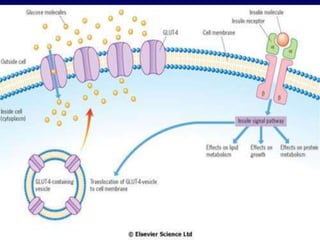
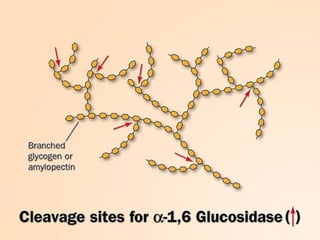
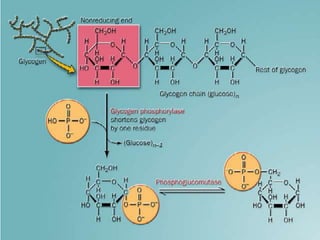
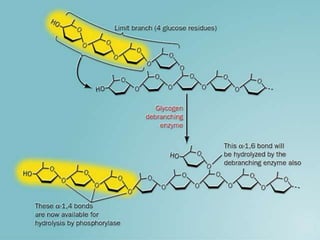
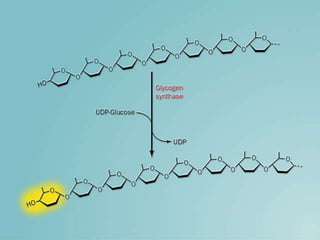
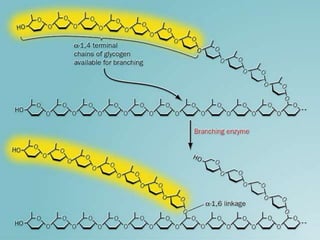
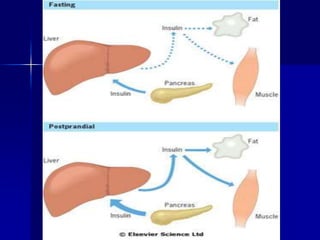
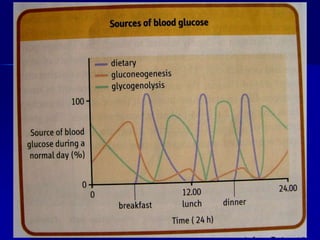
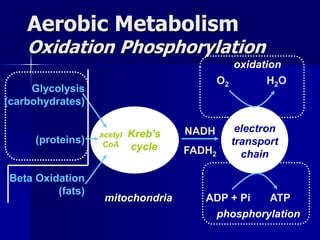
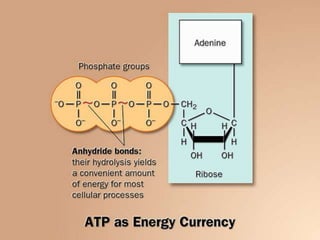
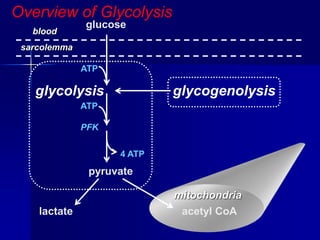
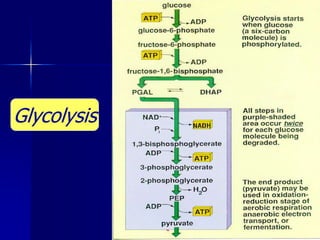
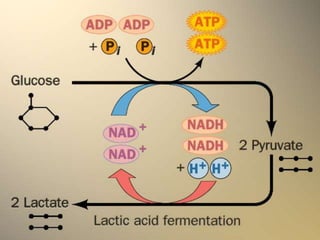
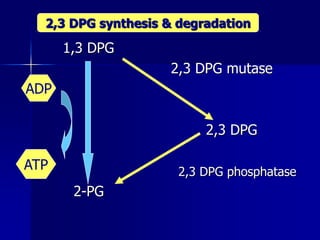
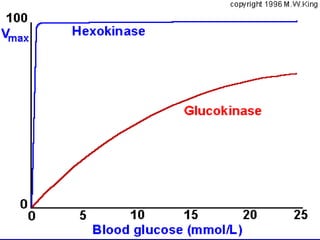
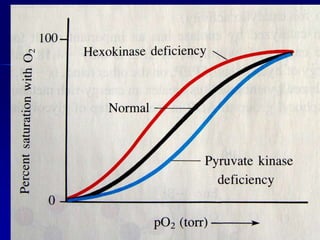
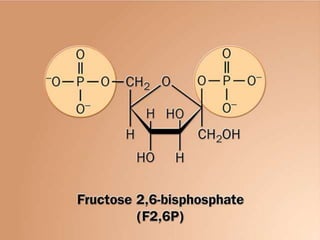
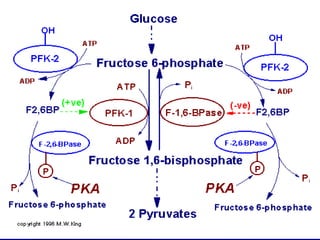
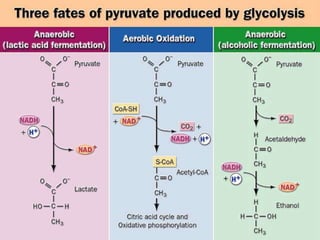
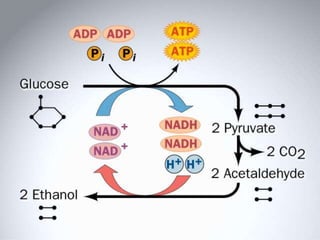
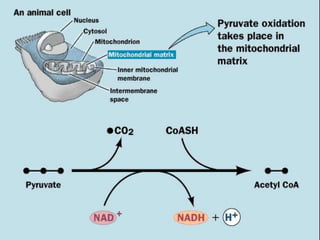
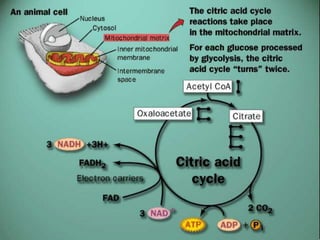
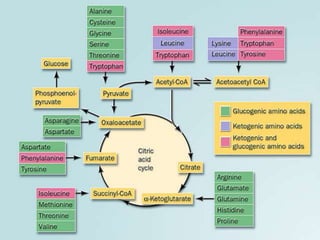
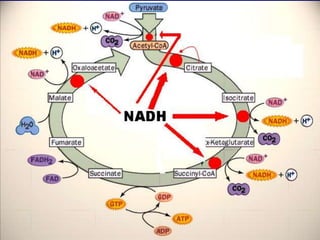
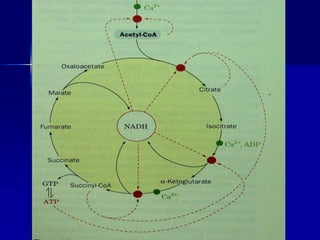
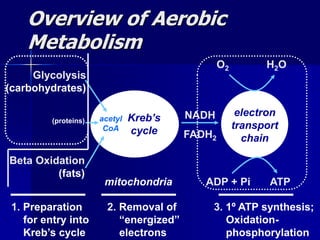
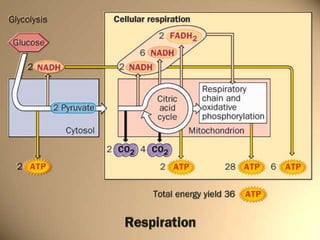
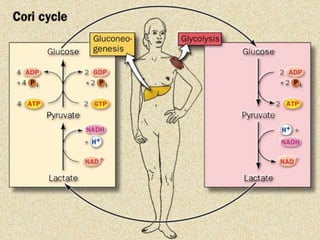
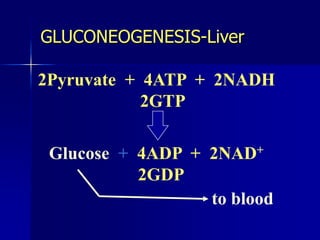
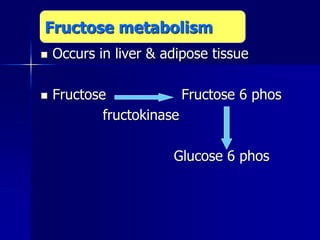
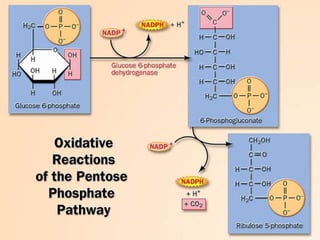
Carbohydrates are the largest source of calories and are broken down into glucose, which is the main carbohydrate used in metabolism. Glucose is absorbed into the bloodstream and transported to tissues like muscle where it is either used for energy or stored as glycogen. Carbohydrate digestion begins in the mouth and small intestine where enzymes break starches and sugars into glucose and other monosaccharides that can be absorbed. Glucose is then regulated through various pathways including glycolysis, the citric acid cycle, and oxidative phosphorylation to produce energy in the form of ATP.