This document summarizes a doctoral thesis on the pharmaceutical and immunological challenges of fungal pathogens. The thesis explored the interactions between the pathogenic fungus Candida albicans and human immune cells like neutrophils and mast cells. It developed a high-throughput screening assay to identify small molecules that block the yeast-to-hypha transition in C. albicans, which is important for its virulence. The screening revealed several FDA-approved drugs with previously unknown antifungal activity. The thesis provides new insights into antifungal defenses and tools to discover more effective antifungal therapies.















![2
has evolved resistance to fluconazole by drug target mutation, whereas C.
krusei is naturally-resistant. In that line, hospitalized individuals are
frequently administered with fluconazole due to suspicion for a mycose or as
routine prophylactic procedure allowing an increase with severe mycoses
caused by C. glabrata or C. krusei 7; 9; 10.
1.2 C. albicans polymorphism
Virulence to cause invasive or superficial infections is closely related to the
capability of C. albicans to reversibly switch between budding yeast and
filamentous forms (Y-H transition)11. Filaments exist as pseudohyphae or
true hyphae (figure 1). Pseudohyphae are characterized by constrictions of
the septum as a chain of unseparated yeast cells with dissimilar cell walls11.
On the other hand, a true hypha grows apically from the mother cell which
periodically can form branches. This polarized growth has perfectly parallel
cell walls without constrictions to the septae11.
Figure 1: Candida albicans morphotypes12. C. albicans filamentation is defined as
switching from yeast (lower left) to hyphae (lower right) or to pseudohyphae (upper).
[Re-printed with permission of the Nature Publishing Group, Licence 3687551119070]](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-15-320.jpg)

![4
Figure 2: Regulatory network of Y-H transition in Candida albicans 15. [Granted Re-
printing permission from American Society for Microbiology]
Farnesol, a quorum-sensing molecule, is secreted by C. albicans to disturb
the Y-H transition via TUP1 activation22-24. It is also assumed to participate
in yeast relocation from biofilms in order to disseminate the infection. In
addition to farnesol, homoserine lactone (HSL) has antidimorphic activity by
blocking hyphal growth. HSL has been observed to be produced and secreted
from Pseudomonas aeruginosa in co-infections with C. albicans22-24. HSL
influences the transcriptional upregulation of TUP1, NRG1 and of yeast wall
protein (YWP1), leading to inhibition of Y-H transition and biofilm
impairment23.
Taken together, the Y-H transition in several fungi and particularly in
C. albicans is an essential virulence trait of the pathogen to establish
successful colonization and infection. Consistent with this notion,
C. albicans cells are incapable to invade human cells, to escape phagocytosis,
and to form biofilms when Y-H transition is disturbed resulting in an](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-17-320.jpg)

![6
Figure 3: Candida albicans cell wall architecture 36. [Re-printed with permission of the
Nature Publishing Group, Licence 3690810405720]
A different escaping mechanism of C. albicans cells is the production of
various superoxide dismutase proteins (Sod) to oppose neutrophil and
macrophage ROS37. C. albicans possesses cytosolic and mitochondrial Sods
to detoxify ROS. Upon superoxide (O2
-) production, the primary ROS during
the oxidative burst from the host cell, C. albicans expresses Sods which
convert O2
- to hydrogen peroxide (H2O2), while in turn catalase protein
(Cat1p) decomposes H2O2 to H2O and O2
38. To date, Sod1p (cytoplasmic),
Sod2p (mitochondrial), Sod3p (cytoplasmic), Sod4p, Sod5p and Sod6p (cell
surface) have shown to prevent host’s hazardous oxygen radicals37-39.](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-19-320.jpg)
![7
Figure 4: Candida albicans cellular components recognized by human immune
system36. [Re-printed with permission of the Nature Publishing Group, Licence
3690810405720]
The fact that Sods 4-6 are located on the cell surface indicates that they play
a role in protection against host-originated ROS stress. Indeed, Sod4p and
Sod5p have shown to counteract the oxidative burst as demonstrated by
reduced cellular viability of SOD4 and SOD5 double-KO mutants when
interacting with macrophages37. In good agreement, Sod5p is induced under
hyphal growth, which is essential for escape from macrophage
phagolysosomes38; 40.
Another example of a C. albicans defense strategy is the characteristic
evasion of the complement system. The human complement system is
mediated by three pathways, the classical, the alternative, and the lectin
pathway. The classical pathway initiates via an antibody-antigen complex,
while, the alternative pathway is antibody-independent and induced by
PAMPs on microbial surfaces41. The lectin pathway is activated via binding of
host-derived lectin to fungal mannose- and mannan-related PAMP
molecules. The lectin pathway, as the other two pathways, drives the
generation of C3 convertase to hydrolyse C3 to C3a and the C3b opsonin.](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-20-320.jpg)


![10
Figure 5: Neutrophil with the characteristic lobulated nucleus (red) and granules
(yellow)49. [Granted re-printing permission from Wikiversity Journal of Medicine]
In addition to above defense tools, neutrophils are also able to prevent
pathogenicity by “sacrificing” themselves, via a novel cell death process.
Upon contact to pathogens, neutrophils undergo a programmable molecular
signalling and intracellular rearrangement to eventually release neutrophil
extracellular traps (NETs)50. NETs are web-like structures composed of a
nuclear DNA scaffold decorated with cytoplasmic and granular material51. By
producing NETs Neutrophils entrap microbes and in the case of pathogenic
fungi, such as C. albicans, NETs have a fungistatic mechanism. Mainly the
NET–bound protein-calprotectin, a zinc-chelator, drives C. albicans to
growth arrest, when it is entangled in NETs52. Eventhough NETosis (NET-
forming process in analogy to apoptosis) is a very defined and distinct form
of cell death and continuously new triggering stimuli are revealed the exact
molecular details and regulatory networks remain poorly defined.
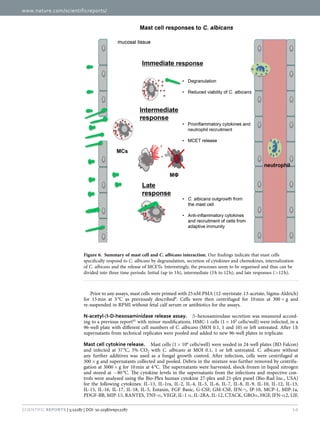
2.2 Mast cells
Mast cells (MCs) are tissue-resident leukocytes originated from
hematopoietic progenitors (figure 6). The respective progenitors are primed
with stem cell factor (SCF) which binds on SCF receptors/CD117/tyrosine-
protein kinase also termed c-kit53-55. After differentiation and maturation
MCs leave the peripheral blood to be further differentiated into two dinstict
subtypes, the mucosal MC (MCT) and the connective tissue MC (MCTC)56; 57
(table 1). The basic differences of MCT and MCTC are their residency to](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-23-320.jpg)
![11
different tissues and granular composition as well as the chymase expression
which is absent in MCT
56; 57.
Figure 6: Hematopoietic tree and mast cell maturation41. [© 2007 Janeway's
Immunobiology, Seventh Edition by Murphy et al. Reproduced by permission of Garland
Science/Taylor & Francis Group LLC.]
2.2.1 Mast cells in allergies
MCs have strong IgE-binding capacities via the expression of the high-
affinity IgE receptor, FcεRI, whereas other IgE-receptor-associated cells,
such as eosinophils, express IgE-binding receptor FcεRII with lower affinity
than FcεRII 57; 58. IgE as well as other secreted allergic triggers, such as
cytokines, anaphylatoxins or neuropeptides59, have beyond doubt connected](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-24-320.jpg)
![12
MCs with allergic responses. Indeed, human MCs undergo IgE binding upon
activation with various stimuli, such as antigen/allergen (peanut, pollen and
latex), allowing mediator production57; 60. The mediator secretion is classified
into two distinct mechanisms, the preformed mechanism and the newly-
synthesized molecule mechanism56;57;61. Preformed mediators are “ready-to-
go” molecules such as histamine, heparin, serotonin, cathepsin G and TNF-
α. Examples of newly-synthetized mediators are nitrogen and oxygen
radicals, different cytokine and chemokines (TNF-α, MCP-1 and MIP-1α) as
well as various lipid mediators (Prostagladin D2 and leukotrienes)56; 57; 61.
Table 1: Differences of mast cell subtypes 57. [Re-printed with permission from Springer
Publishers, Licence 3687600957650].
Feature MCTC cell MCT cell
Structural features
Grating/lattice granule ++ –
Scroll granules Poor Rich
Tissue distribution
Skin ++ –
Intestinal submucosa ++ +
Intestinal mucosa + ++
Alveolar wall – ++
Bronchi + ++
Nasal mucosa ++ ++
Conjunctiva ++ +
Mediator synthesized
Histamine +++ +++
Chymase ++ –
Tryptase ++ ++
Carboxypeptidase ++ –
Cathepsin G ++ –
LTC4 ++ ++
PGD2 ++ ++
TNF-α ++ ++
IL-4, IL-5, IL-6, IL-13 ++ ++](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-25-320.jpg)
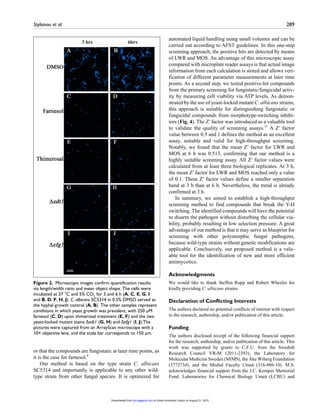
![13
2.2.2 Mast cells and infections
MCs have versatile roles beyond allergies, as they actively participate in
different autoimmune diseases such as rheumatoid arthritis, asthma and
systemic sclerosis56;57. Furthermore, MCs are associated to inflammatory
diseases, such as, graft-versus-host disease, fibrotic disease and ischemic
heart disease as well as in various infectious diseases 56; 57; 62. Human MCs
immediate response against GAS (Group A Streptococci) comprises ROS
production and the release of in vitro and in vivo antimicrobial molecules,
similar to Neutrophils 63.
Figure 7: Role of mast cell responses to different microbial pathogens62.[Re-print
permission from Plos publisher]
PRRs expressed by MCs include TLRs, such as TLR2 and TLR4, well-defined
receptors for recognition of bacteria and fungi. MCs also express non-TLRs,
C-type lectin receptors, such as Dectin-1 and Mincle, for the identification of
fungal components (figure 7)64-66. The antibacterial contribution of MCs
includes the secretion of proteases and tumour necrosis factor alpha (TNF-
α). In vivo experiments highlighted a key role of MCs in innate immunity
against bacterial pathogens, such as for instance Escerichia coli, Klebsiella](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-26-320.jpg)
![14
pneumonia and Listeria monocytogenes, as indicated by a considerable
decrease of mouse survival in infected mast cell-deficient mice compared to
wild-type littermates67. Similar to NETs, mast cell extracellular traps
(MCETs), have been described, at first as a response to S. pyogenes68.
MCETs ensared and killed the bacteria (figure 8)69. Again, in analogy to
NETs, MCETs are composed by a DNA scaffold decorated with granular
proteins and antibacterial components, such as tryptase and LL-3768.
Figure 8: Mast cell extracellular traps. Staphylococcus aureus cells (black arrows)
ensnared by MCETs (white arrows) (modified from Jens Abel et al 2011)69. [Re-printed with
permission from Journal of Innate Immunity, Licence 3693791275161]
MCs mediated responses to viruses are poorly investigated, MCs recognize
viruses or viral double-stranded RNA via TLR3 which mediates the secretion
of antiviral type I interferons (IFNs) 70. Interestingly, in vitro and in vivo
MCs are reported to trigger antiviral properties of CD8+ T cells.
In summary, data exists which highlights the importance of MCs in defence
and clearance of bacteria and viruses, whereas MC-fungus interactions
virtually remain a “clean sheet”71; 72.](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-27-320.jpg)
![15
3.0 Antimycotics and their mode of action
An increase in the number of immunosuppressed patients due to cancer,
organ or bone-marrow transplantation and cystic fibrosis set the ground for
the emergence of opportunistic fungal pathogens. The lack of rapid
diagnostics and efficient antimycotics, results in emerging fungal
infections73. The development of a new class of antimycotics requires a better
understanding of Candida colonization, infectivity and virulence. Current
fungal antibiotics are few with a narrow target range31;32. The most-
frequently used classes of antifungals are polyenes, azole, allylamines and
echinonocandins (figure 9).
Figure 9: Fungal cell anatomy and antifungal targets74; 75. Polyenes and azoles affect
directly or indirectly the ergosterol integrity as well as the β-glucan synthesis of the cell wall.
Other drugs target DNA and protein synthesis.[Adapted from Shankar et al 2013]
3.1 Polyenes
Polyenes aim for ergosterol, the functional analog of cholesterol in
mammalian cell membranes and biosynthesis of this molecule is essential for
fungal growth. Therefore, it is an ideal target for antifungal agents 74. The
mode of action is characterized by channel formation in the plasma](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-28-320.jpg)













![29
Figure 14: Orchestrated responses of human mast cell upon encounter with
Candida albicans. [Re-produced with permission from Nature publishing group109]
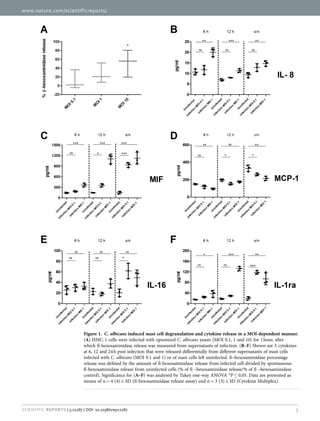
During the intermediate phase MIF concentration increased, whereas IL-8,
though decreased, remained significantly above UC. Importantly, hMCs
release MCETs which begins from the intermediate phase and lasts until the
late phase. Even though hMCs could not terminate fungal growth, the fungal
cells were entrapped within the MCETs according to microscopic
investigation. Finally, the late phase demonstrates the potency of hyphal
forms to cause MC lysis originating either from extracellular and
intracellular space. The cell death in hMCs is C. albicans-mediated either
due to hyphal penetration or due to induction of MCETs (figure 15 A + C). In
addition to MIF, MCP-1 and IL-8 secretion, at late phase, the cytokines IL-16
(adaptive immunity-related) and IL-1rα (anti-inflammatory protein) were
released.](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-42-320.jpg)
![30
Figure 15: Mast cell anticandida and C. albicans antimast cell activity. MCET
secretion upon C. albicans stimulation (A). Transient antifungal activity of MCs which is not
dependent on MCETs as determined with DNase digest of DNA traps (B). . MC cell death
induced by C. albicans is time and concentration dependent (C).[Re-produced with permission
from Nature publishing group109]
This study illustrates the responses of hMCs upon interaction with C.
albicans. MCs secrete cytokines related to innate and adaptive immune cells.
In addition, hMCs show a transient antifungal activity which is independent
from MCET production. From the pathogen point of view, C. albicans
displays antimast cell activity leading to mast cell lysis by hyphal invasion
either from outside-to-inside or from inside-to-ouside. Conclusively, hMCs
indeed serve as tissue-sentinels for commensal fungal pathogen. It will be
beneficial for the development of new antifungal agents to gain more insight
into MC responses to other fungal pathogens.](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-43-320.jpg)

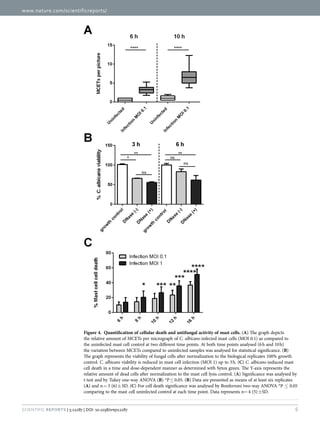
![32
(MOS = [(c2/4π)*area]). Circular objects therefore have a MOS of 1 and
filamentous of >1.5. Wild-type C. albicans grown under hypha-inducing
conditions at 3 hours and 6 hours resulted in LWR and MOS values above
1,5, while grown under yeast-inducing conditions resulted in LWR and MOS
values below 1.5 (figure 16). The control conditions including dead, mutant
and farnesol-treated C. albicans additionally showed LWR and MOS values
below 1.5. Hence, we used a threshold of LWR and MOS of 1.5 to clearly
define yeast or hyphal cells, respectively.
Figure 16: Distinction of yeast and hyphal cells. Different LWR and MOS values for the
indicated strains and conditions are shown. LWR after 3 hours (A) and 6 hours (B) as well as
MOS after 3 hours (C) and 6 hours (D). [Re-produced with permission from Sage publisher113
To distinguish fungistatic or fungicidal compounds from agents that purely
inhibited morphotype transition without affecting yeast growth we added an
additional layer to the assay. We tested for C. albicans viability using ATP
quantification. Only metabolically active and thus living cells produce and](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-45-320.jpg)
![33
retain measurable ATP levels (figure 17). This assay is reliable, rapid and is
not affected by the hyphal morphotype. Serial dilution and plating for colony
counting cannot be applied for quantification of hyphae, since growing cells
do not separate and hyphal filaments additionally tend to clump. Both
features lead to false results in quantification methods based on colony
counting. The ∆edt1 and ∆efg1 strains as well as farnesol-treated C. albicans
resulted in ATP levels close to untreated control. Dead C. albicans and
farnesol-treated overnight resulted in negligible ATP levels (figure 17).
Until today, other assays have been suggested for detection of antidimorphic
molecules96-98. However, the described assays are dependent on reporter
strains which are based on the promoter of hyphal wall protein 1 (HWP1).
HWP1 is expressed under hypha-inducing conditions and repressed during
yeast growth. As reporters placed downstream of the promoter the open
reading frames (ORFs) for either green fluorescent protein (GFP)97; 98 or
beta-galactosidase (lacZ)96 were used.
Figure 17: The cellular viability of Candida albicans when challenged with
farnesol and thimerosal and the cellular viability of Candida albicans mutant
yeast-locked strains. Candida albicans cellular viability was recorded after 3 (A), 6 (B) and
24 (C) hours. Farnesol blocks the Y-H switching, but retains the cellular viability: in contrast
thimerosal kills the cells. For screening purposes farnesol represents an ideal antidimorphic
compound, thimerosal mimics a fungistatic / fungicidal agent. [Re-produced with permission
from Sage publisher 113]
An advantage of our method is the fact that it is applicable for type fungal
strains and not dependent on genetically-modified strains, such as GFP and
lacZ reporter strains. During development we accounted for the antifungal](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-46-320.jpg)



![Table 2 : Candida spp. susceptibility to seven novel antifungal off-patent drugs
[Re-produced with permission from ASM publisher117]
MIC MIC0.3 MIC MIC0.3 MIC MIC0.3 MIC MIC0.3 MIC MIC0.3 MIC MIC0.
3
Haloperidol HCl 6.4 × 10−3
to 3.76 3.76 0.46 3.76 0.38 3.76 0.38 > 3.76 0.38 >3.76 3.76 >3.76 >3.76
Trifluperidol 2HCl 7 × 10−3
to 4.00 4.00 0.40 >4.00 0.40 >4.00 >4.00 > 4.00 0.40 >4.00 >4.00 >4.00 >4.00
Stanozolol 3.3 × 10−3
to 3.29 >3.29 0.33 >3.29 0.33 >3.29 3.29 >3.29 3.29 >3.29 >3.29 >3.29 3.29
Melengestrol acetate 6.8 × 10−3
to 3.97 3.97 0.37 3.97 0.40 >3.97 3.97 >3.97 1.80 >3.97 3.97 >3.97 3.97
Megestrol acetate 6 × 10−3
to 3.85 3.85 0.39 3.85 0.39 >3.85 3.85 >3.85 3.85 >3.85 3.85 >3.85 3.85
Tosedostat 4 × 10−3
to 4.00 >4.00 4.00 >4.00 4.00 >4.00 4.00 >4.00 4.00 >4.00 4.00 >4.00 2.00
Amonafide 2.8 × 10−3
to 2.83 >2.83 1.40 >2.83 2.83 >2.83 2.83 >2.83 1.40 >2.83 >2.83 >2.83 >2.83
Methiothepin maleate 7 × 10−3
to 3.57 3.30 0.31 3.30 0.36 >3.57 0.36 >3.57 0.36 3.57 0.36 3.57 0.36
Auranofin 4 × 10−3
to 6.78 0.68 0.08 0.61 0.07 0.68 0.04 0.62 0.04 1.10 0.62 >3.73 3.73
Rapamycin 9 × 10−3
to 9.14 0.002 <9 × 10−3 0.002 <9 × 10−3 0.009 <9 × 10−3 0.01 <9 × 10−3 0.50 0.04 0.09 0.009
UBC3-7268
(clinical strain)
Antifungal agent Concn range
(μg/ml)
C. albicans C. dubliniensis C. glabrata
SC5314
(type strain)
UBC3-7922
(clinical strain)
CD36/CBS7987
(type strain)
UBC3-3892
(clinical strain)
ATCC 90030
(type strain)
37](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-50-320.jpg)
![38
Table 3: Standard antifungal drugs versus off-patent drugs. [Re-produced with
permission from ASM publisher117]
Drugs MIC MIC0.3
Standard antifungal
Tioconazole 0.39 μg/ml
Oxiconazole nitrate 0.40 μg/ml
Ketoconazole 0.50 μg/ml
Climbazole 0.29 μg/ml
Miconazole 0.40 μg/ml
Fluconazole 0.30 μg/ml
Amorolfine 0.32 μg/ml
Myclobutanil 0.29 μg/ml
Bifonazole 0.30 μg/ml
Sertaconazole 0.40 μg/ml
Itraconazole 0.70 μg/ml
Terbinafine HCl >1 μM
Nystatin >1 μM
Off-target antifungal
Haloperidol HCl 0.38 μg/ml
Methiothepin maleate 0.36 μg/ml
Auranofin 0.68 μg/ml
Trifluperidol 2HCl 0.40 μg/ml
Stanozolol 0.30 μg/ml
Melengestrol acetate 0.40 μg/ml
Megestrol acetate 0.39 μg/ml
Tosedostat >1 μM
Amonafide >1 μM](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-51-320.jpg)

















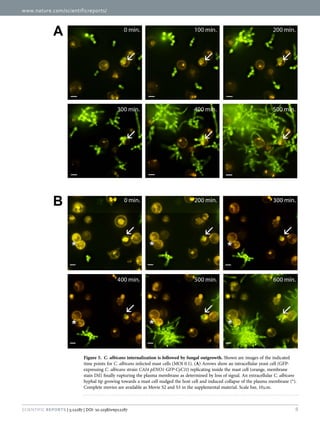

![www.nature.com/scientificreports/
11Scientific Reports | 5:12287 | DOI: 10.1038/srep12287
MCP-3, M-CSF, MIF, MIG, β -NGF, SCF, SCGF-β , SDF-1 α , TNF-β , TRAIL. Samples were mixed with
antibody-coated beads that have a unique fluorescent intensity for the above cytokines. Anti-cytokine
antibody PE-conjugated with streptavidin was added and the fluorescent signals were detected using a
multiplex array reader Bio-Plex 200 System (Bio-Rad Laboratories). Raw data were initially measured
as relative fluorescence intensities and then converted to cytokine concentrations based on the standard
curve generated from reference concentrations supplied by the manufacturer.
Concentrations of IL-8 in supernatants of C. albicans-infected CBMC were measured using a human
IL-8 enzyme-linked immunosorbent assay (ELISA) MAX kit (Biolegend, eBioscience USA). Primary
cells were infected with C. albicans at MOI1 or left uninfected for 6 h. Supernatants were harvested and
stored as described above.
Chemotaxis assay for human neutrophils and monocytes. Neutrophils were harvested from
blood of healthy volunteers according to the recommendations of the local ethical committee (Regionala
etikprövningsnämnden i Umeå). Fully informed consent was obtained, and all investigations were con-
ducted according to the principles stated in the Declaration of Helsinki. Neutrophils were isolated from
venous blood as previously described32
. Chemotactic migration of neutrophils and monocytes towards
supernatants of mast cells infected with C. albicans (MOI 0.1), uninfected controls or the equivalent
amount of fungal cells was measured using a transwell system as previously described32
. Neutrophils
migration was accessed for 30 min and for monocytes migration was accessed for 90 min. Mast cell
infected supernatants tested were collected and stored as described for the cytokine release assay at 6 h,
12 h and 16 h.
Immunostaining and microscopic analysis of mast cells. Mast cells (1 × 105
cells/well) were
seeded onto cover slips coated with 1% poly-l-lysine (Sigma-Aldrich) in 24-well plates and infected with
C. albicans (MOI 0.1, 1). Uninfected mast cells were used as control. After 6 h cells were fixed using 2%
paraformaldehyde and stored at 4 °C.
For visualization of MCETs, primary antibodies directed against human mast cell tryptase (clonal AA1,
mouse, DAKO) and C. albicans antibody (mouse monoclonal, ProSci) diluted in blocking solution were
applied overnight at 4 °C. Primary antibodies were detected with Alexa Fluor 488- and 568-conjugated sec-
ondary antibodies (Life Technologies). DNA was visualized with DAPI (4′ , 6′ -diamidino-2-phenylindole;
Life Technologies). Specimens were mounted in Pro-Long Diamond (Life Technologies).
Imaging data were acquired using a fully motorized inverted microscope (Nikon A1R Laser Scanning
Confocal Microscope) with 60 × oil immersion lens (Plan Apochromat VC; Nikon, Tokyo, Japan) under
control of the NIS-Elements microscope imaging software (Nikon). Final image composition was done
using Adobe Photoshop CS5 (San Jose, CA).
For live cell microscopy mast cells (2 × 105
cells/well) were stained with Vybrant DiI Cell-labelling
solution (Invitrogen) according to the manufacturer’s instructions and seeded into a 35-mm glass-bottom
micro- well dish (MatTek, Ashland, MA, USA). Mast cells were infected with C. albicans strain (CAI4
pENO1-GFP-CyC1t) at MOI 1 and kept at cell-culture conditions throughout the measurement. Frames
were captured 30 min post-infection at 60 × magnification every 10 min for a period of 16 h using the
previously described microscope.
Microscopic quantification was performed using DAPI immuno-stained image samples from six bio-
logical replicates. Images analysed had 130 ± 30 cells per picture and for each infection condition a total
of at least 1000 cells were analysed. The total number of cells was determined by ImageJ version 2.0. The
number of cells that underwent MCET formation was scored from binary images in a blinded fashion by
two trained researchers. Final scores were defined as MCETs per field of view and plotted by condition
and infection end-point.
From 10 independent live cell movies the % of cells undergoing inside-out growth was determined as
the ratio of [(inside-out growth)/100% total cells)]. Similar the % of cells undergoing outside-in growth
was determine as the ratio of [(outside-in side growth)/100% total cells)]. In both cases a total of 80 cells
was analysed.
Fungal viability measurement. To determine the antifungal effect of mast cells we compared fun-
gal viability in the presence of mast cells as follows: Mast cells (5 × 104
cells/well) were infected with
C. albicans at MOI 1 for 3 and 6 h in a 96-well plate coated with poly-l-lysine. The same amount of C.
albicans served as 100% control. To test for contribution of MCETs to fungal viability we added DNaseI
(Sigma Aldrich) prior to infection to one set of experiments. At the end point of the experiment, DNaseI
and subsequently Triton-X100 to a final concentration of 10% were added to all wells. The medium was
removed and fungal viability (ATP) was determined using CellTiter-Glo cell viability kit (Promega) in a
luminometer (Tecan Infinite F200) as previously described62
.
To normalize all values for comparable ATP signals, values were multiplied by the factor: [average of
technical replicates of 100% growth control]/[average of biological replicates of 100% growth controls].
Using these normalized values, the antifungal effect was determined as the ratio of [(infected MCs–
uninfected MCs)/100% growth C. albicans)].
To assure that any differences in cell viability were not due to loss of cells during washing we measured
absorbance before triton lysis and after adding CellTiter-Glo reagent and found no notable variation.](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-76-320.jpg)





![Stylianou et al. 287
with an automated microscope (HCA-Cellomics ArrayScan
VTI, Thermo Scientific) and the C. albicans cell morphol-
ogy analyzed. Based on HCA information, the individual
fungal cell morphotype was determined by means of LWR
and MOS, respectively. These two parameters were suffi-
cient to reliably discriminate between yeast and hyphal cul-
tures (eq 1). LWR determines the average ratio between
length and width, which indeed changes considerably dur-
ing apical growth of a filament versus division of ellipsoid
yeast cells by budding. MOS refers to the average measure
of detected objects based on the formula MOS =
[(c2
/4π)*area], which is the ratio of circumference squared
to 4π*area (MOS = 1 = circular object).
Determination of Cell Viability Using ATP
Quantification
We performed the cellular viability test prior to cell fixation
and chitin staining. The percentage of cellular viability was
determined using the CellTiter-Glo luminescent cell viabil-
ity assay (CTG; Promega, Madison, WI) to identify com-
pounds that are fungistatic or fungicidal. A volume of the
CTG reagent equal to the cell volume per well was added.
After 15 min at room temperature, the luminescence signal
was quantified in a luminometer (Infinite F200, Tecan,
Männedorf, Switzerland). The luminescence signal corre-
sponds to ATP values and thus to cellular viability. The per-
centage of cellular viability was calculated for the four
tested conditions, C. albicans either with farnesol (250 µM)
or thimerosal (0.8%) and the two mutant C. albicans strains
in comparison with C. albicans in DMSO (0.5%) as the
100% hyphal growth control (eq 2). The assay was per-
formed at least to three biological replicates in triplicate
(n = 3[3]) in 96-well plates with clear bottoms. Liquid han-
dling, plate reading, and automated microscopy were per-
formed at Laboratories for Chemical Biology Umeå
(LCBU), Chemical Biology Consortium Sweden (CBCS).
Calculations and Statistical Analysis
Growth inhibition (GrIn) was determined, after 3 and 6 h,
from the MOS and LWR values for all conditions. MOS and
LWR calculations are derived from the average number of
fluorescent pixels from at least 100 cells. The percentage
of GrIn (%GrIn
) was defined as eq 1: [%GrIn
=100 – ( x test
/
x DMSO
)*100]. Furthermore, the switching inhibition (SwIn)
(%SwIn
) was calculated from ATP values as eq 2: [%SwIn
=
100 – ( x test
/ x DMSO
)*100]. Thus, in high-throughput
screenings, the positive hits including growth and morphot-
ype inhibitors are determined using GrIn calculations. The
discrimination of growth from morphotype inhibitors is
defined by the SwIn formula. GrIn and SwIn calculations
were performed in Graphpad Prism 5.0 and analyzed for
statistical significance using a one-way analysis of variance
and Tukey’s multiple comparison test from at least three
biological replicates in triplicate (n = 3[3]) and applied for
0.5% DMSO (SC5314, Δedt1 and Δefg1), farnesol (250
µM), and thimerosal (0.8%). Moreover, the method validity
is defined by the Ζ′ factor as eq 3: [Ζ′ = 1 – [3*(SDDMSO
+
SDtest
)/(ABS( xDMSO
– x test
))]. The Ζ′ factors represent the
mean values from the calculation of at least three biological
replicates in triplicate (n = 3[3]).
Results and Discussion
A crucial virulence trait of polymorphic fungi is their ability
to reversibly switch from yeast-like to filamentous growth.
Hence, the aim of the study was to develop a reliable high-
throughput screening method for the identification of mol-
ecules that break the Y-H transition without disturbing cell
viability. Images gathered from an automated fluorescence
microscope were analyzed on the basis of fluorescent pix-
els. From the substantial amount of parameters created by
HCA, we chose LWR and MOS, because these values were
sufficient to reliably distinguish between yeast and hyphal
morphotypes (Fig. 1). This means in particular that C. albi-
cans samples with LWR and MOS values less than 1.5 are
defined as yeast cells (Fig. 1). After a 24 h incubation, LWR
and MOS values from the hyphal reference samples cannot
be taken into account, as confluent growth renders analysis
unfeasible. Microscopic images are nevertheless available
in substantial amounts for cell morphotype evaluation (data
not shown).
To validate whether our method is suitable for identify-
ing switching inhibitors, we used the quorum-sensing mol-
ecule farnesol. This natural compound prevents hyphal
growth of C. albicans under otherwise hyphae-inducing
conditions (Figs. 1–3). After 24 h of incubation with farne-
sol, however, C. albicans yeast growth was additionally
reduced to low levels, indicating that over long incubation
times, farnesol has growth-inhibitory activity. This is in
good agreement with a previous report that showed that
farnesol challenge of yeast cells prevented hyphal growth
but at the same time significantly reduced cellular viability.8
Farnesol is nevertheless a suitable reference for morphot-
ype switching inhibitors, because it does not affect C. albi-
cans growth within 6 h incubation periods (Fig. 3). We next
used thimerosal to kill off C. albicans cells, which after-
ward remain as dead and thus nonswitching yeasts. LWR
and MOS obtained from HCAdata confirmed that thimerosal-
treated C. albicans remained as yeasts, because values were
less than 1.5 and cellular viability was close to background
levels (Figs. 1–3). Thus, thimerosal could be used as a ref-
erence for fungicidal or fungistatic compounds.
We furthermore assayed two knockout C. albicans
mutants Δefg1 and Δedt1,9,10
both yeast-locked strains. They
serve as additional key references for determining the accu-
racy of discrimination between yeast growth versus hyphal
at Umea University Library on August 21, 2015jbx.sagepub.comDownloaded from](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-82-320.jpg)




![the susceptibility of C. albicans to SADs was comparable to the
antifungal effect of the seven agents identified in this screen.
MATERIALS AND METHODS
Drugs and fungal strains. The in vitro susceptibility of C. albicans strain
SC5314 was tested against 844 drugs from the Enzo FDA-approved drug
library (640 drugs) and the FIMM oncology collection (19) (FDA-ap-
proved anticancer drugs [n ϭ 119] and preclinical compounds [n ϭ 85]).
Thirteen FDA-approved antifungal drugs, 12 of which were active against
C. albicans SC5314, and five nonantifungal drugs with antifungal activity
served as controls. The screen was performed with C. albicans SC5314,
and hits were further confirmed with the type strains C. dubliniensis
CD36/CBS7987 and C. glabrata ATCC 90030, as well as with unrelated
clinical strains of C. albicans UBC3-7922, C. glabrata UCB3-7268, and C.
dubliniensis UCB-3892 from the strain collection of Norrland’s University
Hospital, Umeå, Sweden.
Media and antifungal microdilution testing. Cell concentration and
drug microdilution analyses were performed according to the European
Committee on Antimicrobial Susceptibility Testing (EUCAST) guide-
lines, with modifications (20). Candida yeast cells were grown overnight
at 30°C with shaking in yeast peptone medium plus 2% glucose (YPD).
Subcultures of 107
cells/ml in YPD grew for 4 h at 30°C. Drugs in the
amounts of 15 to 150 nl from the Enzo and FIMM oncology collections
were distributed by a liquid handling platform (Labcyte Echo 550 acoustic
dispenser) in black 96-well plates with clear bottoms in six different con-
centrations from 0.17 nM to 10 M. Subsequently, 50 l RPMI 1640 was
added to each well and the start plates were shaken (30 rpm) prior to the
assay for 1 h to ensure equal distribution of the agents within the well. The
yeast suspension, 100 l of 5 ϫ 105
cells/ml in RPMI 1640 without phenol
red, and 10 mM HEPES (Lonza) were transferred to the 96-well plates
containing medium and agents using a robotic device (Matrix WellMate;
Thermo Scientific), resulting in a final volume of 150 l in each well. The
plates were incubated at 37°C, 5% CO2, for 6 or 24 h.
Determination of fungal growth using absorbance. The growth of C.
albicans SC5314 was analyzed using a microdilution plate assay according
to EUCAST recommendations (20). One-hundred-microliter suspen-
sions of yeasts (5 ϫ 105
cells/ml) in RPMI 1640 were incubated in the
presence or absence of drugs in a total volume of 150 l at 37°C, 5% CO2,
for 6 h and 24 h. The optical densities at 450 nm (OD450) in the plates were
determined using a plate reader (Tecan Infinite F200). ODs of Ͻ0.1 for 6
h and 0.2 for 24 h for the 100% growth control were considered to repre-
sent poor growth and were not taken into account for the evaluation. As
described above, 100% and 0% growth controls were included with every
plate. All assays were performed at least as two biological replicates in
triplicate (n ϭ 2 [3]).
Determination of fungal viability using ATP levels. In order to de-
termine the viability of the C. albicans, C. glabrata, and C. dubliniensis
strains, the CellTiter-Glo luminescent cell viability kit (Promega) was
used. One hundred-microliter suspensions of yeasts (5 ϫ 105
cells/ml) in
RPMI 1640 were incubated in the presence or absence of drugs in a total
volume of 150 l at 37°C, 5% CO2, for 6 h and 24 h. An equal volume of
the CellTiter-Glo reagent was added to the medium and incubated for 15
min at room temperature with shaking at 900 rpm. The luminescent sig-
nals after 6 h and 24 h were detected using a luminometer (Tecan Infinite
F200). The resulting signal intensity corresponds to ATP amounts and
thus to the number of viable microbial cells upon drug exposure. In all
96-well plates, 100% and 0% growth controls were included as microbes
plus dimethyl sulfoxide (DMSO) (0.1%) and microbes plus benzetho-
nium chloride (BzCl) (100 M), respectively. All assays were performed at
least as two biological replicates in triplicate (n ϭ 2 [3]).
Microscopic analysis of morphological changes occurring upon
drug treatment. For a morphological analysis of C. albicans SC5314
treated with antifungal agents (1 M), an IncuCyte automated micro-
scope was used (Essen Bioscience). The plates were incubated at 37°C
under 5% CO2. After the indicated time points, prior to analysis, the fungi
were fixed with 2% paraformaldehyde (PFA) and phase-contrast images
were captured. In this study, 4 pictures per well were taken from two
biological and three technical replicates.
Statistical and data analysis. Percent growth inhibition (%Inh) was
calculated from the ATP and OD measurements resulting from the mean
values from all biological replicates, using the equation %Inh ϭ 100 Ϫ
(valuesample/valuecontrol) ϫ 100. The %Inh values (y axis) were plotted
against the drug concentration (x axis), and the according trend line of the
dose-response curve was defined and the resulting linear equation was
applied to calculate the MICs using Microsoft Office Excel 2007. The MIC
was the lowest drug concentration resulting in Ն50% growth inhibition
compared to that of the drug-free control according to the EUCAST
guidelines for flucytosine, azole antifungal agents, and echinocandins
(20). Additionally, we defined MIC0.3 as the lowest drug concentration
resulting in Ն30% growth inhibition compared to that of the drug-free
control.
The data were analyzed and evaluated from 3 biological replicates in
triplicate (n ϭ 3 [3]) (Tables 2 and 3), as well as from 4 biological repli-
cates in triplicate (n ϭ 4 [3]) (Table 4). The strains C. dubliniensis CD36/
CBS7987 and C. glabrata ATCC 90030 shown in Table 4 were analyzed in
2 biological replicates in triplicate (n ϭ 2 [3]). The R2
values for all dose-
response curves ranged between 0.87 and 0.92. Additionally, the coeffi-
cients of variation (the ratio of the standard deviation to the mean) ex-
pressed as a percentage (also referred to as relative standard deviation) for
all biological replicates ranged from 13 to 28%.
RESULTS
Outline of the study. Our main goal was to identify antifungal
activities in drugs that were designed for other purposes. Two
collection libraries, Enzo and FIMM oncology, comprising a total
of 844 agents, were screened for activity against C. albicans. A
TABLE 1 All drugs with antifungal activity identified in this study
(n ϭ 26)
Identified drug
Previously
described as
antifungal
Previously
described as
anti-Candida
Therapeutic
use
Reference
no. or
source
Haloperidol HCl Yes No Antipsychotic 21, this
study
Trifluperidol 2HCl No No Antipsychotic This study
Stanozolol No No Anemia,
angioedema
This study
Melengestrol acetate No No Anticancer This study
Megestrol acetate No No Anticancer This study
Tosedostat No No Anticancer This study
Amonafide No No Anticancer This study
Methiothepin
maleate
Yes Yes Antipsychotic 25
Rapamycin Yes Yes Anticancer 26
Auranofin Yes Yes Antirheumatic 27
Bleomycin sulfate Yes Yes Anticancer 40
Disulfiram Yes Yes Anticancer 41
Artemisinin Yes Yes Antimalarial 42
Tamoxifen citrate Yes Yes Anticancer 43
Tioconazole Yes Yes Antifungal NAa
Oxiconazole nitrate Yes Yes Antifungal NA
Ketoconazole Yes Yes Antifungal NA
Climbazole Yes Yes Antifungal NA
Miconazole Yes Yes Antifungal NA
Myclobutanil Yes Yes Antifungal NA
Fluconazole Yes Yes Antifungal NA
Amorolfine Yes Yes Antifungal NA
Bifonazole Yes Yes Antifungal NA
Sertaconazole Yes Yes Antifungal NA
Itraconazole Yes Yes Antifungal NA
Terbinafine HCl Yes Yes Antifungal NA
a
NA, not applicable.
Stylianou et al.
1056 aac.asm.org Antimicrobial Agents and Chemotherapy
onAugust21,2015byUMEAUNIVERSITYLIBRARYhttp://aac.asm.org/Downloadedfrom](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-88-320.jpg)
![major challenge for screenings with C. albicans is the characteristic
of the fungus to grow as hyphal filaments (8). Filamentation com-
plicates assessments of growth using OD, for instance, since the
number of individual cells does not increase and hyphae tend to
clump excessively. Therefore, we used a luciferase-based quanti-
fication of ATP to assess fungal viability. We additionally con-
firmed the screening results by quantifying fungal growth using
OD measurements. Both methods resulted in highly comparable
results for all tested drugs.
Seven off-target drugs revealed to have anti-Candida activi-
ties. The screen identified a total of 26 agents that are active against
C. albicans (Table 1). Of those, 12 were SADs and 7 were off-target
drugs with known antifungal activities. Additionally, the screen
revealed 7 drugs from 4 different categories of therapy with pre-
viously unidentified potent anti-Candida activities (Table 2). Two
are antipsychotic (haloperidol and trifluperidol), one is used for
the treatment of anemia (stanozolol), and 4 are used for cancer
therapy (melengestrol acetate, megestrol acetate, tosedostat, and
amonafide). Haloperidol, but not trifluperidol, has previously
been identified in a chemical-genetic screen to have antimicrobial
activity against Saccharomyces cerevisiae (21). Four agents are
FDA-approved drugs and 2 are anticancer agents (amonafide and
tosedostat) that are currently being tested in clinical trials (22, 23).
Although it has been applied in animal husbandry, of the identi-
fied drugs, only melengestrol acetate is not currently used in hu-
mans (24). Moreover, we identified the antipsychotic drug me-
thiothepin maleate, which only very recently has been identified in
a repurposing screen for anticryptococcal agents (25). We used
the immunosuppressant drug rapamycin and the antirheumatic
drug auranofin as references for the antifungal activities of the
newly identified agents (Table 2). Interestingly, rapamycin was
originally identified as an antifungal agent (26), and gold (I) com-
plexes, such as auranofin, have been recognized for their antimi-
crobial activities (27).
We determined the MIC and MIC0.3 values for C. albicans by
OD and ATP measurements. As mentioned above, the methods
resulted in highly similar values, and thus one value for each agent
is presented (Table 2). In general, the MICs were slightly lower
after 6 h than after 24 h of incubation. However, the activities of
the 7 compounds against C. albicans were stable over a period of
24 h (Table 2). Importantly, in this screen, we did not use concen-
TABLE 2 MIC and MIC0.3 values against Candida albicans type straina
Antifungal agent
This study
Other studiesc
Concn range
(g/ml)
ATP level and OD450
b
MIC at: MIC0.3 at:
6 h 24 h 6 h 24 h Cmax (g/ml) Ref. for Cmax
Haloperidol HCl 6.4 ϫ 10Ϫ5
to 3.76 0.38 3.76 0.04 0.35 2.00–3.00 44
Trifluperidol 2HCl 7 ϫ 10Ϫ5
to 4.00 4.00 4.00 0.40 0.40 UAd
UA
Stanozolol 3.3 ϫ 10Ϫ5
to 3.29 3.29 Ͼ3.29 0.30 0.30 0.007 45
Melengestrol acetate 6.8 ϫ 10Ϫ5
to 3.97 2.20 3.97 0.40 0.22 0.01 46
Megestrol acetate 6 ϫ 10Ϫ5
to 3.85 2.10 3.85 0.39 0.40 0.50–0.70 47
Tosedostat 4 ϫ 10Ϫ3
to 4.00 Ͼ4.00 Ͼ4.00 4.00 4.00 0.15 23
Amonafide 2.8 ϫ 10Ϫ3
to 2.83 Ͼ2.83 Ͼ2.83 1.50 Ͼ2.83 4.00 22
Methiothepin maleatee
7 ϫ 10Ϫ5
to 3.57 0.35 3.57 0.044 0.25 UA UA
Auranofine
1 ϫ 10Ϫ4
to 6.78 0.70 0.38 0.007 0.07 6.60 48
Rapamycine
1.55 ϫ 10Ϫ5
to 9.14 0.001 0.005 1 ϫ 10Ϫ5
1 ϫ 10Ϫ5
0.01–0.21 49
a
The data were determined from three biological replicates in triplicate (n ϭ 3 [3]). MIC, minimal concentration of drug resulting in Ն50% growth inhibition; MIC0.3, minimal
concentration of drug resulting in Ն30% growth inhibition.
b
OD450, optical density at 450 nm.
c
Cmax, plasma peak concentrations reachable in humans upon first dose of the drugs; Ref., literature reference.
d
UA, unavailable.
e
The anti-Candida albicans activities of these drugs were demonstrated previously.
TABLE 3 Comparison of SADs with off-target antifungal agents
identified in this study at a concentration of 1 Ma
Drugs MIC MIC0.3
Standard antifungalb
Tioconazole 0.39 g/ml
Oxiconazole nitrate 0.40 g/ml
Ketoconazole 0.50 g/ml
Climbazole 0.29 g/ml
Miconazole 0.40 g/ml
Fluconazole 0.30 g/ml
Amorolfine 0.32 g/ml
Myclobutanil 0.29 g/ml
Bifonazole 0.30 g/ml
Sertaconazole 0.40 g/ml
Itraconazole 0.70 g/ml
Terbinafine HCl Ͼ1 M
Nystatin Ͼ1 M
Off-target antifungal
Haloperidol HCl 0.38 g/ml
Methiothepin maleate 0.36 g/ml
Auranofin 0.68 g/ml
Trifluperidol 2HCl 0.40 g/ml
Stanozolol 0.30 g/ml
Melengestrol acetate 0.40 g/ml
Megestrol acetate 0.39 g/ml
Tosedostat Ͼ1 M
Amonafide Ͼ1 M
a
SADs, standard antifungal drugs. C. albicans SC5314 was challenged with SADs and
antifungal agents identified in this study.
b
MIC, minimal concentration of drug resulting in Ն50% growth inhibition; MIC0.3,
minimal concentration of drug resulting in Ն30% growth inhibition. The MIC and
MIC0.3 were determined by ATP measurement after 6 h of incubation. Nystatin did not
show any activity against C. albicans SC5314 in this assay. The data are determined
from three biological replicates in performed triplicate (n ϭ 3 [3]).
Dual Use of Known Drugs as Antimycotics
February 2014 Volume 58 Number 2 aac.asm.org 1057
onAugust21,2015byUMEAUNIVERSITYLIBRARYhttp://aac.asm.org/Downloadedfrom](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-89-320.jpg)
![trations of Ͼ10 M (corresponding to 3 to 10 g/ml, depending
on the molecular weight of the agent), since in the therapy of
systemic mycoses, maximal peak blood serum concentrations
above this level are unlikely to be reached. Haloperidol, trifluperi-
dol, stanozolol, melengestrol acetate, and megestrol acetate
showed MIC values of Ͻ4 g/ml. For tosedostat and amonafide,
the MIC0.3 values were determined to be 4 and 2.8 g/ml, respec-
tively. All 7 substances displayed a dose-dependent effect on C.
albicans SC5314. The antifungal activities of amonafide and tose-
dostat (Table 2) increased slowly over a wide concentration range,
from approximately 3 ϫ 10Ϫ3
g to 4 g/ml.
Novel antifungal off-target drugs and SADs have similar
anti-Candida activities. We next compared the antifungal activ-
ities of the 7 identified agents to 13 established SADs present in the
Enzo library. Notably, the novel candidates were inhibitory
against C. albicans at a level similar to those of 12 of the SADs at a
concentration of 1 M, ranging from 0.3 g to 0.7 g/ml, depend-
ing on individual molecular weights (Table 3). Terbinafine HCl,
tosedostat, and amonafide had an MIC0.3 at a concentration of Ͼ1
M. At this concentration, nystatin was the only SAD that lacked
anti-Candida activity after 6 h. Additionally, five off-target drugs
with previously known antifungal activities were also identified in
this screen, confirming that the applied methods were suitable to
identify antifungal activity against C. albicans (Table 5).
Microscopic analysis of morphological changes in C. albicans
occurring upon treatment with newly identified agents. The an-
tifungal effects of tosedostat and amonafide were milder than
those of other drugs (Tables 2 and 3). To verify the possible effects
of the selected agents identified in this study on C. albicans, we
additionally performed a direct microscopic investigation of
treated C. albicans (Fig. 1). DMSO- and BzCl-treated C. albicans
served as 100% and 0% growth controls, respectively (Fig. 1A and
B). Haloperidol and trifluperidol (Fig. 1E and G) show a very
similar effect as fluconazole (Fig. 1C). The hyphae are consider-
ably shorter, with the tendency to form branches more frequently
than with untreated control hyphae. Notably, tosedostat and
amonafide (Fig. 1F and H) caused similar morphological changes
as those observed in the samples treated with rapamycin (Fig. 1D).
The hyphae are significantly shorter, with the germ tubes having a
curved shape. The control hyphae, in contrast, are longer and
straight. Thus, our screen identified substances with comparable
effects on C. albicans morphology as the well-known antifungal
agent fluconazole or the immunosuppressant drug with antifun-
gal activity, rapamycin. This indicates that the identified agents
indeed inhibit the growth of C. albicans.
Confirmation of antifungal activities of identified drugs on
clinical isolates from different Candida spp. To assess whether
the 7 new antifungal candidate agents were also effective against
other clinical isolates of C. albicans, as well as other Candida spe-
cies, we compared C. albicans SC5314 to other clinical isolates
from C. albicans, C. dubliniensis, and C. glabrata (Table 4). C.
albicans SC5314 and the off-target drugs with known antifungal
TABLE 4 MIC and MIC0.3 values of antifungal agents for type strains and clinical isolates of Candida spp.a
Antifungal agent
Concn range
(g/ml)
C. albicans C. dubliniensis C. glabrata
SC5314 (type
strain)b
UBC3-7922 (clinical
strain)
CD36/CBS7987
(type strain)c
UBC3-3892 (clinical
strain)
ATCC 90030
(type strain)c
UBC3-7268
(clinical strain)
MIC MIC0.3 MIC MIC0.3 MIC MIC0.3 MIC MIC0.3 MIC MIC0.3 MIC MIC0.3
Haloperidol HCl 6.4 ϫ 10Ϫ3
to 3.76 3.76 0.46 3.76 0.38 3.76 0.38 Ͼ 3.76 0.38 Ͼ3.76 3.76 Ͼ3.76 Ͼ3.76
Trifluperidol 2HCl 7 ϫ 10Ϫ3
to 4.00 4.00 0.40 Ͼ4.00 0.40 Ͼ4.00 Ͼ4.00 Ͼ 4.00 0.40 Ͼ4.00 Ͼ4.00 Ͼ4.00 Ͼ4.00
Stanozolol 3.3 ϫ 10Ϫ3
to 3.29 Ͼ3.29 0.33 Ͼ3.29 0.33 Ͼ3.29 3.29 Ͼ3.29 3.29 Ͼ3.29 Ͼ3.29 Ͼ3.29 3.29
Melengestrol acetate 6.8 ϫ 10Ϫ3
to 3.97 3.97 0.37 3.97 0.40 Ͼ3.97 3.97 Ͼ3.97 1.80 Ͼ3.97 3.97 Ͼ3.97 3.97
Megestrol acetate 6 ϫ 10Ϫ3
to 3.85 3.85 0.39 3.85 0.39 Ͼ3.85 3.85 Ͼ3.85 3.85 Ͼ3.85 3.85 Ͼ3.85 3.85
Tosedostat 4 ϫ 10Ϫ3
to 4.00 Ͼ4.00 4.00 Ͼ4.00 4.00 Ͼ4.00 4.00 Ͼ4.00 4.00 Ͼ4.00 4.00 Ͼ4.00 2.00
Amonafide 2.8 ϫ 10Ϫ3
to 2.83 Ͼ2.83 1.40 Ͼ2.83 2.83 Ͼ2.83 2.83 Ͼ2.83 1.40 Ͼ2.83 Ͼ2.83 Ͼ2.83 Ͼ2.83
Methiothepin maleate 7 ϫ 10Ϫ3
to 3.57 3.30 0.31 3.30 0.36 Ͼ3.57 0.36 Ͼ3.57 0.36 3.57 0.36 3.57 0.36
Auranofin 4 ϫ 10Ϫ3
to 6.78 0.68 0.08 0.61 0.07 0.68 0.04 0.62 0.04 1.10 0.62 Ͼ3.73 3.73
Rapamycin 9 ϫ 10Ϫ3
to 9.14 0.002 Ͻ9 ϫ 10Ϫ3
0.002 Ͻ9 ϫ 10Ϫ3
0.009 Ͻ9 ϫ 10Ϫ3
0.01 Ͻ9 ϫ 10Ϫ3
0.50 0.04 0.09 0.009
a
Candida clinical strains were tested with the 7 identified drugs.
b
MIC, minimal concentration of drug resulting in Ն50% growth inhibition; MIC0.3, minimal concentration of drug resulting in Ն30% growth inhibition. MIC and MIC0.3 were
determined by ATP measurement after 24 h of incubation. The data were analyzed and evaluated from 4 biological replicates in triplicate (n ϭ 4 [3]).
c
The type strains C. dubliniensis CD36/CBS7987 and C. glabrata ATCC 90030 were analyzed in 2 biological replicates in triplicate (n ϭ 2 [3]).
TABLE 5 Nonantifungal drugs with known antifungal activitya
Antifungal agent
This study Previous studies
Reference(s)
Tested concn
(g/ml)
Incubation
times (h)
Tested concn
(g/ml)
Incubation
times (h)
Rapamycin 1.55 ϫ 10Ϫ5
to 9.14 6 and 24 0.09–100 48 and 72 26, 38
Auranofin 1 ϫ 10Ϫ4
to 6.78 6 and 24 12.5–200 48 27
Methiothepin maleate 7 ϫ 10Ϫ5
to 3.57 6 and 24 64 48 25
Bleomycin sulfate 2.6 ϫ 10Ϫ4
to 15 6 and 24 1.56 6 and 12 40
Disulfiram 5.1 ϫ 10Ϫ5
to 2.97 6 and 24 1–8 24 41
Artemisinin 4.8 ϫ 10Ϫ5
to 2.82 6 and 24 8–50 24 42
Tamoxifen citrate 9.7 ϫ 10Ϫ5
to 5.63 6 and 24 8–32 24 43
a
The tested concentrations of off-target drugs with previously demonstrated antifungal activity used in this study were compared to concentrations used in previous studies with
similar incubation times.
Stylianou et al.
1058 aac.asm.org Antimicrobial Agents and Chemotherapy
onAugust21,2015byUMEAUNIVERSITYLIBRARYhttp://aac.asm.org/Downloadedfrom](https://image.slidesharecdn.com/142218c8-9673-4277-871f-08af32c6ba20-151005120906-lva1-app6891/85/Marios_Stylianou_PhD-thesis-90-320.jpg)
































































