the white-blooded does not have a swim bladder. Walesby, 1982; Jakubowski, 1977 Żabrowski, 2000; Byrd, 2012
- 174. Icefishhuk. Additional oxygen from skin breathing of large head for the heart (Detrich, 2012; Kils, 2008).
- 175. Walesby, 1982; Twelves, 1972 Pelagic life of Channichthyidae. Lack of myoglobin, which enhances oxygen diffusion by 600% should limited locomotion activity of muscles becase lack of oxygen. It is not for C. gunnari, which increases oxygenation by larger capillaries and large spaces supplying capillaries with blood.
- 177. The activity of alkaline phosphatase determines the size of the muscle vasculature. Reis, 1970. Greatest is in C. gunnari hence this species has most capillaries. ±s [enzyme units/g wet muscle×h] species / muscle type pectoral fin oxidative glycolytic C. gunnari, n=7, 8, 8 1185,5±101,4 770,0±62,5 400,0±43,4 Notothenia rossii, n=8, 9. Gadus morhua, n=10 698,1±118,5 405,8±37,5 483,2±71,0 353,5±65,1 310,0±37,6 100,3±19,4 Its slender body shape increases heat loss, by Allen’s rule. 16 6 1 Mass = 8 Area = 192+12+32=236 Area/mass = 236/8=29,5 Mass=8 Area = 96+48+16=160 Area/mass = 160/8=20 2 2 4 12 Ps. georgianus C. gunnari It is agree with Allen's Rule for energy benefits of having a more slender (less resistance) in the warm waters.
- 178. Low-energy swimming is on the pectoral fins – only? High energy swimming by body waves Channichthyidae Nototheniidae Antarctic Gadidae C. gunnari N. rossii Pollachius virens Oncorhynchusmykiss myomer muscule lateral line channel oxi 1 oxi 2 Oxi 2 mosaic oxi 1 & oxi 2 White fibers, fast reaction mosaic of white and red fibers red fibers oxidative slow reactionAltringham, Ellerby, 1999; Davison i Macdonald 1985, Harrison i in. 1987, Walesby 1982 swimming on the pectoral fins saves more energy (than by body waving), because the muscles of that fins are slow oxidative fibers adapted to continuous low-intensity movements (consuming less energy), ensuring the long-term swimming at low speed. C. gunnari has also oxidative and glycolytic fibers in axial muscles. Oxi 1 pectoral fin muscle