All good things come in threes - species delimitation through shape analysis of saccular, lagenar and utricular otoliths.pdf
•
0 likes•3 views

The authors noticed errors in previously published otolith area values from a study on species delimitation through shape analysis of saccular, lagenar, and utricular otoliths in four Poecilia species. The errors were a factor of 1000, but do not affect the analyses, results, or conclusions of the study. Corrected tables of otolith area values for the four Poecilia species are provided.
![All good things come in threes ] species delimitation
through shape analysis of saccular, lagenar
and utricular otoliths
Tanja Schulz-MirbachA,C
and Martin PlathB
A
University of Vienna, Department of Behavioural Biology, Althanstrasse 14,
A-1090 Vienna, Austria.
B
J.W. Goethe-University, Frankfurt am Main, Evolutionary Ecology Group,
Max-von-Laue Strasse 13, 60438 Frankfurt am Main, Germany.
C
Corresponding author. Email: tanja.schulz-mirbach@univie.ac.at
Abstract. Otoliths are calcium carbonate biomineralisates in the inner ear of teleost fishes. Otoliths of the saccule
(sagittae) are known to show species-specific (or even population-specific) contour differences and, thus, are regularly
used in fisheries management for stock identification. However, the other two otolith types from the utricle (lapilli) and
lagena (asterisci) are typically neglected in studies of this kind, such that little information is available regarding potential
species-specific contour differences. Using four species of livebearing fishes of the genus Poecilia (Cyprinodontiformes,
Poeciliidae), we compared contour outlines of all three otolith types by applying Fourier shape analysis and tested for
species delimitation success of the different otolith types alone, and all three otoliths combined. Our results indicated that
also lapilli and especially asterisci convey species-specific information, and the classification success of discriminant
function analyses was highest when combining shape information from all three otolith types. We propose that future
studies on species delimitation or stock identification may benefit from considering all three otolith types together.
Additional keywords: inner ear, Fast Fourier Tranform, freshwater fishes, Guppy, Molly.
Received 15 May 2012, accepted 17 August 2012, published online 24 October 2012
Introduction
Otoliths are massive calcium carbonate biomineralisates over-
lying the sensory epithelia in the inner ear of teleost fishes.
Within the endolymph-filled, sac-like compartments of the
inner ear (the so-called otolithic end organs), otoliths are inti-
mately attached to the sensory epithelia, and movement of the
fish’s body relative to the slower motion of the otolith, which is
about three times denser than the sensory epithelium, provokes a
stimulation of the sensory hair cells. In this way, the teleost inner
ear can perceive linear acceleration and sound (Popper 2011).
According to the respective end organ, the following three dif-
ferent otolith types can be distinguished: the otolith of the sac-
cule (sagitta), of the lagena (asteriscus) and of the utricle
(lapillus).
Although saccular otoliths are widely used in fisheries
management for stock identification through shape analysis
(Lombarte et al. 2006: AFORO database; Cañás et al. 2012;
Tuset et al. 2012), in paleontology for the reconstruction of past
fish diversity and paleoecology (e.g. Reichenbacher and
Kowalke 2009) and in ecology for the investigation of migration
patterns through isotope and trace-element analyses (Stransky
et al. 2005; Rohtla et al. 2012), only few studies have investi-
gated inter- and intra-specific shape variation of the other two
otolith types (Assis 2003, 2005; Schulz-Mirbach et al. 2011a).
In the majority of teleosts, the saccular otolith is the largest of the
three otolith types, and it has been claimed that only this otolith
type provides sufficient species-specific morphological infor-
mation to allow species delimitation (e.g. Popper and Coombs
1982; Nolf 1985), at least in ‘non-otophysan’ teleosts. In
otophysans (Cypriniformes, Characiformes, Siluriformes and
Gymnotiformes), however, including all taxa possessing a
Weberian apparatus (Fink and Fink 1996), the asteriscus and/
or lapillus are by far larger than the fragile, needle-like sagitta.
Few studies have illustrated and described the diversity of
lapilli and asterisci in otophysans (e.g. Frost 1925; Adams
1940; Berinkey 1956; Assis 2003, 2005; Schulz-Mirbach and
Reichenbacher 2006), and even fewer descriptions of non-
otophysan lapilli and asterisci exist (e.g. Frost 1926; Assis
2003, 2005); all of these studies described otolith contours on
a purely qualitative basis.
In his overview of asterisci and lapilli in teleost fishes, Assis
(2003, 2005) argued that those otoliths types, too, are likely to
show species-specific morphological differences. Moreover,
two recent studies quantifying contour outlines of all three
otolith types in different populations of the non-otophysan
speciesPoeciliamexicanaSteindachner, 1863,andP. sulphuraria
(Álvarez, 1948) have provided first evidence that contours of
lapilli and asterisci may indeed convey species-specific (or even
CSIRO PUBLISHING
Marine and Freshwater Research, 2012, 63, 934–940
http://dx.doi.org/10.1071/MF12132
Journal compilation Ó CSIRO 2012 www.publish.csiro.au/journals/mfr
Short communication](data:image/gif;base64,R0lGODlhAQABAIAAAAAAAP///yH5BAEAAAAALAAAAAABAAEAAAIBRAA7)
Recommended
More Related Content
Similar to All good things come in threes - species delimitation through shape analysis of saccular, lagenar and utricular otoliths.pdf
Similar to All good things come in threes - species delimitation through shape analysis of saccular, lagenar and utricular otoliths.pdf (20)
More from Liz Adams
More from Liz Adams (20)
Recently uploaded
Recently uploaded (20)
All good things come in threes - species delimitation through shape analysis of saccular, lagenar and utricular otoliths.pdf
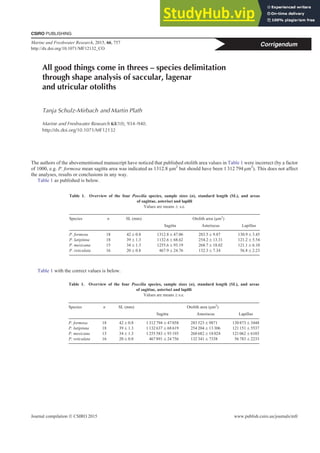
- 1. All good things come in threes – species delimitation through shape analysis of saccular, lagenar and utricular otoliths Tanja Schulz-Mirbach and Martin Plath Marine and Freshwater Research 63(10), 934–940. http://dx.doi.org/10.1071/MF12132 The authors of the abovementioned manuscript have noticed that published otolith area values in Table 1 were incorrect (by a factor of 1000, e.g. P. formosa mean sagitta area was indicated as 1312.8 mm2 but should have been 1 312 794 mm2 ). This does not affect the analyses, results or conclusions in any way. Table 1 as published is below. Table 1 with the correct values is below. Table 1. Overview of the four Poecilia species, sample sizes (n), standard length (SL), and areas of sagittae, asterisci and lapilli Values are means s.e. Species n SL (mm) Otolith area (mm2 ) Sagitta Asteriscus Lapillus P. formosa 18 42 0.8 1312.8 47.06 283.5 9.87 130.9 3.45 P. latipinna 18 39 1.3 1132.6 68.62 254.2 13.31 121.2 5.54 P. mexicana 15 34 1.3 1255.6 93.19 268.7 18.02 121.1 6.10 P. reticulata 16 20 0.8 467.9 24.76 132.3 7.34 56.8 2.23 Table 1. Overview of the four Poecilia species, sample sizes (n), standard length (SL), and areas of sagittae, asterisci and lapilli Values are means s.e. Species n SL (mm) Otolith area (mm2 ) Sagitta Asteriscus Lapillus P. formosa 18 42 0.8 1 312 794 47 058 283 523 9871 130 873 3448 P. latipinna 18 39 1.3 1 132 637 68 619 254 204 13 306 121 151 5537 P. mexicana 15 34 1.3 1 255 583 93 193 268 682 18 024 121 062 6103 P. reticulata 16 20 0.8 467 891 24 756 132 341 7338 56 783 2233 CSIRO PUBLISHING Marine and Freshwater Research, 2015, 66, 757 http://dx.doi.org/10.1071/MF12132_CO Journal compilation Ó CSIRO 2015 www.publish.csiro.au/journals/mfr Corrigendum
- 2. All good things come in threes ] species delimitation through shape analysis of saccular, lagenar and utricular otoliths Tanja Schulz-MirbachA,C and Martin PlathB A University of Vienna, Department of Behavioural Biology, Althanstrasse 14, A-1090 Vienna, Austria. B J.W. Goethe-University, Frankfurt am Main, Evolutionary Ecology Group, Max-von-Laue Strasse 13, 60438 Frankfurt am Main, Germany. C Corresponding author. Email: tanja.schulz-mirbach@univie.ac.at Abstract. Otoliths are calcium carbonate biomineralisates in the inner ear of teleost fishes. Otoliths of the saccule (sagittae) are known to show species-specific (or even population-specific) contour differences and, thus, are regularly used in fisheries management for stock identification. However, the other two otolith types from the utricle (lapilli) and lagena (asterisci) are typically neglected in studies of this kind, such that little information is available regarding potential species-specific contour differences. Using four species of livebearing fishes of the genus Poecilia (Cyprinodontiformes, Poeciliidae), we compared contour outlines of all three otolith types by applying Fourier shape analysis and tested for species delimitation success of the different otolith types alone, and all three otoliths combined. Our results indicated that also lapilli and especially asterisci convey species-specific information, and the classification success of discriminant function analyses was highest when combining shape information from all three otolith types. We propose that future studies on species delimitation or stock identification may benefit from considering all three otolith types together. Additional keywords: inner ear, Fast Fourier Tranform, freshwater fishes, Guppy, Molly. Received 15 May 2012, accepted 17 August 2012, published online 24 October 2012 Introduction Otoliths are massive calcium carbonate biomineralisates over- lying the sensory epithelia in the inner ear of teleost fishes. Within the endolymph-filled, sac-like compartments of the inner ear (the so-called otolithic end organs), otoliths are inti- mately attached to the sensory epithelia, and movement of the fish’s body relative to the slower motion of the otolith, which is about three times denser than the sensory epithelium, provokes a stimulation of the sensory hair cells. In this way, the teleost inner ear can perceive linear acceleration and sound (Popper 2011). According to the respective end organ, the following three dif- ferent otolith types can be distinguished: the otolith of the sac- cule (sagitta), of the lagena (asteriscus) and of the utricle (lapillus). Although saccular otoliths are widely used in fisheries management for stock identification through shape analysis (Lombarte et al. 2006: AFORO database; Cañás et al. 2012; Tuset et al. 2012), in paleontology for the reconstruction of past fish diversity and paleoecology (e.g. Reichenbacher and Kowalke 2009) and in ecology for the investigation of migration patterns through isotope and trace-element analyses (Stransky et al. 2005; Rohtla et al. 2012), only few studies have investi- gated inter- and intra-specific shape variation of the other two otolith types (Assis 2003, 2005; Schulz-Mirbach et al. 2011a). In the majority of teleosts, the saccular otolith is the largest of the three otolith types, and it has been claimed that only this otolith type provides sufficient species-specific morphological infor- mation to allow species delimitation (e.g. Popper and Coombs 1982; Nolf 1985), at least in ‘non-otophysan’ teleosts. In otophysans (Cypriniformes, Characiformes, Siluriformes and Gymnotiformes), however, including all taxa possessing a Weberian apparatus (Fink and Fink 1996), the asteriscus and/ or lapillus are by far larger than the fragile, needle-like sagitta. Few studies have illustrated and described the diversity of lapilli and asterisci in otophysans (e.g. Frost 1925; Adams 1940; Berinkey 1956; Assis 2003, 2005; Schulz-Mirbach and Reichenbacher 2006), and even fewer descriptions of non- otophysan lapilli and asterisci exist (e.g. Frost 1926; Assis 2003, 2005); all of these studies described otolith contours on a purely qualitative basis. In his overview of asterisci and lapilli in teleost fishes, Assis (2003, 2005) argued that those otoliths types, too, are likely to show species-specific morphological differences. Moreover, two recent studies quantifying contour outlines of all three otolith types in different populations of the non-otophysan speciesPoeciliamexicanaSteindachner, 1863,andP. sulphuraria (Álvarez, 1948) have provided first evidence that contours of lapilli and asterisci may indeed convey species-specific (or even CSIRO PUBLISHING Marine and Freshwater Research, 2012, 63, 934–940 http://dx.doi.org/10.1071/MF12132 Journal compilation Ó CSIRO 2012 www.publish.csiro.au/journals/mfr Short communication
- 3. population-specific) information (Schulz-Mirbach et al. 2010, 2011a). However, no study has directly quantified and com- pared the potential of asterisci and lapilli for species delimita- tion. In the present study, we address this question on the basis of shape (contour) differences of all three otolith types in a group of closely related, non-otophysan teleost fishes from the family Poeciliidae (livebearers). Using four representatives of the genus Poecilia, we demonstrate that species could be success- fully discriminated with each of the otolith types alone and that information from all three types combined yields the best separation. Material and methods Study animals and otolith dissection In total, we analysed 67 otolith ‘triplets’, i.e. sagittae, asterisci and lapilli of four species of the poeciliid genus Poecilia, namely Guppies, Poecilia reticulata Peters, 1859, Atlantic mollies, P. mexicana, Amazon mollies, P. formosa (Girard, 1859) and Sailfin mollies, P. latipinna (Le Sueur, 1821) (Table 1). Specimens of P. reticulata investigated in our study were provided by I. Schlupp and stemmed from a stock at the Department of Zoology of the University of Oklahoma in Norman that was founded using wild-caught fish from a feral population inhabiting the San Antonio River in Texas. Speci- mens of P. formosa and P. latipinna were wild catches origi- nating from Martindale (San Marcos River, central Texas), whereas P. mexicana individuals were collected near Tapijulapa (Tabasco, southern Mexico). P. latipinna was introduced to the San Marcos River from Louisiana in the 1930s, and P. formosa from Olmito near Brownsville in the 1950s (Schlupp et al. 2002; Lamatsch et al. 2004). P. formosa is a gynogenetic species that probably originated from a single, inter-specific hybridisation event between a P. latipinna-like male and a P. mexicana limantouri female ,100 000–40 000 years ago (Schartl et al. 1995). It is an all-female species that relies on the coexistence with at least one so-called sperm-donor species (mainly P. latipinna or P. mexicana), the sperm of which is needed to trigger embryogenesis, whereas genetic material of the sperm donor is excluded from the unreduced diploid oocyte. As a result, inheritance of this hybrid species is clonal. We preserved adult specimens in 70% ethanol until dissec- tion. Then, we measured standard length to the nearest milli- metre and determined sex by inspection of the gonopodium (the transformed anal fin) of males and, if necessary, by inspection of the ovary of females. To avoid any potentially confounding effects caused by potential sex differences, we used only females (Schulz-Mirbach et al. 2010). We extracted the three otolith types – sagitta, asteriscus and lapillus – through ventral dissection from both labyrinths, cleaned them of organic resi- dues with 1% potassium hydroxide solution for 4–6 h and rinsed them several times in distilled water. Afterwards, we stored otoliths dry in small plastic cells (Krantz cells, Krantz, Bonn, Germany). Shape analysis We positioned either left or right otoliths from each specimen with their lateral (sagittae, asterisci) or dorsal face (lapilli) down on plasticine lying plainly under a stereo-microscope (Leica MZ 6; camera: Leica DFC 295, Leica Microsystems, Wetzlar, Germany), with the medial (sagittae, asterisci) or ventral face (lapilli) oriented horizontally. We then took digital images at maximum magnifications of 40–102 (sagittae), or 80–128 (asterisci and lapilli) by using the Leica Image Access Software (IMAGIC 1000, Imagic Bildverarbeitung AG, Glattbrugg, Switzerland) while focusing on the outermost otolith contour. We processed all digital images in Adobe Photoshop CS2 (Adobe Systems Incorporated, San Jose, CA, USA) by gener- ating a contrast of 100% along the otolith contour. Prior to shape analysis, images of right otoliths were mirrored. Terminology of sagittae follows Nolf (1985) and Tuset et al. (2008), the termi- nology of asterisci and lapilli follows Assis (2003, 2005) (see also Fig. 1). As starting points for digitising outlines, we used the tip of the rostrum of sagittae and asterisci, and the extremum posterior of lapilli. Raw (x- and y-) coordinates of each otolith contour were obtained by using tpsDIG2 (Rohlf 2004) and subjected to the shape analysis software Hshape (Crampton and Haines 1996; Haines and Crampton 2000) consisting of the three programs Hangle, Hmatch and Hcurve. The output of Hangle and Hmatch are Fourier descriptors (FDs), describing the contour by a combination of sine- and cosine-waves. In this type of Fourier analysis, Fourier functions are fitted to a function of the tangent angle dependent on the arc length (see Haines and Crampton (2000) for a comparison of their Fourier method with the ‘classical’ elliptic Fourier analysis), and calculated with the Fast Fourier Transform (FFT) algorithm. Normalisation of size was performed automatically in Hangle (smoothing iterations: 16 for sagittae, 7 for astersici and lapilli each), whereas normal- isation of rotation and starting point was ensured by applying Hmatch. We plotted the amplitude against the harmonic number so as to evaluate the minimum number of harmonics and, thus, Fourier descriptors that were necessary for an adequate shape description. Thirty-four FDs were used to analyse contours of sagittae, 30 for asterisci and 32 for lapilli, respectively. To visualise differences in otolith contours among species, we back-calculated averaged FDs of each species and otolith type into x- and y-coordinates (1024 per contour) by applying Hcurve, resulting in mean shapes of the groups. Outlines of sagittae from P. formosa (Nre-analysed ¼ 18) and P. latipinna (Nre-analysed ¼ 18), and partly sagittae, asterisci and lapilli from P. mexicana (Nre-analysed ¼ 15) published in Schulz-Mirbach et al. (2008a, 2008b, 2011a) were re-analysed in the present study. Table 1. Overview of the four Poecilia species, sample sizes (n), standard length (SL), and areas of sagittae, asterisci and lapilli Values are means s.e. Species n SL (mm) Otolith area (mm2 ) Sagitta Asteriscus Lapillus P. formosa 18 42 0.8 1312.8 47.06 283.5 9.87 130.9 3.45 P. latipinna 18 39 1.3 1132.6 68.62 254.2 13.31 121.2 5.54 P. mexicana 15 34 1.3 1255.6 93.19 268.7 18.02 121.1 6.10 P. reticulata 16 20 0.8 467.9 24.76 132.3 7.34 56.8 2.23 Species delimitation by multiple otolith types Marine and Freshwater Research 935
- 4. Statistical analyses We performed all statistical analyses in SPSS 15.0 (SPSS Inc. 2006) and PAST 2.15 (Hammer et al. 2001). In a first step, we conducted three variance–covariance-based principal compo- nent analyses (one PCA for each otolith type), using the FDs as input variables to reduce the number of variables for further multivariate analyses (see below). We evaluated relevant prin- cipal components (PCs) explaining more variance than expected by chance alone according to the ‘broken stick model’ sensu Jackson (1993). FDs per se cannot be assigned to special parts of the contour; however, by interpreting the morphospace of the PCAs by using synthetic model shapes (for a detailed descrip- tion see Haines and Crampton (2000) and Schulz-Mirbach et al. (2008b)), we could identify the otolith characters explained by the respective PC (see also Fig. 1). Synthetic model shapes were calculated for each principal component corresponding to unit standard deviation steps at 2, 0 and þ2. In a second step, we used all retained PCs as input variables for a MANOVA (the first eight PCs for sagittae, the first three for asterisci and the first four for lapilli). We asked to which extent species were significantly separated from one another on the basis of a given otolith type (or specific otolith characters) and, thus, conducted all pair-wise post hoc tests (Tamhane2) for each PC separately. So as to illustrate the magnitude of species separation, we performed four canonical discriminant function analyses (CDAs) using PCs as input variables. We calculated three CDAs using only one otolith type each, in which the first eight (sagittae), three (asterisci), or four (lapilli) PCs served as input variables; the fourth CDA was based on all otolith types combined, using the aforementioned 15 PCs together. PAST v. 2.15 (Hammer et al. 2001) was used to estimate 95% confidence ellipses for species. We additionally tested for classification success using jack-knifed cross-validation and calculated the overall discriminatory power of the respective otolith character. To evaluate the relative contribution of sagittae, asterisci and lapilli to the overall classification success in the combined dataset, we calculated the mean discriminant coefficient (MDC) according to the equation provided in Backhaus et al. (2006, p. 188), as follows: Mean bi ¼ S bi lk=ltotal ð Þ ð Þ; where bi is the discriminant coefficient of variable i (here: PC), lk is the eigenvalue of the respective discriminant function k, SAGITTA Posterodorsal edge (tip-like) Posteroventral edge PC1 PC2 PC3 PC4 PC1 PC1 PC2 PC3 PC4 PC3 PC2 PC5 PC6 PC7 PC8 Ventral rim Antirostrum Excisura ostii Rostrum Extremum ventralis Rostrum asterisci Excisura major edge Anterodorsal Medial edge Extremum anterior Incision Extremum posterior Postero- dorsal edge ASTERISCUS LAPILLUS (a1) (b1) (c1) (a2) (b2) (c2) ⫺2 0 ⫹2 ⫺2 0 ⫹2 ⫺2 0 ⫹2 Fig. 1. Illustration of morphological characters of each otolith type explained by the respective principal components (PCs) using synthetic model shapes. In a1–c1 otolith terminology is shown by means of scanning electron microscope images of representative otoliths of Poecilia mexicana. Black arrows in a2–c2 indicate parts of the otolith contour explained by the respective PC (for description of those characters see also Table 2). Scale bars ¼ 100 mm. 936 Marine and Freshwater Research T. Schulz-Mirbach and M. Plath
- 5. and ltotal is the sum of the eigenvalues of all discriminant functions (here: discriminant function 1 through 3). To account for potential effects of otolith size on shape differences (i.e. allometric effects), we determined otolith size as the orthogonal projection of the area of the macula-oriented face of each otolith type, using tpsDIG2 (Rohlf 2004) as a proxy for size (e.g. Lombarte et al. 2010). We then tested principal components for potential size correlations by calculating Pearson’s correlation coefficient (rP) between the respective PC and ln-transformed otolith area. Alpha levels (0.05) were Bonferroni-corrected for multiple comparisons (Rice 1989). Because PC 1 for sagittae and PC 3 for lapilli showed significant size correlations (PC 1: rP ¼ 0.75, P , 0.0001; PC 3: rP ¼ 0.80, P , 0.0001), we conducted another CDA on the combined dataset, with those two PCs removed, i.e. using 13 input variables instead of 15. Results Three PCs of each otolith type contributed significantly to species discrimination (Table 2). In the case of sagittae (Fig. 1a2), these characters were identified as the overall shape varying from elongate to short (PC 1), the position of the dorsal tip and the rostrum tip (PC 2), and the shape of the postero- ventral edge (angular to pointed; PC 4). In asterisci, the devel- opment of the excisura major (PC 1), the development of the postero-dorsal edge and the rostrum asterisci (PC 2), and antero- dorsal edge (PC 3) significantly contributed to species separa- tion (Fig. 1b2). In lapilli, the development of the medial edge (PC 1), as well as the incision (PC 2), and shape of the extremum posterior, varying from angular to round (PC 3; Fig. 1c2), were identified as separating characters. The combined analysis including PCs from all three otolith types (sagittae: PC1–8; asterisci: PC1–3; lapilli: PC1–4) resulted in a statistically significant separation among the four Poecilia species (MANOVA, Wilk’s l ¼ 0.0008, P , 0.0001). The canonical discriminant function analyses found species separation to be best when shape information of all three otolith types was combined (Fig. 2a–c v. d, Table 3), resulting in a classification success of 98.5%. The overall classification suc- cess was still high (97.0%) after the removal of the size- correlated PCs (Table 3). The CDAs of the datasets using only one otolith type found asterisci (91.0%) and lapilli (88.1%) to show higher overall classification success than sagittae (83.6%; Table 3). On the basis of the shapes of asterisci, P. reticulata was clearly separated from the other three Poecilia species, whereas the shapes of lapilli distinguished P. formosa from the remaining species because of the prominent development of the extremum posterior in P. formosa (Fig. 1c1). For the most part, the same characters as identified to significantly separate the species in the three separate PCAs also had the highest discriminatory power in the canonical discriminant analysis using the combined dataset (see values of the mean discriminatory coefficient, MDC in Table 2). Exceptions were characters of the sagittae; PC 3 showed a low MDC value, whereas PC 7 (ratio of antirostrum to rostrum) and PC 8 (development of the ventral rim) were not significant in the MANOVA post hoc tests, but had high MDC values and also showed differences among species in the overlay of the mean shapes of the sagittae (Fig. 2). Table 2. Significance of separation of the four Poecilia species provided and characters explained by the first eight (sagittae), three (asterisci) or four (lapilli) principal components (PCs) based on the Fourier descriptors (see Fig. 1) Significant P-values and values for the mean discriminatory coefficient (MDC) of .0.3 are in bold. Var%, explained variance in %. Size-correlated PCs are indicated by asterisks Principal component Sagittae Asterisci Lapilli Var% Character explained Species separation (Tamhane2 post hoc) MDC Var% Character explained Species separation (Tamhane2 post hoc) MDC Var% Character explained Species separation (Tamhane2 post hoc) MDC PC 1 17.2* Overall shape: elongate or short P # 0.031 0.477 27.6 Excisura major; postero- dorsal edge P , 0.001 0.630 29.5 Medial edge P , 0.001 0.470 PC 2 11.2 Position of dorsal tip; rostrum P , 0.001 0.593 11.8 Postero-dorsal edge; rostrum P # 0.039 0.389 11.6 Development of incision P # 0.003 0.216 PC 3 7.9 Excisura ostii; postero-ventral edge P . 0.05 0.101 8.8 Antero-dorsal edge P # 0.005 0.184 9.4* Extremum posterior: angular or round P , 0.001 0.212 PC 4 7.1 Postero-ventral edge P # 0.037 0.037 8.7 Extremum posterior: flat or pointed P . 0.05 0.206 PC 5 6.9 Postero-ventral edge; rostrum P . 0.05 0.122 PC 6 6.3 Postero-ventral edge P . 0.05 0.159 PC 7 6.0 Postero-dorsal edge; ratio of antirostrum to rostrum P . 0.05 0.522 PC 8 4.9 Ventral rim flat or convex P . 0.05 0.306 Total 67.5 48.2 59.2 Species delimitation by multiple otolith types Marine and Freshwater Research 937
- 6. Discussion Our study, for the first time, demonstrated, on a quantitative basis, that shape information not only of sagittae, but also of lapilli and especially asterisci significantly contributes to spe- cies discrimination. Moreover, we found that a combined analysis of shape information from all three otolith types results in a much better separation than the analysis of one otolith type alone. For the most part, the same characters of sagitta, asteriscus and lapillus shape that separated species in our present study were also found to separate locally adapted populations of P. mexicana living under starkly divergent habitat conditions, especially in habitats containing toxic hydrogen sulfide of volcanic origin (Schulz-Mirbach et al. 2011a: fig. 2 therein). Our results thus support the notion of Assis (2003, 2005) and Schulz-Mirbach et al. (2011a) that asterisci and lapilli in non- otophysans have the potential to show species-specific (or even population-specific) differences in gross morphology, namely, outlines. We encourage future studies to evaluate whether asterisci and lapilli of species that are relevant from an economic perspective are also a source of additional information for stock identification and, thus, fisheries management. This may be of particular interest when stock or species discrimination based on shape information of the sagittae alone is weak. It needs to be mentioned, however, that the asterisci and lapilli of non- otophysan teleosts tend to be much smaller than is the sagittae and that members of the Cyprinodontiformes (including the family Poeciliidae) represent an exception in that they possess comparatively large asterisci and lapilli (Schulz-Mirbach et al. 2011b). Then again, the small size of asterisci and lapilli in other teleost groups does not necessarily preclude morphological complexity and species-specificity of outlines. Several studies noted that sagittae may convey morphologi- cal information suitable for phylogenetic reconstructions (Nolf and Tyler 2006; Assis 2003, 2005), and it was therefore suggested to include saccular otolith characters in phylogenetic analyses of teleosts (see Knudsen et al. 2007; Lombarte et al. 2010). We propose that future studies in the field of teleost phylogeny should also take the morphology of asterisci and lapilli into account. An exciting set of questions arises from our present study. For example, do changes in one otolith type correlate with changes in the other two types (concerted evolution), or are the different end organs and their corresponding otoliths subject to entirely different selective forces and undergo independent (mosaic) evolution? Our study has provided first evidence regarding this question. For instance, although P. formosa is rather similar to the other Poecilia species investigated herein, with respect to the shape of its sagitta and asteriscus, it displays a very distinct lapillus shape, separating it from P. reticulata as well as from both parental species P. latipinna and P. mexicana. Among vertebrates, the structure of the utricle appears to be rather conservative compared with the two other otolithic end organs –theexceptionbeingtheorderClupeiformes(e.g.Popper and Platt 1979) and some species of deep-sea fishes (Deng 2009) – and the utricular otolith (lapillus) has been hypothesised to have the least potential for species delimitation of the three otolith types (Assis 2005). Still, we found lapilli to separate even closely related species; basically, they did not performed worse than did sagittae. Although all three end organs may serve both the sense of balance and the sense of hearing, the utricle in most 0.16 0.12 0.08 0.04 ⫺0.04 ⫺0.08 ⫺0.12 ⫺0.12 ⫺0.08 ⫺0.04 Discriminant axis 1 (64.2%) Discriminant axis 2 (31.3%) Sagittae Asterisci Lapilli All Mean shapes 0.04 0.08 0 ⫺0.12 ⫺0.08 ⫺0.04 Discriminant axis 1 (84.8%) 0.04 0.08 0 ⫺0.12 ⫺0.08 ⫺0.04 Discriminant axis 1 (66.0%) 0.04 0.08 0 ⫺0.12 ⫺0.08 ⫺0.04 Discriminant axis 1 (44.4%) 0.04 0.08 0 ⫺0.16 0 0.18 0.15 0.12 0.09 0.03 0.06 0 ⫺0.03 Discriminant axis 2 (9.8%) ⫺0.06 ⫺0.09 0.16 0.12 0.08 0.04 0 ⫺0.04 ⫺0.08 ⫺0.12 Discriminant axis 2 (23.6%) ⫺0.16 0.16 0.12 0.08 0.04 ⫺0.04 0 ⫺0.08 ⫺0.12 Discriminant axis 2 (39.6%) ⫺0.16 (a) (b) (c) (d) P. latipinna P. mexicana P. formosa P. reticulata Fig. 2. Plots of canonical discriminant analyses (discriminant axis 2 against axis 1) of the contours of (a–c) each otolith type alone, or (d) all three otolith types combined. Separation among species was best when shape information of sagittae, asterisci and lapilli was combined (d). Ellipses represent 95% confidence ellipses for each group (i.e. species). Mean shapes of each otolith type illustrate parts of the contours differing among species. 938 Marine and Freshwater Research T. Schulz-Mirbach and M. Plath
- 7. teleosts is claimed to play a major role in the vestibular sense, and the saccule (and potentially the lagena) are mainly involved in the perception of sound (Popper and Schilt 2008; Popper 2011). Therefore, the different end organs and their otoliths may indeed not all be projected to the same selective forces. How- ever, even if the utricle is the most conservative of the three end organs, it is likely that different species exhibit differences in the development of their vestibular sense, e.g. because they differ in their need of sophisticated maneuverability. This, in turn, may translate into slight differences in utricule and lapillus morphol- ogy among species. Acknowledgements We are grateful to I. Schlupp (Norman, Oklahoma), R. Riesch (Raleigh, North Carolina), and M. Tobler (Stillwater, Oklahoma) for providing wild- caught specimens and I. Schlupp for providing P. reticulata from his labo- ratory. The Mexican Government (Permiso de Pesca de Fomento No. DGOPA.06192.240608.-1562), Semarnat (No. SGPA/DGVS/04148/08 and SGPA/DGVS/04751/08), as well as the Municipal of Tacotalpa (SM/1133/208) provided collection permits. Finally, we thank C. A. Assis and an anonymous reviewer for their constructive remarks. References Adams, L. A. (1940). Some characteristic otoliths of American Ostario- physi. Journal of Morphology 66, 497–527. doi:10.1002/JMOR. 1050660307 Assis, C. A. (2003). The lagenar otoliths of teleosts: their morphology and application in species identification, phylogeny and systematics. Journal of Fish Biology 62, 1268–1295. doi:10.1046/J.1095-8649.2003.00106.X Assis, C. A. (2005). The utricular otoliths, lapilli, of teleosts: their morphol- ogy and relevance for species identification and systematics studies. Scientia Marina 69, 259–273. Backhaus, K., Erichson, B., Plinke, W., and Weiber, R. (2006). ‘Multivariate Analysemethoden. Eine anwendungsorientierte Einführung.’ 11th edn. (Springer-Verlag: Berlin.) Berinkey, L. (1956). The taxonomical examination of the otoliths of the Cyprinidae of Hungary. Annales Historico2Naturales Musei Nationalis Hungarici 7, 455–462. Cañás, L., Stransky, C., Schlickeisen, J., Sampedro, M. P., and Fariña, A. C. (2012). Use of the otolith shape analysis in stock identification of anglerfish (Lophius piscatorius) in the Northeast Atlantic. ICES Journal of Marine Science 69, 250–256. doi:10.1093/ICESJMS/FSS006 Crampton, J. S., and Haines, A. J. (1996). User’s manual for programs HANGLE, HMATCH, and HCURVE for the Fourier shape analysis of Table 3. Jack-knifed classification matrix of the canonical discriminant analyses among the four Poecilia species using principal components (PCs) as input variables either from each otolith type separately or from all three otolith types combined The combined dataset was also analysed with the two size-correlated PCs (PC 1 of the sagittae and PC 3 of lapilli, see also Table 2) removed. The percentages in rows represent the classification into species given in columns; the corresponding number of specimens is given in parentheses. Percentages of correctly classified individuals and significant P-values are in bold Otolith type P. formosa P. latipinna P. mexicana P. reticulata Overall class success (%) Wilk’s l x2 P-value Sagitta 83.6 0.030 210.0 0.001 P. formosa 83.3 (15) 0 (0) 16.7 (3) 0 (0) P. latipinna 0 (0) 88.9 (16) 5.6 (1) 5.6 (1) P. mexicana 26.7 (4) 6.7 (1) 60.0 (9) 6.7 (1) P. reticulata 0 (0) 0 (0) 0 (0) 100 (16) Asteriscus 91.0 0.037 205.3 0.001 P. formosa 100 (18) 0 (0) 0 (0) 0 (0) P. latipinna 11.1 (2) 83.3 (15) 5.6 (1) 0 (0) P. mexicana 6.7 (1) 13.3 (2) 80.0 (12) 0 (0) P. reticulata 0 (0) 0 (0) 0 (0) 100 (16) Lapillus 88.1 0.048 188.9 0.001 P. formosa 94.4 (17) 0 (0) 0 (0) 5.6 (1) P. latipinna 0 (0) 88.9 (16) 5.6 (1) 5.6 (1) P. mexicana 0 (0) 6.7 (1) 80.0 (12) 13.3 (2) P. reticulata 0 (0) 6.3 (1) 6.3 (1) 87.5 (14) All 98.5 0.001 405.8 0.001 P. formosa 100 (18) 0 (0) 0 (0) 0 (0) P. latipinna 0 (0) 100 (18) 0 (0) 0 (0) P. mexicana 0 (0) 6.7 (1) 93.3 (14) 0 (0) P. reticulata 0 (0) 0 (0) 0 (0) 100 (16) All (size effects removed) 97.0 0.003 344.2 0.001 P. formosa 100 (18) 0 (0) 0 (0) 0 (0) P. latipinna 0 (0) 94.4 (17) 5.6 (1) 0 (0) P. mexicana 0 (0) 6.7 (1) 93.3 (14) 0 (0) P. reticulata 0 (0) 0 (0) 0 (0) 100 (16) Species delimitation by multiple otolith types Marine and Freshwater Research 939
- 8. two-dimensional outlines. Institute of Geological Nuclear Sciences, Science Report 96, 1–28. Deng, X. (2009). Comparative studies on the structure of the ears of deep-sea fishes. Ph.D. Thesis, University of Maryland, College Park, MD. Fink, S. V., and Fink, W. L. (1996). Interrelationships of Ostariophysan fishes (Teleostei). In ‘Interrelationships of Fishes’. (Eds M. L. Stiassny, L. R. Parenti and G. D. Johnson.) pp. 209–249. (Academic Press: San Diego, CA.) Frost, G. A. (1925). A comparative study of the otoliths of the neopterygian fishes. Annals Magazine of Natural History 15, 152–163. doi:10.1080/ 00222932508633194 Frost, G. A. (1926). A comparative study of the otoliths of the neopterygian fishes. Annals Magazine of Natural History 17, 99–104. doi:10.1080/ 00222932608633376 Haines, A. J., and Crampton, J. S. (2000). Improvements to the method of Fourier shape analysis as applied in morphometric studies. Palaeontol- ogy 43, 765–783. doi:10.1111/1475-4983.00148 Hammer, Ø., Harper, D. A. T., and Ryan, P. D. (2001). Past: paleontological statistics software package for education and data analysis. Palaeonto- logia Electronica 4, 1–9. Jackson, D. A. (1993). Stopping rules in principal components analysis: a comparison of heuristical and statistical approaches. Ecology 74, 2204–2214. doi:10.2307/1939574 Knudsen, S. W., Møller, P. R., and Gravlund, P. (2007). Phylogeny of the snailfishes (Teleostei: Liparidae) based on molecular and morphological data. Molecular Phylogenetics and Evolution 44, 649–666. doi:10.1016/ J.YMPEV.2007.04.005 Lamatsch, D. K., Nanda, I., Schlupp, I., Epplen, J. T., Schmid, M., and Schartl, M. (2004). Distribution and stability of supernumerary micro- chromosomes in natural populations of the Amazon molly, Poecilia formosa. Cytogenetic and Genome Research 106, 189–194. doi:10.1159/000079286 Lombarte, A., Chic, Ò., Parisi-Baradad, V., Olivella, R., Piera, J., and Garcı́a-Ladona, E. (2006). A web-based environment from shape analy- sis of fish otoliths. The AFORO database. Scientia Marina 70, 147–152. Lombarte, A., Palmer, M., Matallanas, J., Gómez-Zurita, J., and Morales- Nin, B. (2010). Ecomorphological trends and phylogenetic inertia of otolith sagittae in Nototheniidae. Environmental Biology of Fishes 89, 607–618. doi:10.1007/S10641-010-9673-2 Nolf, D. (1985). Otolithi piscium. In ‘Handbook of Paleoichthyology 10’. (Ed. H.-P. Schultze.) (Gustav Fischer Verlag: Stuttgart.) Nolf, D., and Tyler, J. C. (2006). Otolith evidence concerning interrelation- ships of caproid, zeiform and tetraodontiform fishes. Bulletin de l’Insti- tut Royal des Sciences Naturelles de Belgique. Biologie 76, 147–189. Popper, A. N. (2011). Auditory system morphology. In ‘Encyclopedia of Fish Physiology: from Genome to Environment’. (Ed. A. P. Farrell.) Vol. 1. pp. 252–261. (Academic Press: San Diego, CA.) Popper, A. N., and Coombs, S. (1982). The morphology and evolution of the ear in Actinopterygian fishes. American Zoologist 22, 311–328. Popper, A. N., and Platt, C. (1979). The herring ear has a unique receptor pattern. Nature 280, 832–833. doi:10.1038/280832A0 Popper, A. N., and Schilt, C. R. (2008). Hearing and acoustic behavior: basic and applied considerations. In ‘Fish Bioacoustics’. (Eds J. F. Webb, R. R. Fay and A. N. Popper.) pp. 17–48. (Springer: New York.) Reichenbacher, B., and Kowalke, T. (2009). Neogene and present-day zoogeography of killifishes (Aphanius and Aphanolebias) in the Mediterranean and Paratethys areas. Palaeogeography, Palaeoclimatol- ogy, Palaeoecology 281, 43–56. doi:10.1016/J.PALAEO.2009.07.008 Rice, W. R. (1989). Analyzing tables of statistical tests. Evolution 43, 223–225. doi:10.2307/2409177 Rohlf, F. J. (2004). ‘tpsDIG2’. Available at http://life.bio.sunysb.edu/ morph/ [accessed 8 June 2011]. Rohtla, M., Vetemaa, M., Urtson, K., and Soesoo, A. (2012). Early life migration patterns of Baltic Sea pike Esox lucius. Journal of Fish Biology 80, 886–893. doi:10.1111/J.1095-8649.2012.03226.X Schartl, M., Wilde, B., Schlupp, I., and Parzefall, J. (1995). Evolutionary origin of a parthenoform, the Amazon molly Poecilia formosa, on the basis of a molecular genealogy. Evolution 49, 827–835. doi:10.2307/ 2410406 Schlupp, I., Parzefall, J., and Schartl, M. (2002). Biogeography of the Amazon molly, Poecilia formosa. Journal of Biogeography 29, 1–6. doi:10.1046/J.1365-2699.2002.00651.X Schulz-Mirbach, T., and Reichenbacher, B. (2006). Reconstruction of Oligocene and Neogene freshwater fish faunas – an actualistic study on cypriniform otoliths. Acta Palaeontologica Polonica 51, 283–304. Schulz-Mirbach, T., Scherb, H., and Reichenbacher, B. (2008a). Are hybridization and polyploidization phenomena detectable in the fossil record? – A case study on otoliths of a natural hybrid, Poecilia formosa (Teleostei: Poeciliidae). Neues Jahrbuch für Geologie und Palaontolo- gie. Abhandlungen 249(2), 223–238. doi:10.1127/0077-7749/2008/ 0249-0223 Schulz-Mirbach, T., Stransky, C., Schlickeisen, J., and Reichenbacher, B. (2008b). Differences in otolith morphologies between surface- and cave- dwelling populations of Poecilia mexicana (Teleostei, Poeciliidae) reflect adaptations to life in an extreme habitat. Evolutionary Ecology Research 10, 537–558. Schulz-Mirbach, T., Ladich, F., Riesch, R., and Plath, M. (2010). Otolith morphology and hearing abilities in cave- and surface-dwelling ecotypes of the Atlantic molly, Poecilia mexicana (Teleostei: Poeciliidae). Hearing Research 267, 137–148. doi:10.1016/J.HEARES.2010.04.001 Schulz-Mirbach, T., Riesch, R., Garcı́a de León, F. J., and Plath, M. (2011a). Effects of extreme habitat conditions on otolith morphology – a case study on extremophile livebearing fishes (Poecilia mexicana, P. sul- phuraria). Zoology (Jena, Germany) 114, 321–334. doi:10.1016/ J.ZOOL.2011.07.004 Schulz-Mirbach, T., Heß, M., and Plath, M. (2011b). Inner ear morphology in the Atlantic Molly, Poecilia mexicana – first detailed microanatomi- cal study of the inner ear of a cyprinodontiform species. PLoS ONE 6(11), e27734. doi:10.1371/JOURNAL.PONE.0027734 SPSS Inc (2006). ‘SPSS. Ver. 15.0. Base.’ (SPSS Inc.: Chicago, IL.) Stransky, C., Garbe-Schönberg, C.-D., and Günther, D. (2005). Geographic variation and juvenile migration in Atlantic redfish inferred from otolith microchemistry. Marine and Freshwater Research 56, 677–691. doi:10.1071/MF04153 Tuset, V.M., Lombarte, A., and Assis, C.A. (2008). Otolith atlas for the western Mediterranean, north and central eastern Atlantic. Scientia Marina 72, 7–198. doi:10.3989/SCIMAR.2008.72S1199 Tuset, V. M., Azzurro, E., and Lombarte, A. (2012). Identification of Lessepsian fish species using the sagittal otolith. Scientia Marina 76, 289–299. doi:10.3989/SCIMAR.03420.18E www.publish.csiro.au/journals/mfr 940 Marine and Freshwater Research T. Schulz-Mirbach and M. Plath